
Diversity and Antimicrobial Activity of Intestinal Fungi from Three Species of Coral Reef Fish



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Fungal Isolation of Fish Intestines and Seawater
2.3. Extraction of Fungal Genomic DNA
2.4. PCR Amplification of Target DNA and Sequencing of ITS Fragments
2.5. Sequence Alignment and Phylogenetic Analyses
2.6. Bioassay of Antimicrobial Activity of Fungal Isolates
2.7. Data Analysis
2.8. Nucleotide Sequence Accession Number
3. Results
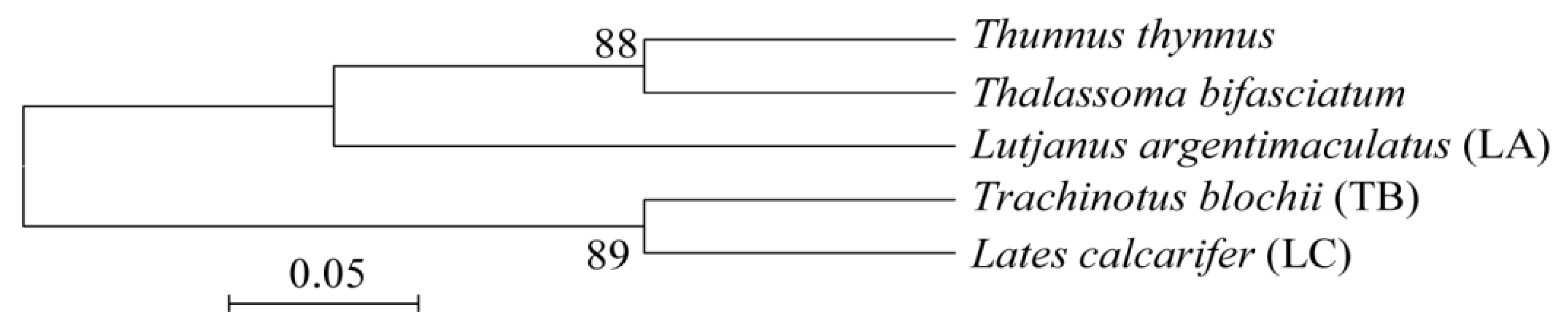
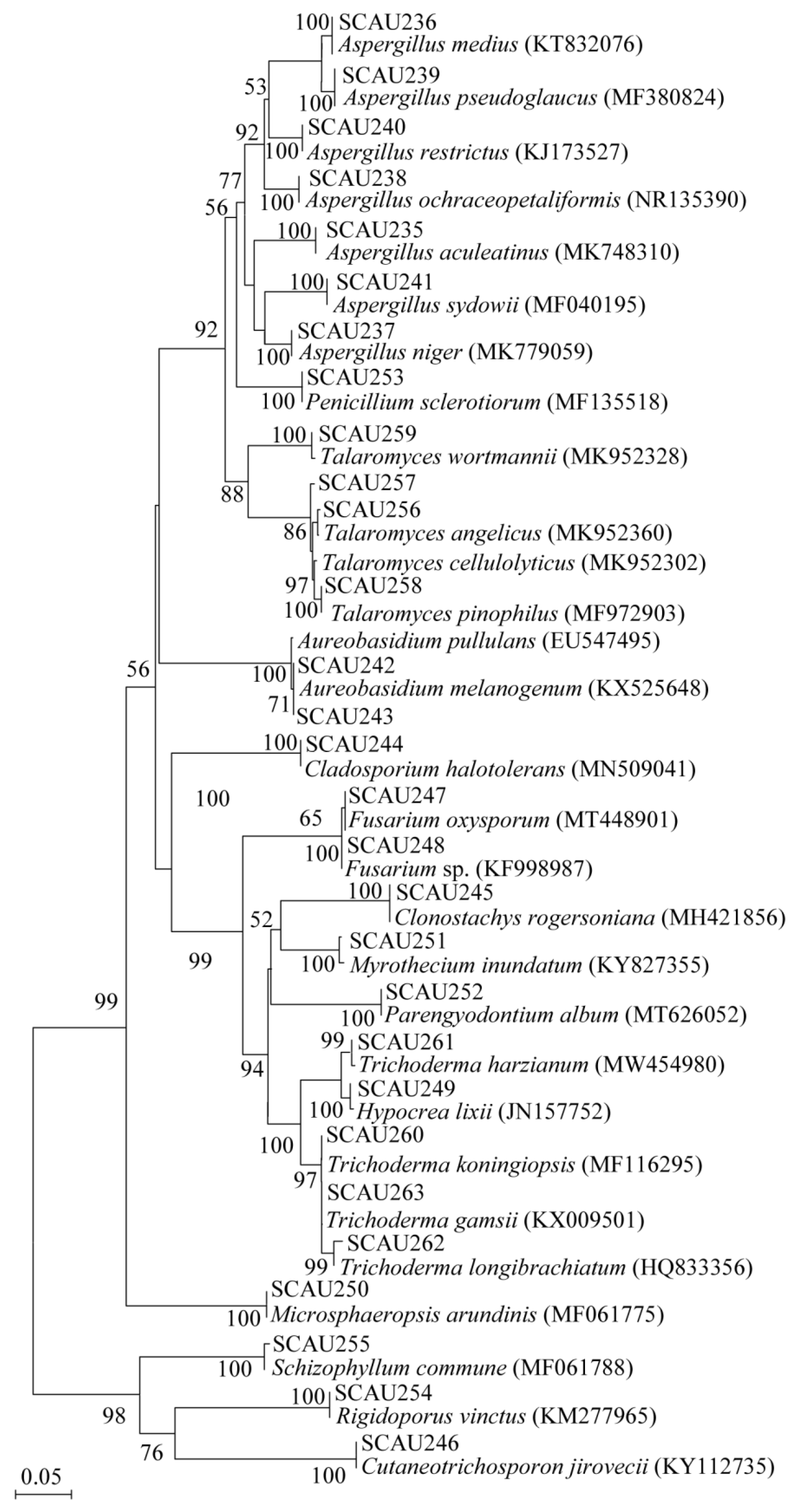
3.1. Isolation and Phylogenetic Diversity of Intestinal Fungi from the Three Coral Reef Fish
3.2. Dissimilarity of Fungal Communities in the Intestines of Three Coral Reef Fish and Seawater
3.3. Comparison of the Fungal Community in Fore-, Mid- and Hind-Intestinal Segments
3.4. Analysis of the Antimicrobial Activity of the Tested Fungi
4. Discussion
4.1. Comparison of Fungal Community in Environments and Intestines from the Three Fish
4.2. Comparison of Fungal Community in Fore-, Mid- and Hind-Intestinal Segments
4.3. Potential Antimicrobial Properties of Fungi from Fish Intestine and Seawater
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, R.; O’Leary, R.A.; Low-Choy, S.; Mengersen, K.; Knowlton, N.; Brainard, R.E.; Caley, M.J. Species richness on coral reefs and the pursuit of convergent global estimates. Curr. Biol. 2015, 25, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.M.; Zou, K.S.; Zhou, L.; Huang, X.D.; Li, Y.Y.; Gao, X.Y.; Chen, X.; Zhang, X.Y. Deep insights into gut microbiota in four carnivorous coral reef fishes from the south china sea. Microorganisms 2020, 8, 426. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.A.J.; Nash, K.L. The importance of structural complexity in coral reef ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- Pandolfi, J.M.; Connolly, S.R.; Marshall, D.J.; Cohen, A.L. Projecting coral reef futures under global warming and ocean acidification. Science 2011, 333, 418–422. [Google Scholar] [CrossRef]
- Brandl, S.J.; Casey, J.M.; Meyer, C.P. Dietary and habitat niche partitioning in congeneric cryptobenthic reef fish species. Coral Reefs 2020, 39, 305–317. [Google Scholar] [CrossRef]
- Kane, C.N.; Tissot, B.N. Trophic designation and live coral cover predict changes in reef-fish community structure along a shallow to mesophotic gradient in hawaii. Coral Reefs 2017, 36, 891–901. [Google Scholar] [CrossRef]
- Ray, A.K.; Ghosh, K.; Ringø, E. Enzyme-producing bacteria isolated from fish gut: A review. Aquac. Nutr. 2012, 18, 465–492. [Google Scholar] [CrossRef]
- Reinoso, S.; Gutierrez, M.S.; Dominguez-Borbor, C.; Arguello-Guevara, W.; Bohorquez-Cruz, M.; Sonnenholzner, S.; Nova-Baza, D.; Mardones, C.; Navarrete, P. Selection of autochthonous yeasts isolated from the intestinal tracts of cobia fish (Rachycentron canadum) with probiotic potential. J. Fungi 2023, 9, 274. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. New Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Tran, N.T.; Zhang, J.; Xiong, F.; Wang, G.T.; Li, W.X.; Wu, S.G. Altered gut microbiota associated with intestinal disease in grass carp (Ctenopharyngodon idellus). World J. Microbiol. Biotechnol. 2018, 34, 71. [Google Scholar] [CrossRef]
- Talwar, C.; Nagar, S.; Lal, R.; Negi, R.K. Fish gut microbiome: Current approaches and future perspectives. Indian J. Microbiol. 2018, 58, 397–414. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Chen, B.; Wang, M.; Wang, S.; Zhang, M.; Sun, Z.; Juventus, A.J.; Ma, H.; Li, Y. Effects of dietary lipid sources on the intestinal microbiome and health of golden pompano (Trachinotus ovatus). Fish Shellfish. Immunol. 2019, 89, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yang, R.; Hu, J.; Lin, S.; Gu, Z.; Ma, Z. The gut microbiota community and antioxidant enzymes activity of barramundi reared at seawater and freshwater. Fish Shellfish Immunol. 2019, 89, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Reshma, K.J.; Sumithra, T.G.; Nair, A.V.; Stefi Raju, V.; Kishor, T.G.; Sreenath, K.R.; Sanil, N.K. An insight into the gut microbiology of wild-caught mangrove red snapper, Lutjanus argentimaculatus (forsskal, 1775). Aquaculture 2018, 497, 320–330. [Google Scholar] [CrossRef]
- Banerjee, G.; Ray, A.K. Bacterial symbiosis in the fish gut and its role in health and metabolism. Symbiosis 2017, 72, 1–11. [Google Scholar] [CrossRef]
- Nie, L.; Zhou, Q.J.; Qiao, Y.; Chen, J. Interplay between the gut microbiota and immune responses of ayu (plecoglossus altivelis) during vibrio anguillarum infection. Fish Shellfish. Immunol. 2017, 68, 479–487. [Google Scholar] [CrossRef]
- Perez, T.; Balcazar, J.L.; Ruiz-Zarzuela, I.; Halaihel, N.; Vendrell, D.; de Blas, I.; Muzquiz, J.L. Host-microbiota interactions within the fish intestinal ecosystem. Mucosal Immunol. 2010, 3, 355–360. [Google Scholar] [CrossRef]
- Schmidt, V.T.; Smith, K.F.; Melvin, D.W.; Amaral-Zettler, L.A. Community assembly of a euryhaline fish microbiome during salinity acclimation. Mol. Ecol. 2015, 24, 2537–2550. [Google Scholar] [CrossRef]
- Zhou, L.; Han, Y.; Wang, D.; Li, Y.; Huang, X.; He, A. Comparison of fungal community composition within different intestinal segments of tilapia and bighead carp. J. Oceanol. Limnol. 2021, 39, 1961–1971. [Google Scholar] [CrossRef]
- Maas, E.; Penders, J.; Venema, K. Fungal-bacterial interactions in the human gut of healthy individuals. J. Fungi 2023, 9, 139. [Google Scholar] [CrossRef]
- Satala, D.; Bras, G.; Kozik, A.; Rapala-Kozik, M.; Karkowska-Kuleta, J. More than just protein degradation: The regulatory roles and moonlighting functions of extracellular proteases produced by fungi pathogenic for humans. J. Fungi 2023, 9, 121. [Google Scholar] [CrossRef] [PubMed]
- Gatesoupe, F.J. Live yeasts in the gut: Natural occurrence, dietary introduction, and their effects on fish health and development. Aquaculture 2007, 267, 20–30. [Google Scholar] [CrossRef]
- Caruffo, M.; Navarrete, N.; Salgado, O.; Díaz, A.; López, P.; García, K.; Feijóo, C.G.; Navarrete, P. Potential probiotic yeasts isolated from the fish gut protect zebrafish (Danio rerio) from a vibrio anguillarum challenge. Front. Microbiol. 2015, 6, 1093. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Becerril, M.; Tovar-Ramírez, D.; Ascencio-Valle, F.; Civera-Cerecedo, R.; Gracia-López, V.; Barbosa-Solomieu, V. Effects of dietary live yeast debaryomyces hansenii on the immune and antioxidant system in juvenile leopard grouper mycteroperca rosacea exposed to stress. Aquaculture 2008, 280, 39–44. [Google Scholar] [CrossRef]
- Rajilić-Stojanović, M.; Heilig, H.G.H.J.; Molenaar, D.; Kajander, K.; Surakka, A.; Smidt, H.; de Vos, W.M. Development and application of the human intestinal tract chip, a phylogenetic microarray: Analysis of universally conserved phylotypes in the abundant microbiota of young and elderly adults. Environ. Microbiol. 2009, 11, 1736–1751. [Google Scholar] [CrossRef]
- Huber, I.; Spanggaard, B.; Appel, K.F.; Rossen, L.; Nielsen, T.; Gram, L. Phylogenetic analysis and in situ identification of the intestinal microbial community of rainbow trout (Oncorhynchus mykiss, walbaum). J. Appl. Microbiol. 2004, 96, 117–132. [Google Scholar] [CrossRef]
- Shang, S.; Ren, J.; Wang, J.; Xin, S.; Xia, J.; Tang, X. High-throughput sequencing reveals significant diversity in the gut microbiomes of humpback (Chanodichthys dabryi) and crucian carp (Carassius carassius). Biologia 2021, 76, 655–662. [Google Scholar] [CrossRef]
- McCulloch, M.; Cappo, M.; Aumend, J.; Müller, W. Tracing the life history of individual barramundi using laser ablation mc-icp-ms sr-isotopic and sr/ba ratios in otoliths. Mar. Freshw. Res. 2005, 56, 637–644. [Google Scholar] [CrossRef]
- Shi, R.; Xu, S.; Qi, Z.; Zhu, Q.; Huang, H.; Weber, F. Influence of suspended mariculture on vertical distribution profiles of bacteria in sediment from daya bay, southern china. Mar. Pollut. Bull. 2019, 146, 816–826. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Hao, H.L.; Lau, S.; Wang, H.Y.; Han, Y.; Dong, L.M.; Huang, R.M. Biodiversity and antifouling activity of fungi associated with two soft corals from the south china sea. Arch. Microbiol. 2019, 201, 757–767. [Google Scholar] [CrossRef]
- Wu, F.; Chen, B.; Liu, S.; Xia, X.; Gao, L.; Zhang, X.; Pan, Q. Effects of woody forages on biodiversity and bioactivity of aerobic culturable gut bacteria of tilapia (Oreochromis niloticus). PLoS ONE 2020, 15, e235560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Zeng, X.N.; Liu, S.; Wu, F.; Li, Y.Y.; Pan, Q. Effects of dietary four different woody forages on gut microbiota of nile tilapia (Oreochromis niloticus). Aquac. Res. 2021, 52, 1733–1742. [Google Scholar] [CrossRef]
- Tejesvi, M.V.; Kajula, M.; Mattila, S.; Pirttilä, A.M. Bioactivity and genetic diversity of endophytic fungi in rhododendron tomentosum harmaja. Fungal Divers. 2011, 47, 97–107. [Google Scholar] [CrossRef]
- Lai, X.; Cao, L.; Tan, H.; Fang, S.; Huang, Y.; Zhou, S. Fungal communities from methane hydrate-bearing deep-sea marine sediments in south china sea. ISME J. 2007, 1, 756–762. [Google Scholar] [CrossRef]
- Cale, J.A.; Scott, N.; Pec, G.J.; Landhausser, S.M.; Karst, J. Choices on sampling, sequencing, and analyzing dna influence the estimation of community composition of plant fungal symbionts. Appl. Plant Sci. 2021, 9, e11449. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.C.K.; Mak, K.K.W.; Chen, F.; Qian, P.Y. Bioactivity of bacterial strains isolated from marine biofilms in Hong Kong waters for the induction of larval settlement in the marine polychaete Hydroides elegans. Mar. Ecol. Prog. Ser. 2002, 226, 301–310. [Google Scholar] [CrossRef]
- Phongpaichit, S.; Preedanan, S.; Rungjindama, N.; Sakayaroj, J.; Benzies, C.; Chuaypat, J.; Plathong, S. Aspergillosis of the gorgonian sea fan Annella sp. After the 2004 tsunami at Mu Ko Similan National Park, Andaman Sea, Thailand. Coral Reefs 2006, 25, 296. [Google Scholar] [CrossRef]
- Ricotta, C.; Podani, J. On some properties of the bray-curtis dissimilarity and their ecological meaning. Ecol. Complex. 2017, 31, 201–205. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Fu, W.; Chen, X.; Yan, M.T.; Huang, X.D.; Bao, J. Phylogenetic analysis and antifouling potentials of culturable fungi in mangrove sediments from techeng isle, china. World J. Microbiol. Biotechnol. 2018, 34, 90. [Google Scholar] [CrossRef]
- Bass, D.; Howe, A.; Brown, N.; Barton, H.; Demidova, M.; Michelle, H.; Li, L.; Sanders, H.; Watkinson, S.C.; Willcock, S.; et al. Yeast forms dominate fungal diversity in the deep oceans. Proc. R. Soc. B-Biol. Sci. 2007, 274, 3069–3077. [Google Scholar] [CrossRef] [PubMed]
- Jebaraj, C.S.; Raghukumar, C.; Behnke, A.; Stoeck, T. Fungal diversity in oxygen-depleted regions of the arabian sea revealed by targeted environmental sequencing combined with cultivation. FEMS Microbiol. Ecol. 2010, 71, 399–412. [Google Scholar] [CrossRef]
- Edgcomb, V.P.; Beaudoin, D.; Gast, R.; Biddle, J.F.; Teske, A. Marine subsurface eukaryotes: The fungal majority. Environ. Microbiol. 2011, 13, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, V.N.; Vitoreli, G.A.; de Menezes, G.; Mendes, C.; Secchi, E.R.; Rosa, C.A.; Rosa, L.H. Taxonomy, phylogeny and ecology of cultivable fungi present in seawater gradients across the northern antarctica peninsula. Extremophiles 2017, 21, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, H.; Gatesoupe, F.J.; She, R.; Lin, Q.; Yan, X.; Li, J.; Li, X. Bacterial signatures of “red-operculum” disease in the gut of crucian carp (Carassius auratus). Microb. Ecol. 2017, 74, 510–521. [Google Scholar] [CrossRef]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev. Aquac. 2018, 10, 626–640. [Google Scholar] [CrossRef]
- Romero, J.; Navarrete, P. 16s rdna-based analysis of dominant bacterial populations associated with early life stages of coho salmon (Oncorhynchus kisutch). Microb. Ecol. 2006, 51, 422–430. [Google Scholar] [CrossRef]
- Li, X.; Yan, Q.; Ringø, E.; Wu, X.; He, Y.; Yang, D. The influence of weight and gender on intestinal bacterial community of wild largemouth bronze gudgeon (Coreius guichenoti, 1874). BMC Microbiol. 2016, 16, 191. [Google Scholar] [CrossRef]
- Ringø, E.; Sperstad, S.; Myklebust, R.; Refstie, S.; Krogdahl, Å. Characterisation of the microbiota associated with intestine of atlantic cod (Gadus morhua L.): The effect of fish meal, standard soybean meal and a bioprocessed soybean meal. Aquaculture 2006, 261, 829–841. [Google Scholar] [CrossRef]
- Le, H.T.M.D.; Lie, K.K.; Giroud-Argoud, J.; Ronnestad, I.; Saele, O. Effects of cholecystokinin (cck) on gut motility in the stomachless fish ballan wrasse (Labrus bergylta). Front. Neurosci. 2019, 13, 553. [Google Scholar] [CrossRef]
- Lin, K.; Wang, W.; Ruan, H.; Dai, J.; Sun, J.; Liu, L.; Huang, X. Transcriptome analysis of differentially expressed genes in the fore- and hind-intestine of ovate pompano Trachinotus ovatus. Aquaculture 2019, 508, 76–82. [Google Scholar] [CrossRef]
- Mongkol, P.; Bunphimpapha, P.; Rungrassamee, W.; Arayamethakorn, S.; Klinbunga, S.; Menasveta, P.; Chaiyapechara, S. Bacterial community composition and distribution in different segments of the gastrointestinal tract of wild-caught adultpenaeus monodon. Aquac. Res. 2018, 49, 378–392. [Google Scholar] [CrossRef]
- Callegari, M.; Crotti, E.; Fusi, M.; Marasco, R.; Gonella, E.; De Noni, I.; Romano, D.; Borin, S.; Tsiamis, G.; Cherif, A.; et al. Compartmentalization of bacterial and fungal microbiomes in the gut of adult honeybees. Npj Biofilms Microbiomes 2021, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, O.G.; Khalil, Z.G.; Capon, R.J. N-amino-l-proline methyl ester from an australian fish gut-derived fungus: Challenging the distinction between natural product and artifact. Mar. Drugs 2021, 19, 151. [Google Scholar] [CrossRef]
- Qi, J.; Chen, C.; He, Y.; Wang, Y. Genomic analysis and antimicrobial components of m7, an aspergillus terreus strain derived from the south china sea. J. Fungi 2022, 8, 1051. [Google Scholar] [CrossRef]
- Gao, H.; Zhou, L.; Cai, S.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Diorcinols b-e, new prenylated diphenyl ethers from the marine-derived fungus aspergillus versicolor zln-60. J. Antibiot. 2013, 66, 539–542. [Google Scholar] [CrossRef]
- Torres-Rodriguez, J.M.; Madrenys-Brunet, N.; Siddat, M.; Lopez-Jodra, O.; Jimenez, T. Aspergillus versicolor as cause of onychomycosis: Report of 12 cases and susceptibility testing to antifungal drugs. J. Eur. Acad. Dermatol. Venereol. 1998, 11, 25–31. [Google Scholar] [CrossRef]
- Miskovic, J.; Karaman, M.; Raseta, M.; Krsmanovic, N.; Berezni, S.; Jakovljevic, D.; Piattoni, F.; Zambonelli, A.; Gargano, M.L.; Venturella, G. Comparison of two schizophyllum commune strains in production of acetylcholinesterase inhibitors and antioxidants from submerged cultivation. J. Fungi 2021, 7, 115. [Google Scholar] [CrossRef]
- Klaus, A.; Kozarski, M.; Niksic, M.; Jakovljevic, D.; Todorovic, N.; Van Griensven, L.J.L.D. Antioxidative activities and chemical characterization of polysaccharides extracted from the basidiomycete schizophyllum commune. Lwt—Food Sci. Technol. 2011, 44, 2005–2011. [Google Scholar] [CrossRef]
- Toyotome, T.; Takino, M.; Takaya, M.; Yahiro, M.; Kamei, K. Identification of volatile sulfur compounds produced by schizophyllum commune. J. Fungi 2021, 7, 465. [Google Scholar] [CrossRef]
- Alam, N.; Cha, Y.J.; Shim, M.J.; Lee, T.S.; Lee, U.Y. Cultural conditions for mycelial growth and molecular phylogenetic relationship in different wild strains of schizophyllum commune. Mycobiology 2010, 38, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Ayukawa, Y.; Komatsu, K.; Taga, M.; Arie, T. Cytological karyotyping of fusarium oxysporum by the germ tube burst method (GTBM). J. Gen. Plant Pathol. 2018, 84, 254–261. [Google Scholar] [CrossRef]
- Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [CrossRef]
- Kulatunga, D.C.; Dananjaya, S.H.; Park, B.K.; Kim, C.H.; Lee, J.; De Zoysa, M. First report of fusarium oxysporum species complex infection in zebrafish culturing system. J. Fish Dis. 2017, 40, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Mallik, S.K.; Shahi, N.; Joshi, N.; Pant, K.; Kala, K.; Chandra, S.; Sarma, D. The emergence of zoonotic fusarium oxysporum infection in captive-reared fingerlings of golden mahseer, tor putitora (hamilton, 1822) from the central himalayan region of india. Transbound. Emerg. Dis. 2020, 67, 555–563. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Fungal Species | Representative Isolates (Accession Number in GenBank) | Similarity of ITS Sequence | Number of Fungal Isolates | |||
|---|---|---|---|---|---|---|
| LC | LA | TB | SW | |||
| Aspergillus aculeatinus | SCAU235 (OK275106) | 99.6% | ||||
| A. medius | SCAU236 (OK275107) | 99.4% | 4 ± 1 | |||
| A. niger | SCAU237 (OK275108) | 100.0% | 1 ± 1 | 9 ± 2 | 5 ± 2 | 7 ± 1 |
| A. ochraceopetaliformis | SCAU238 (OK275109) | 99.8% | 3 ± 2 | |||
| A. pseudoglaucus | SCAU239 (OK275110) | 99.0% | 2 ± 1 | 2 ± 0 | 2 ± 1 | |
| A. restrictus | SCAU240 (OK275111) | 99.1% | 1 ± 1 | |||
| A. sydowii | SCAU241 (OK275112) | 99.3% | 10 ± 3 | 4 ± 1 | ||
| Aureobasidium melanogenum | SCAU242 (OK275113) | 99.6% | 1 ± 0 | 3 ± 1 | ||
| A. pullulans | SCAU243 (OK275114) | 99.8% | 2 ± 1 | 4 ± 1 | 5 ± 2 | |
| Cladosporium halotolerans | SCAU244 (OK275115) | 100.0% | 3 ± 1 | 4 ± 3 | 2 ± 1 | |
| Clonostachys rogersoniana | SCAU245 (OK275116) | 99.6% | 3 ± 1 | |||
| Cutaneotrichosporon jirovecii | SCAU246 (OK275117) | 99.4% | 1 ± 0 | |||
| Fusarium oxysporum | SCAU247 (OK275118) | 99.8% | 2 ± 0 | |||
| Fusarium sp. | SCAU248 (OK275119) | 99.8% | 2 ± 1 | |||
| Hypocrea lixii | SCAU249 (OK275120) | 99.3% | 3 ± 2 | |||
| Microsphaeropsis arundinis | SCAU250 (OK275121) | 99.0% | 2 ± 1 | |||
| Myrothecium inundatum | SCAU251 (OK275122) | 99.67% | 2 ± 1 | |||
| Parengyodontium album | SCAU252 (OK275123) | 99.6% | 2 ± 1 | 2 ± 1 | ||
| Penicillium sclerotiorum | SCAU253 (OK275124) | 99.5% | 2 ± 0 | |||
| Rigidoporus vinctus | SCAU254 (OK275125) | 99.5% | 1 ± 0 | 2 ± 1 | ||
| Schizophyllum commune | SCAU255 (OK275126) | 99.7% | 2 ± 1 | |||
| Talaromyces angelicus | SCAU256 (OK275127) | 98.4% | 1 ± 0 | |||
| T. cellulolyticus | SCAU257 (OK275128) | 98.9% | 2 ± 1 | |||
| T. pinophilus | SCAU258 (OK275129) | 99. 5% | 3 ± 1 | |||
| T. wortmannii | SCAU259 (OK275130) | 99.5% | 3 ± 1 | |||
| Trichoderma gamsii | SCAU260 (OK275131) | 99.7% | 3 ± 1 | |||
| T. harzianum | SCAU261 (OK275132) | 100.0% | 1 ± 0 | |||
| T. koningiopsis | SCAU262 (OK275133) | 99.1% | 2 ± 1 | |||
| T. longibrachiatum | SCAU263 (OK275134) | 99.7% | 6 ± 2 | 2 ± 1 | 3 ± 1 | |
| LC | LA | TB | SW | |
|---|---|---|---|---|
| LC | / | 71.4% | 41.2% | 66.7% |
| LA | 71.4% | / | 60.0% | 81.8% |
| TB | 41.2% | 60.0% | / | 76.0% |
| SW | 66.7% | 81.8% | 76.0% | / |
| LC-F | LC-M | LC-H | LA-F | LA-M | LA-H | TB-F | TB-M | TB-H | |
|---|---|---|---|---|---|---|---|---|---|
| LC-F | / | 27.3% | 71.4% | 100.0% | 60.0% | 100.0% | 75.0% | 66.7% | 100.0% |
| LC-M | 27.3% | / | 100.0% | 81.8% | 45.5% | 80.0% | 77.8% | 71.4% | 81.8% |
| LC-H | 71.4% | 100.0% | / | 100.0% | 100.0% | 100.0% | 60.0% | 100.0% | 100.0% |
| LA-F | 100.0% | 81.8% | 100.0% | / | 60.0% | 33.3% | 75.0% | 100.0% | 20.0% |
| LA-M | 60.0% | 45.5% | 100.0% | 60.0% | / | 55.6% | 75.0% | 66.7% | 60.0% |
| LA-H | 100.0% | 80.0% | 100.0% | 33.3% | 55.6% | / | 71.4% | 100.0% | 33.3% |
| TB-F | 75.0% | 77.8% | 60.0% | 75.0% | 75.0% | 71.4% | / | 100.0% | 75.0% |
| TB-M | 66.7% | 71.4% | 100.0% | 100.0% | 66.7% | 100.0% | 100.0% | / | 100.0% |
| TB-H | 100.0% | 81.8% | 100.0% | 20.0% | 60.0% | 33.3% | 75.0% | 100.0% | / |
| Fungal Species | Representative | Diameter of the Growth Inhibition Zone (mm) | |||||
|---|---|---|---|---|---|---|---|
| Isolates | PP | VA | ML | AV | AS | PC | |
| Aspergillusochraceopetaliformis | SCAU238 | / | 11.3 ± 5.3 | / | / | / | / |
| A. pseudoglaucus | SCAU239-1 | / | 10.8 ± 0.3 | / | / | 19.0 ± 2.0 | 17.0 ± 1.0 |
| A. pseudoglaucus | SCAU239-2 | 12 ± 1.0 | / | / | 20.0 ± 2.3 | / | / |
| A. sydowii | SCAU241 | / | / | / | 20.5 ± 0.5 | / | / |
| Aureobasidium melanogenum | SCAU242 | 13.5 ± 0.5 | 13.5 ± 1.5 | / | / | / | / |
| A. pullulans | SCAU243 | / | / | / | 36.0 ± 1.0 | / | / |
| Cladosporium halotolerans halo tolerans | SCAU244 | 11.5 ± 0.5 | / | / | / | 17.5 ± 0.5 | / |
| Fusarium oxysporum | SCAU247-1 | / | / | / | / | / | 17.0 ± 1.0 |
| F. oxysporum | SCAU247-2 | 14.5 ± 2.5 | / | / | / | / | / |
| F. oxysporum | SCAU247-3 | 14.5 ± 0.5 | 15.5 ± 3.5 | / | / | / | 20.5 ± 0.5 |
| Hypocrea lixii | SCAU249 | 12.0 ± 0.3 | 13.0 ± 1.0 | 18.0 ± 4.0 | / | / | / |
| Microsphaeropsis arundinis | SCAU250 | / | / | / | 22.5 ± 0.5 | / | 17.5 ± 1.5 |
| Parengyodontium album | SCAU252 | 13.5 ± 1.5 | 13.5 ± 2.5 | / | / | / | / |
| Penicillium sclerotiorum | SCAU253 | / | / | / | / | 15.5 ± 0.5 | 19.5 ± 2.5 |
| Schizophyllum commune | SCAU255 | 12.5 ± 0.5 | 13.3 ± 2.7 | 13.0 ± 1.0 | / | / | 26.5 ± 3.5 |
| Talaromyces cellulolyticus | SCAU257 | / | / | / | 17.8 ± 2.8 | / | 12.5 ± 0.5 |
| T. wortmannii | SCAU259 | / | / | / | 26.0 ± 4.0 | / | 18.5 ± 1.5 |
| T. gamsii | SCAU260 | / | 8.3 ± 3.9 | / | / | / | / |
| T. harzianum | SCAU261 | / | / | 20.5 ± 5.5 | / | / | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, X.; Yang, J.; Zhou, Z.; Wu, J.; Xu, D.; Yang, Q.; Zhong, S.; Zhang, X. Diversity and Antimicrobial Activity of Intestinal Fungi from Three Species of Coral Reef Fish. J. Fungi 2023, 9, 613. https://doi.org/10.3390/jof9060613
Liao X, Yang J, Zhou Z, Wu J, Xu D, Yang Q, Zhong S, Zhang X. Diversity and Antimicrobial Activity of Intestinal Fungi from Three Species of Coral Reef Fish. Journal of Fungi. 2023; 9(6):613. https://doi.org/10.3390/jof9060613
Chicago/Turabian StyleLiao, Xinyu, Jiadenghui Yang, Zanhu Zhou, Jinying Wu, Dunming Xu, Qiaoting Yang, Saiyi Zhong, and Xiaoyong Zhang. 2023. "Diversity and Antimicrobial Activity of Intestinal Fungi from Three Species of Coral Reef Fish" Journal of Fungi 9, no. 6: 613. https://doi.org/10.3390/jof9060613
APA StyleLiao, X., Yang, J., Zhou, Z., Wu, J., Xu, D., Yang, Q., Zhong, S., & Zhang, X. (2023). Diversity and Antimicrobial Activity of Intestinal Fungi from Three Species of Coral Reef Fish. Journal of Fungi, 9(6), 613. https://doi.org/10.3390/jof9060613