
Molecular Response of Meyerozyma guilliermondii to Patulin: Transcriptomic-Based Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast
2.2. Total RNA Extraction from Yeast
2.3. High Throughput RNA Sequencing and Bioinformatic Analysis
2.4. Validation of DEGs by RT-qPCR
3. Results
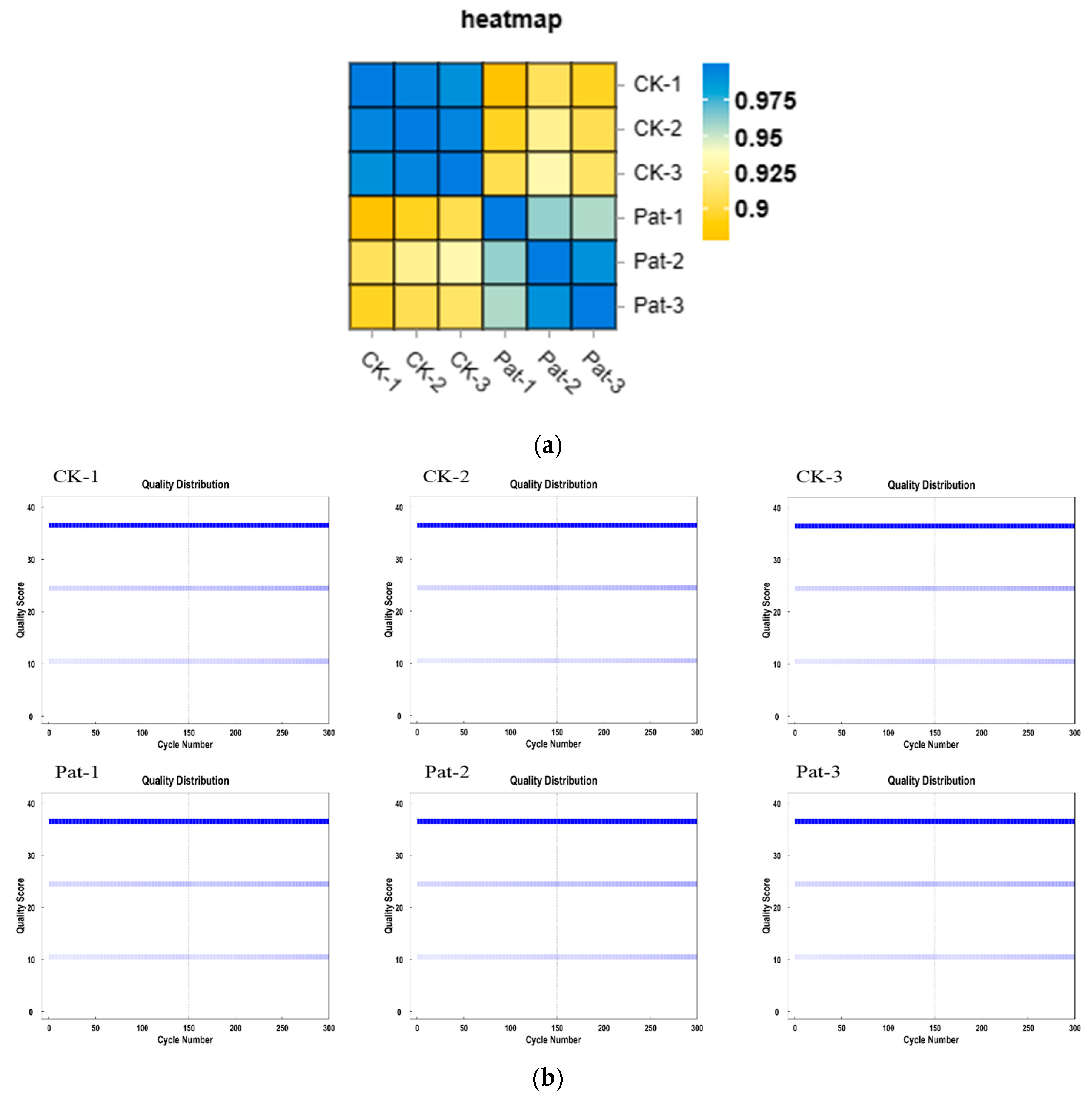
3.1. Sample Relationship Analysis
3.2. Differential Expression of Genes
3.3. GO Enrichment Analysis of DEGs
3.4. COG Enrichment Analysis of Differentially Expressed Genes
3.5. KEGG Pathway Enrichment Analysis of DEGs
3.6. Differential Expression of Genes Involved in PAT Detoxification
3.7. Validation of RNA-seq Data by RT-qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahato, D.K.; Kamle, M.; Sharma, B.; Pandhi, S.; Devi, S.; Dhawan, K.; Selvakumar, R.; Mishra, D.; Kumar, A.; Arora, S.; et al. Patulin in food: A mycotoxin concern for human health and its management strategies. Toxicon 2021, 198, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Ngea, G.L.N.; Yang, Q.Y.; Castoria, R.; Zhang, X.Y.; Routledge, M.N.; Zhang, H.Y. Recent trends in detecting, controlling, and detoxifying of patulin mycotoxin using biotechnology methods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2447–2472. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Yuan, Y.; Yue, T.; Hatab, S.; Wang, Z. Binding mechanism of patulin to heat-treated yeast cell. Lett. Appl. Microbiol. 2012, 55, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.Y.; Feussner, K.; Wu, T.; Yan, F.J.; Karlovsky, P.; Zheng, X.D. Detoxification of mycotoxin patulin by the yeast Rhodosporidium paludigenum. Food Chem. 2015, 179, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peng, H.M.; Wang, X.; Li, B.Q.; Long, M.Y.; Tian, S.P. Biodegradation mechanisms of patulin in Candida guilliermondii: An iTRAQ-based proteomic analysis. Toxins 2017, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, X.Y.; Zheng, X.F.; Apaliya, M.T.; Yang, Q.Y.; Zhao, L.N.; Gu, X.Y.; Zhang, H.Y. Control of postharvest blue mold decay in pears by Meyerozyma guilliermondii and it’s effects on the protein expression profile of pears. Postharvest Biol. Technol. 2018, 136, 124–131. [Google Scholar] [CrossRef]
- Yang, Q.Y.; Ma, J.F.; Solairaj, D.; Fu, Y.; Zhang, H.Y. Efficacy of Meyerozyma guilliermondii in controlling patulin production by Penicillium expansum in shuijing pears. Biol. Control. 2022, 168, 104856. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, Q.Y.; Solairaj, D.; Esa, A.G.; Michael, N.R.; Zhang, H.Y. Biodegradation of mycotoxin patulin by the yeast Meyerozyma guilliermondii. Biol. Control. 2021, 160, 104692. [Google Scholar] [CrossRef]
- Michaelis, S.; Barrowman, J. Biogenesis of the Saccharomyces cerevisiae pheromone a-factor, from yeast mating to human disease. Microbiol. Mol. Biol. Rev. 2012, 76, 626–651. [Google Scholar] [CrossRef]
- Shim, E.; Park, S.H. Identification of a novel Ser/Thr protein phosphatase Ppq1 as a negative regulator of mating MAP kinase pathway in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2014, 443, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Papp, G.; Horvath, E.; Mike, N.; Gazdag, Z.; Belagyi, J.; Gyongyi, Z.; Banfalvi, G.; Hornok, L.; Pesti, M. Regulation of patulin-induced oxidative stress processes in the fission yeast Schizosaccharomyces pombe. Food Chem. Toxicol. 2012, 50, 3792–3798. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-S.; Lin, Y.-S.; Chio, L.-M.; Liang, W.-Z. Evaluation of the mycotoxin patulin on cytotoxicity and oxidative stress in human glioblastoma cells and investigation of protective effect of the antioxidant N-acetylcysteine (NAC). Toxicon 2023, 221, 106957. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Lei, B.K.; Zhou, N.; Feng, B.W.; Yao, W.; Zhao, X.; Yu, Y.; Lu, H. Identification of novel genes involved in DNA damage response by screening a genome-wide Schizosaccharomyces pombe deletion library. Bmc Genomics 2012, 13, 662. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Petes, T.D. Recombination between Homologous Chromosomes Induced by Unrepaired UV-Generated DNA Damage Requires Mus81p and Is Suppressed by Mms2p. PLoS Genet. 2015, 11, e1005026. [Google Scholar] [CrossRef]
- Goel, S.; Krishnamurthy, S.; Hampsey, M. Mechanism of start site selection by RNA polymerase II Interplay between TFIIB and Ssl2/XPB helicase subunit of TFIIH. J. Biol. Chem. 2012, 287, 557–567. [Google Scholar] [CrossRef]
- Tang, H.M.V.; Pan, K.W.; Kong, K.Y.E.; Hu, L.G.; Chan, L.C.; Siu, K.L.; Sun, H.Z.; Wong, C.M.; Jin, D.Y. Loss of APD1 in Yeast Confers Hydroxyurea Sensitivity Suppressed by Yap1p Transcription Factor. Sci. Rep. 2015, 5, 7897. [Google Scholar] [CrossRef]
- Fei, X.W.; Li, X.H.; Li, P.; Deng, X.D. Involvement of Chlamydomonas DNA damage tolerence gene UBC2 in lipid accumulation. Algal Res.-Biomass Biofuels Bioprod. 2017, 22, 148–159. [Google Scholar] [CrossRef]
- Freiberg, G.; Mesecar, A.D.; Huang, H.H.; Hong, J.Y.; Liebman, S.W. Characterization of novel rad6/ubc2 ubiquitin-conjugating enzyme mutants in yeast. Curr. Genet. 2000, 37, 221–233. [Google Scholar] [CrossRef]
- Borst, P.; Evers, R.; Kool, M.; Wijnholds, J. A family of drug transporters: The multidrug resistance-associated proteins. Jnci-J. Natl. Cancer Inst. 2000, 92, 1295–1302. [Google Scholar] [CrossRef]
- Knorre, D.A.; Galkina, K.V.; Shirokovskikh, T.; Banerjee, A.; Prasad, R. Do multiple drug resistance transporters interfere with cell functioning under normal conditions? Biochem.-Mosc. 2020, 85, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Wu, T.S.; Yu, F.Y.; Su, C.C. Induction of oxidative stress response by the mycotoxin patulin in mammalian cells. Toxicol. Sci. 2007, 95, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Manoharlal, R.; Puri, N.; Prasad, R. Antifungal curcumin induces reactive oxygen species and triggers an early apoptosis but prevents hyphae development by targeting the global repressor TUP1 in Candida albicans. Biosci. Rep. 2010, 30, 391–404. [Google Scholar] [CrossRef]
- Leach, M.D.; Stead, D.A.; Argo, E.; MacCallum, D.M.; Brown, A.J.P. Molecular and proteomic analyses highlight the importance of ubiquitination for the stress resistance, metabolic adaptation, morphogenetic regulation and virulence of Candida albicans. Mol. Microbiol. 2011, 79, 1574–1593. [Google Scholar] [CrossRef]
- Xu, N.; Qian, K.F.; Dong, Y.J.; Chen, Y.L.; Yu, Q.L.; Zhang, B.; Xing, L.J.; Li, M.C. Novel role of the Candida albicans ferric reductase gene CFL1 in iron acquisition, oxidative stress tolerance, morphogenesis and virulence. Res. Microbiol. 2014, 165, 252–261. [Google Scholar] [CrossRef]
- Giancaspero, T.A.; Dipalo, E.; Miccolis, A.; Boles, E.; Caselle, M.; Barile, M. Alteration of ROS Homeostasis and Decreased Lifespan in S. cerevisiae Elicited by Deletion of the Mitochondrial Translocator FLX1. BioMed Res. Int. 2014, 2014, 101286. [Google Scholar] [CrossRef]
- Hagen, S.; Drepper, F.; Fischer, S.; Fodor, K.; Passon, D.; Platta, H.W.; Zenn, M.; Schliebs, W.; Girzalsky, W.; Wilmanns, M.; et al. Structural insights into cargo recognition by the yeast PTS1 receptor. J. Biol. Chem. 2015, 290, 26610–26626. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, F.; Li, S.Y.; Dai, J.B.; Deng, H.T. Glutaredoxin deletion shortens chronological life span in Saccharomyces cerevisiae via ROS-mediated Ras/PKA activation. J. Proteome Res. 2018, 17, 2318–2327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Liu, B.; Li, X.H.; Ouyang, Z.G.; Huang, L.; Hong, Y.B.; Zhang, H.J.; Li, D.Y.; Song, F.M. The de novo biosynthesis of vitamin B6 is required for disease resistance against Botrytis cinerea in tomato. Mol. Plant-Microbe Interact. 2014, 27, 688–699. [Google Scholar] [CrossRef]
- Venkatesh, A.; Murray, A.L.; Coughlan, A.Y.; Wolfe, K.H. Giant GAL gene clusters for the melibiose-galactose pathway in Torulaspora. Yeast 2021, 38, 117–126. [Google Scholar] [CrossRef]
- Torkko, J.M.; Koivuranta, K.T.; Kastaniotis, A.J.; Airenne, T.T.; Glumoff, T.; Ilves, M.; Hartig, A.; Gurvitz, A.; Hiltunen, J.K. Candida tropicalis expresses two mitochondrial 2-enoyl thioester reductases that are able to form both homodimers and heterodimers. J. Biol. Chem. 2003, 278, 41213–41220. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Orozco, H.; Friant, S.; Aranda, A.; del Olmo, M. The Saccharomyces cerevisiae flavodoxin-like proteins Ycp4 and Rfs1 play a role in stress response and in the regulation of genes related to metabolism. Arch. Microbiol. 2011, 193, 515–525. [Google Scholar] [CrossRef]
- Davis-Kaplan, S.R.; Compton, M.A.; Flannery, A.R.; Ward, D.M.; Kaplan, J.; Stevens, T.H.; Graham, L.A. PKR1 encodes an assembly factor for the yeast V-type ATPase. J. Biol. Chem. 2006, 281, 32025–32035. [Google Scholar] [CrossRef]
- Mathangi, S.; Chandrasekharan, H.V.A.; Ramya, L. Exploring the molecular insights of intrinsically disordered mitochondrial intermembrane protein in its unbound and substrate-bound state. Mol. Simul. 2020, 46, 1239–1247. [Google Scholar] [CrossRef]
- Kim, J.; Park, M.; Do, E.; Jung, W.H. Mitochondrial protein Nfu1 influences homeostasis of essential metals in the human fungal pathogen Cryptococcus neoformans. Mycobiology 2014, 42, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Guiguen, A.; Soutourina, J.; Dewez, M.; Tafforeau, L.; Dieu, M.; Raes, M.; Vandenhaute, J.; Werner, M.; Hermand, D. Recruitment of P-TEFb (Cdk9-Pch1) to chromatin by the cap-methyl transferase Pcm1 in fission yeast. Embo J. 2007, 26, 1552–1559. [Google Scholar] [CrossRef] [PubMed]
- Saha, D.; Khandelia, P.; O’Keefe, R.T.; Vijayraghavan, U. Saccharomyces cerevisiae nineteen complex (NTC)-associated factor Bud31/Ycr063w assembles on precatalytic spliceosomes and improves first and second step pre-mRNA splicing efficiency. J. Biol. Chem. 2012, 287, 5390–5399. [Google Scholar] [CrossRef]
- Florencio-Martinez, L.E.; Cano-Santiago, A.; Mondragon-Rosas, F.; Gomez-Garcia, M.; Flores-Perez, C.; Roman-Carraro, F.C.; Barocio-Rodriguez, L.A.; Manning-Cela, R.G.; Nepomuceno-Mejia, T.; Martinez-Calvillo, S. Participation of TFIIIB subunit Brf1 in transcription regulation in the human pathogen Leishmania major. Genes 2021, 12, 280. [Google Scholar] [CrossRef]
- Zheng, S.D.; Ye, K.Q. Purification, crystallization and preliminary X-ray diffraction analysis of Imp3 in complex with an Mpp10 peptide involved in yeast ribosome biogenesis. Acta Crystallogr. Sect. F-Struct. Biol. Commun. 2014, 70, 918–921. [Google Scholar] [CrossRef]
- Ianiri, G.; Idnurm, A.; Castoria, R. Transcriptomic responses of the basidiomycete yeast Sporobolomyces sp. to the mycotoxin patulin. BMC Genom. 2016, 17, 210. [Google Scholar] [CrossRef]
- Ramirez-Zavala, B.; Manz, H.; Englert, F.; Rogers, P.D.; Morschhauser, J. A Hyperactive form of the zinc cluster transcription factor Stb5 causes YOR1 overexpression and beauvericin resistance in Candida albicans. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, R.E.; Feng, Z.L.; Schwelm, A.; Yang, Y.Z.; Zhang, S.G. Functional analysis of a putative dothistromin toxin MFS transporter gene. Toxins 2009, 1, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Kawai, S.; Murata, K. Secretion of quinolinic acid, an intermediate in the kynurenine pathway, for utilization in NAD(+) biosynthesis in the yeast Saccharomyces cerevisiae. Eukaryotic Cell 2013, 12, 648–653. [Google Scholar] [CrossRef]
- Belenky, P.; Stebbins, R.; Bogan, K.L.; Evans, C.R.; Brenner, C. Nrt1 and Tna1-independent export of NAD(+) precursor vitamins promotes NAD(+) homeostasis and allows engineering of vitamin production. PLoS ONE 2011, 6, e01655-18. [Google Scholar] [CrossRef]
- Remy, E.; Nino-Gonzalez, M.; Godinho, C.P.; Cabrito, T.R.; Teixeira, M.C.; Sa-Correia, I.; Duque, P. Heterologous expression of the yeast Tpo1p or Pdr5p membrane transporters in Arabidopsis confers plant xenobiotic tolerance. Sci. Rep. 2017, 7, 1–14. [Google Scholar]
- Yang, Y.K.; Pan, Y.Y.; Liu, G.Q.; Dong, G.B.; Zhan, C.J.; Koffas, M.A.G.; Liu, X.X.; Li, Y.; Liu, C.L.; Bai, Z.H. Glycerol transporter 1 (Gt1) and zinc-regulated transporter 1 (Zrt1) function in different modes for zinc homeostasis in Komagataella phaffii (Pichia pastoris). Biotechnol. Lett. 2020, 42, 2413–2423. [Google Scholar] [CrossRef]
- Stolz, J.; Caspari, T.; Carr, A.M.; Sauer, N. Cell division defects of Schizosaccharomyces pombe liz1(-) mutants are caused by defects in pantothenate uptake. Eukaryotic Cell 2004, 3, 406–412. [Google Scholar] [CrossRef]
- Persson, B.; Kallberg, Y.; Bray, J.E.; Bruford, E.; Dellaporta, S.L.; Favia, A.D.; Duarte, R.G.; Jornvall, H.; Kavanagh, K.L.; Kedishvili, N.; et al. The SDR (short-chain dehydrogenase/reductase and related enzymes) nomenclature initiative. Chem.-Biol. Interact. 2009, 178, 94–98. [Google Scholar] [CrossRef]
- Zheng, X.F.; Yang, Q.Y.; Zhao, L.N.; Apaliya, M.T.; Zhang, X.Y.; Zhang, H.Y. Crosstalk between proteins expression and lysine acetylation in response to patulin stress in Rhodotorula mucilaginosa. Sci. Rep. 2017, 7, 9843. [Google Scholar] [CrossRef]
- Xing, M.Y.; Chen, Y.; Li, B.Q.; Tian, S.P. Characterization of a short-chain dehydrogenase/reductase and its function in patulin biodegradation in apple juice. Food Chem. 2021, 348, 129046. [Google Scholar] [CrossRef]
- Zhang, Y.; Solairaj, D.; Ngea, G.L.N.; Yang, Q.Y.; Zhang, H.Y. Overexpression of the SDR gene improves the ability of Meyerozyma guilliermondii to degrade patulin in pears and juices. Food Chem. 2023, 417, 135785. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Description |
|---|---|---|
| PGUG_00994 | NOP53 | Ribosome biogenesis protein |
| PGUG_01741 | MRR1 | Hypothetical protein |
| PGUG_01881 | SPCC24B 10.20 | Hypothetical protein |
| PGUG_05287 | YML 131W | Hypothetical protein |
| PGUG_01077 | SPCC 663.08c | Hypothetical protein |
| PGUG_04552 | GRP 2 | NADPH-dependent methylglyoxal reductase |
| PGUG_05192 | YPR022C | Hypothetical protein |
| PGUG_04009 | YJR096W | Hypothetical protein |
| PGUG_05193 | SPCC663.09c | Hypothetical protein |
| PGUG_01005 | GSH 2 | Hypothetical protein |
| PGUG_03271 | MDR 1 | Multidrug resistance protein 1 |
| PGUG_00888 | YHK8 | Probable drug/proton antiporter |
| Gene ID | Gene Name | Log2 FC | Description |
|---|---|---|---|
| Protein modification | |||
| PGUG_02927 | STE14 | 1.05 | Protein-S-isoprenylcysteine O-methyltransferase |
| PGUG_00221 | PPQ1 | 1.32 | Serine/threonine-protein phosphatase |
| DNA damage repair | |||
| PGUG_04669 | RAD14 | 2.26 | DNA repair protein |
| PGUG_03571 PGUG_00484 | SDS22 | 1.91 2.30 | Protein phosphatase 1 regulatory subunit |
| PGUG_04889 | SPBC2A9.02 | 1.34 | Hypothetical protein |
| PGUG_01549 | MAG1 | 2.65 | DNA-3-methyladenine glycosylase |
| PGUG_04806 | RPN4 | 1.14 | Transcriptional regulator |
| PGUG_05450 | SSL2 | 1.12 | DNA repair factor IIH helicase subunit XPB |
| Resistance and drug resistance | |||
| PGUG_04879 | YCF1 | 1.67 | Metal resistance protein |
| PGUG_03453 | APD1 | 1.32 | Actin patches distal protein 1 |
| PGUG_05641 | UBC2 | 1.23 | Ubiquitin-conjugating enzyme E2 |
| PGUG_01353 PGUG_01354 PGUG_00697 | SNG1 | 1.80 1.80 3.06 | Nitrosoguanidine resistance protein |
| PGUG_03271 PGUG_05048 PGUG_04886 | MDR1 | 10.11 9.43 3.22 | Multidrug resistance protein 1 |
| PGUG_03388 | FCR3 | 1.67 | Fluconazole resistance protein 3 |
| PGUG_03871 | FAP1 | 2.08 | FKBP12-associated protein 1 |
| PGUG_04884 PGUG_01265 | AZR1 | 2.15 2.88 | Azole resistance protein 1 |
| Antioxidant stress | |||
| PGUG_03261 PGUG_05692 PGUG_04552 PGUG_05714 | GRP2 | 6.70 1.03 4.79 3.40 | NADPH-dependent methylglyoxal reductase |
| PGUG_05342 | CFL1 | 1.23 | Ferric reductase transmembrane component |
| PGUG_02968 PGUG_05160 | TRX1 | 2.06 1.36 | Thioredoxin-1 |
| PGUG_05497 | PEX5 | 2.23 | Peroxisomal targeting signal receptor |
| PGUG_04222 | PST2 | 1.49 | Protoplast secreted protein 2 |
| MSTRG.3296 | GRX2 | 1.72 | Glutaredoxin-2, mitochondrial |
| PGUG_03196 | GRX3 | 1.11 | Monothiol glutaredoxin-3 |
| PGUG_02781 | TSA1 | 1.29 | Peroxiredoxin TSA1-A |
| PGUG_02681 | TRR1 | 1.25 | Thioredoxin reductase |
| PGUG_03985 | SNZ1 | 1.03 | Pyridoxal 5 and apos-phosphate synthase subunit |
| PGUG_05341 | SRX1 | 1.87 | Sulfiredoxin |
| PGUG_05768 | NPY1 | 1.57 | NADH pyrophosphatase |
| PGUG_00152 | FLX1 | 1.21 | Mitochondrial FAD carrier protein |
| Cell wall and membrane formation | |||
| PGUG_04330 | YMR244W | 2.62 | Beta-glucosidase (SUN family) |
| PGUG_03333 | HAP1 | 1.13 | Heme-responsive zinc finger transcription factor |
| PGUG_00027 | RBE1 | 1.69 | Repressed by EFG1 protein 1 |
| Growth and reproduction | |||
| PGUG_04612 | MAL31 | 1.32 | Maltose permease |
| PGUG_03041 | FOL2 | 1.04 | GTP cyclohydrolase |
| PGUG_03333 | HAP1 | 1.13 | Heme-responsive zinc finger transcription factor |
| PGUG_04594 | mel1 | 1.81 | Alpha-galactosidase |
| PGUG_01862 | ETR1 | 3.37 | Enoyl-[acyl-carrier-protein] reductase 1, mitochondrial |
| PGUG_04221 | YCP4 | 1.10 | Flavoprotein-like protein |
| PGUG_00387 | LAC12 | 1.69 | Lactose permease |
| PGUG_04842 | nirA | 2.02 | Nitrogen assimilation transcription factor |
| PGUG_03008 | ALD5 | 1.03 | Aldehyde dehydrogenase 5, mitochondrial |
| PGUG_00303 | PKR1 | 1.03 | V-type ATPase assembly factor |
| PGUG_00474 | CCS1 | 1.57 | Superoxide dismutase 1 copper chaperone |
| PGUG_04752 | MIA40 | 1.05 | Mitochondrial intermembrane space import and assembly protein 40 |
| PGUG_02696 | NFU1 | 1.21 | NifU-like protein, mitochondrial |
| PGUG_04929 | COX17 | 1.24 | Cytochrome c oxidase copper chaperone |
| PGUG_04481 | MAS1 | 1.76 | Mitochondrial-processing peptidase subunit beta |
| Transcription and translation regulation | |||
| PGUG_00994 | NOP53 | 1.41 | Ribosome biogenesis protein |
| PGUG_01029 | NBP35 | 1.002 | Cytosolic Fe-S cluster assembly factor |
| PGUG_02131 | CFD1 | 1.80 | Cytosolic Fe-S cluster assembly factor |
| PGUG_01264 | Pch1 | 2.39 | Cyclin pch1 |
| PGUG_01171 | RPL10 | 1.45 | 60S ribosomal protein |
| PGUG_03610 | BUD31 | 1.52 | Pre-mRNA-splicing factor |
| PGUG_04342 | RPC1 | 1.50 | DNA-directed RNA polymerase III subunit |
| PGUG_00512 | SQT1 | 1.48 | Ribosome assembly protein |
| PGUG_04094 | cdc22 | 1.21 | Ribonucleoside-diphosphate reductase large chain |
| PGUG_04921 | MPP10 | 1.31 | U3 small nucleolar RNA-associated protein |
| PGUG_03426 | rnc1 | 1.13 | RNA-binding protein |
| PGUG_02152 | itt1 | 1.01 | E3 ubiquitin-protein ligase |
| PGUG_04056 | SRP40 | 3.12 | Suppressor protein |
| PGUG_03307 | EMG1 | 1.19 | Ribosomal RNA small subunit methyltransferase |
| Transporter | |||
| PGUG_01272 | SPAC10F6.14c | 1.26 | ABC1 family protein |
| PGUG_03561 PGUG_02559 PGUG_01605 PGUG_03562 | YOR1 | 2.20 1.03 1.73 2.22 | Oligomycin resistance ATP-dependent permease |
| PGUG_00888 | YHK8 | 1.96 | Probable drug/proton antiporter |
| PGUG_05418 | ESBP6 | 2.70 | Uncharacterized transporter |
| PGUG_05416 | TNA1 | 1.02 | High-affinity nicotinic acid transporter |
| PGUG_05858 | SEO1 | 1.33 | Probable transporter |
| PGUG_03366 | SPBC947.06c | 1.67 | MFS-type transporter |
| PGUG_01124 | TPO1 | 3.56 | Polyamine transporter 1 |
| PGUG_04286 | TPO1_2 | 1.01 | Multidrug transporter |
| PGUG_00124 | dotC | 1.08 | Efflux pump |
| MSTRG.1806 | zrt1 | 1.64 | Zinc-regulated transporter |
| PGUG_05857 | liz1 | 1.39 | Pantothenate transporter |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Zhang, X.; Solairaj, D.; Fu, Y.; Zhang, H. Molecular Response of Meyerozyma guilliermondii to Patulin: Transcriptomic-Based Analysis. J. Fungi 2023, 9, 538. https://doi.org/10.3390/jof9050538
Yang Q, Zhang X, Solairaj D, Fu Y, Zhang H. Molecular Response of Meyerozyma guilliermondii to Patulin: Transcriptomic-Based Analysis. Journal of Fungi. 2023; 9(5):538. https://doi.org/10.3390/jof9050538
Chicago/Turabian StyleYang, Qiya, Xi Zhang, Dhanasekaran Solairaj, Yu Fu, and Hongyin Zhang. 2023. "Molecular Response of Meyerozyma guilliermondii to Patulin: Transcriptomic-Based Analysis" Journal of Fungi 9, no. 5: 538. https://doi.org/10.3390/jof9050538
APA StyleYang, Q., Zhang, X., Solairaj, D., Fu, Y., & Zhang, H. (2023). Molecular Response of Meyerozyma guilliermondii to Patulin: Transcriptomic-Based Analysis. Journal of Fungi, 9(5), 538. https://doi.org/10.3390/jof9050538