

Wood Degradation by Fomitiporia mediterranea M. Fischer: Exploring Fungal Adaptation Using Metabolomic Networking

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
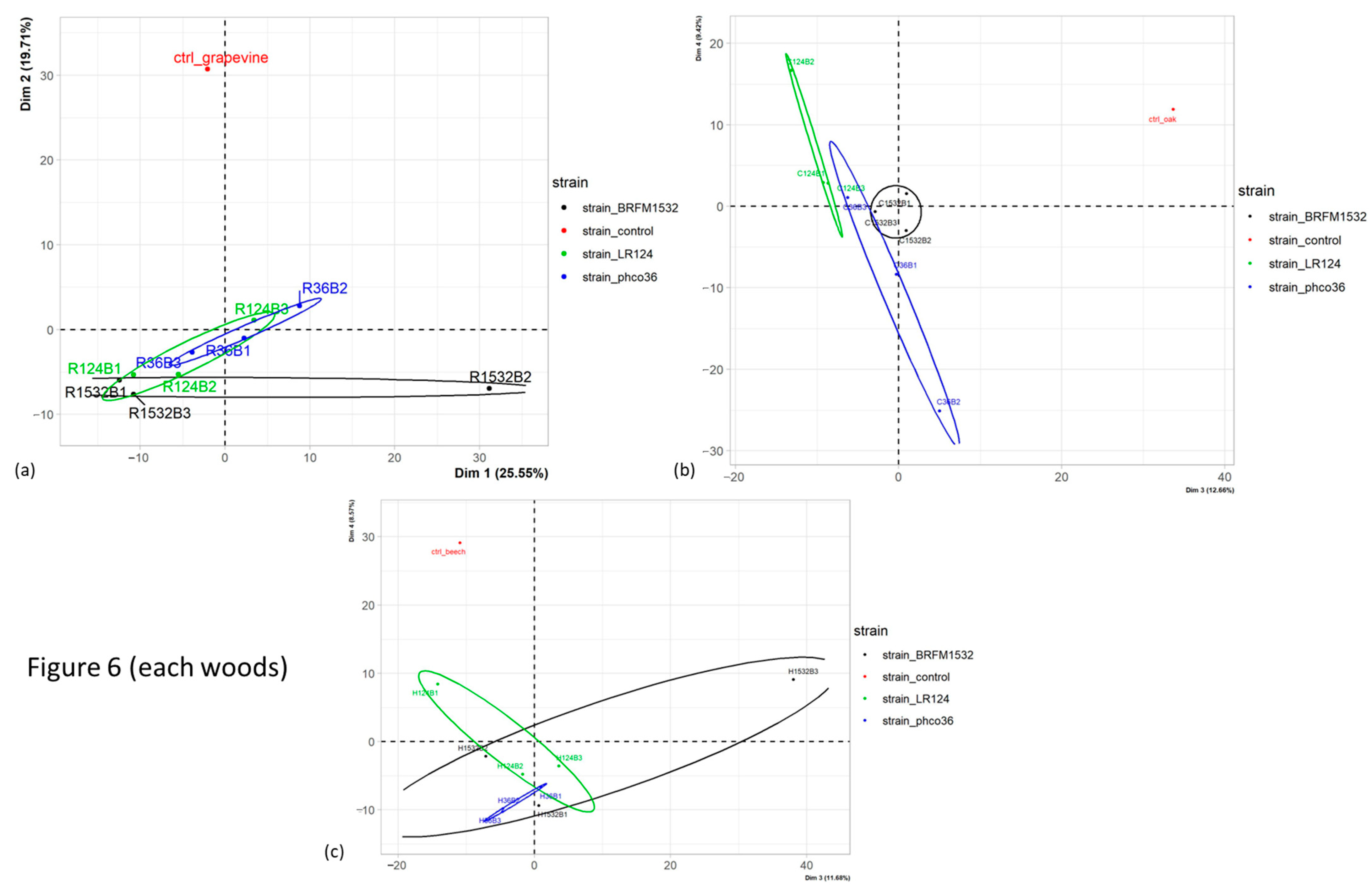{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Schilling, M.; Levasseur, M.; Barbier, M.; Oliveira-Correia, L.; Henry, C.; Touboul, D.; Farine, S.; Bertsch, C.; Gelhaye, E. Wood Degradation by Fomitiporia mediterranea M. Fischer: Exploring Fungal Adaptation Using Metabolomic Networking. J. Fungi 2023, 9, 536. https://doi.org/10.3390/jof9050536
Schilling M, Levasseur M, Barbier M, Oliveira-Correia L, Henry C, Touboul D, Farine S, Bertsch C, Gelhaye E. Wood Degradation by Fomitiporia mediterranea M. Fischer: Exploring Fungal Adaptation Using Metabolomic Networking. Journal of Fungi. 2023; 9(5):536. https://doi.org/10.3390/jof9050536
Chicago/Turabian StyleSchilling, Marion, Marceau Levasseur, Muriel Barbier, Lydie Oliveira-Correia, Céline Henry, David Touboul, Sibylle Farine, Christophe Bertsch, and Eric Gelhaye. 2023. "Wood Degradation by Fomitiporia mediterranea M. Fischer: Exploring Fungal Adaptation Using Metabolomic Networking" Journal of Fungi 9, no. 5: 536. https://doi.org/10.3390/jof9050536
APA StyleSchilling, M., Levasseur, M., Barbier, M., Oliveira-Correia, L., Henry, C., Touboul, D., Farine, S., Bertsch, C., & Gelhaye, E. (2023). Wood Degradation by Fomitiporia mediterranea M. Fischer: Exploring Fungal Adaptation Using Metabolomic Networking. Journal of Fungi, 9(5), 536. https://doi.org/10.3390/jof9050536