
New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collecting and Site Description
2.2. Morphological Study
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Alignment and Phylogenetic Analyses
3. Results
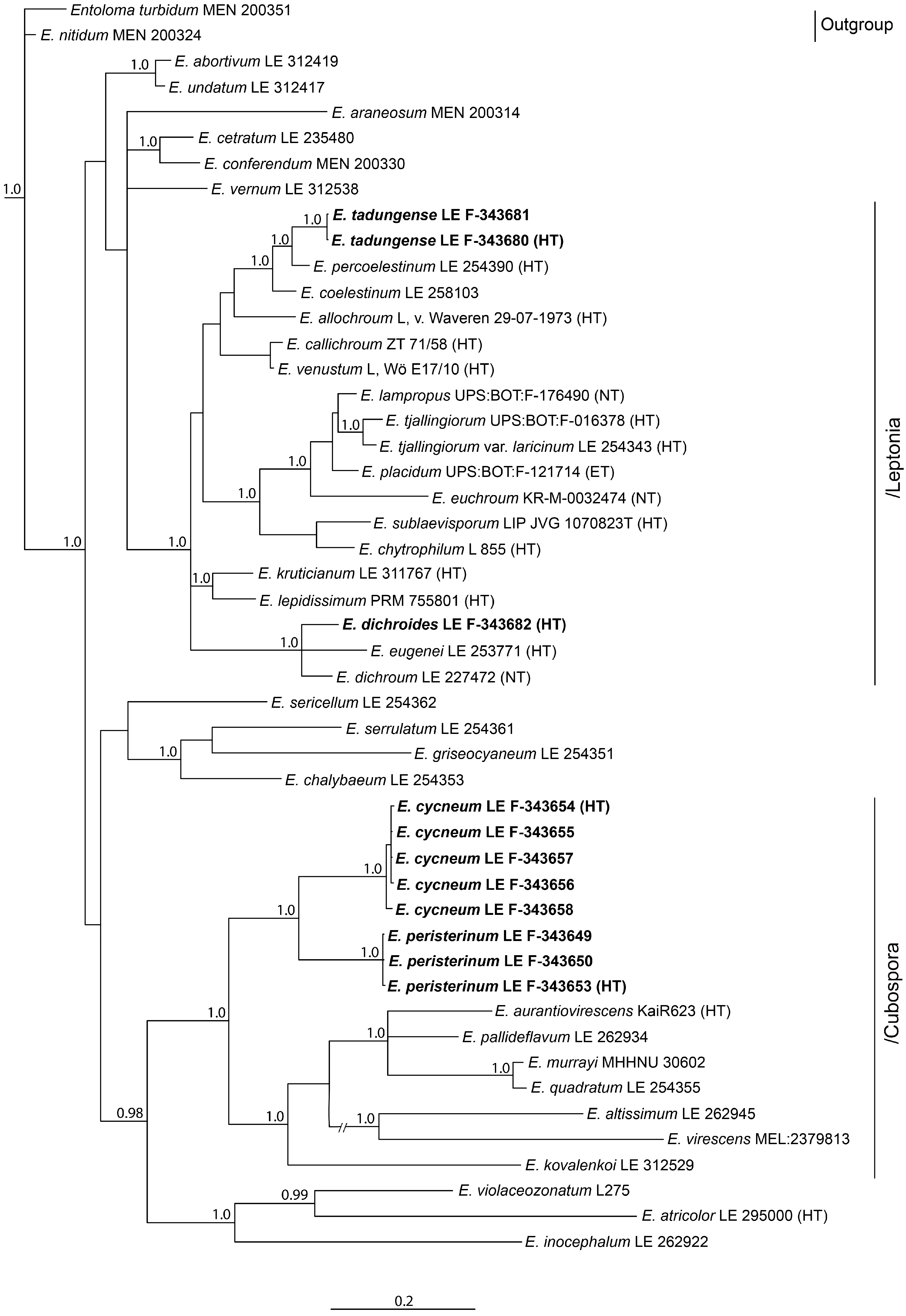
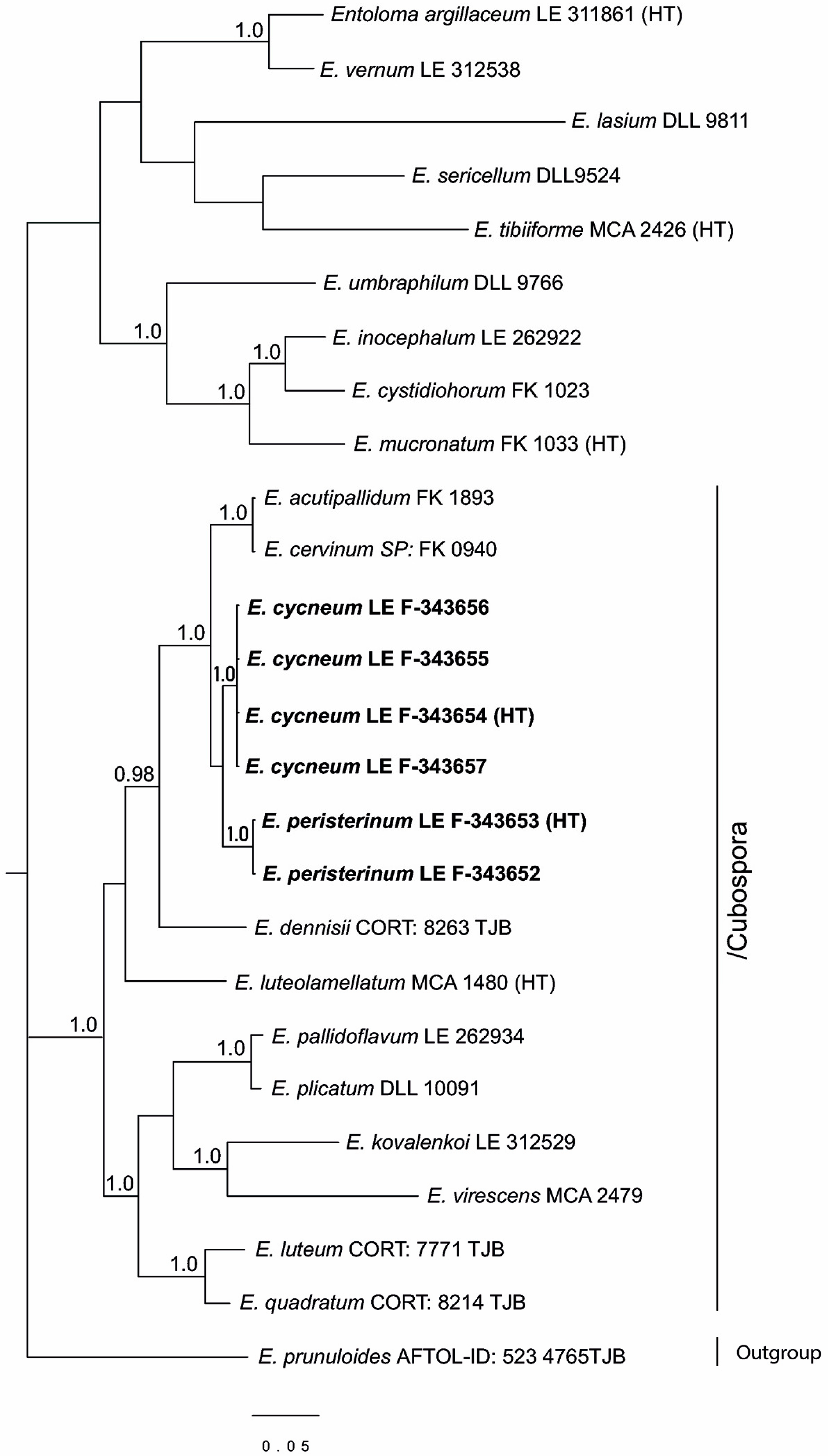
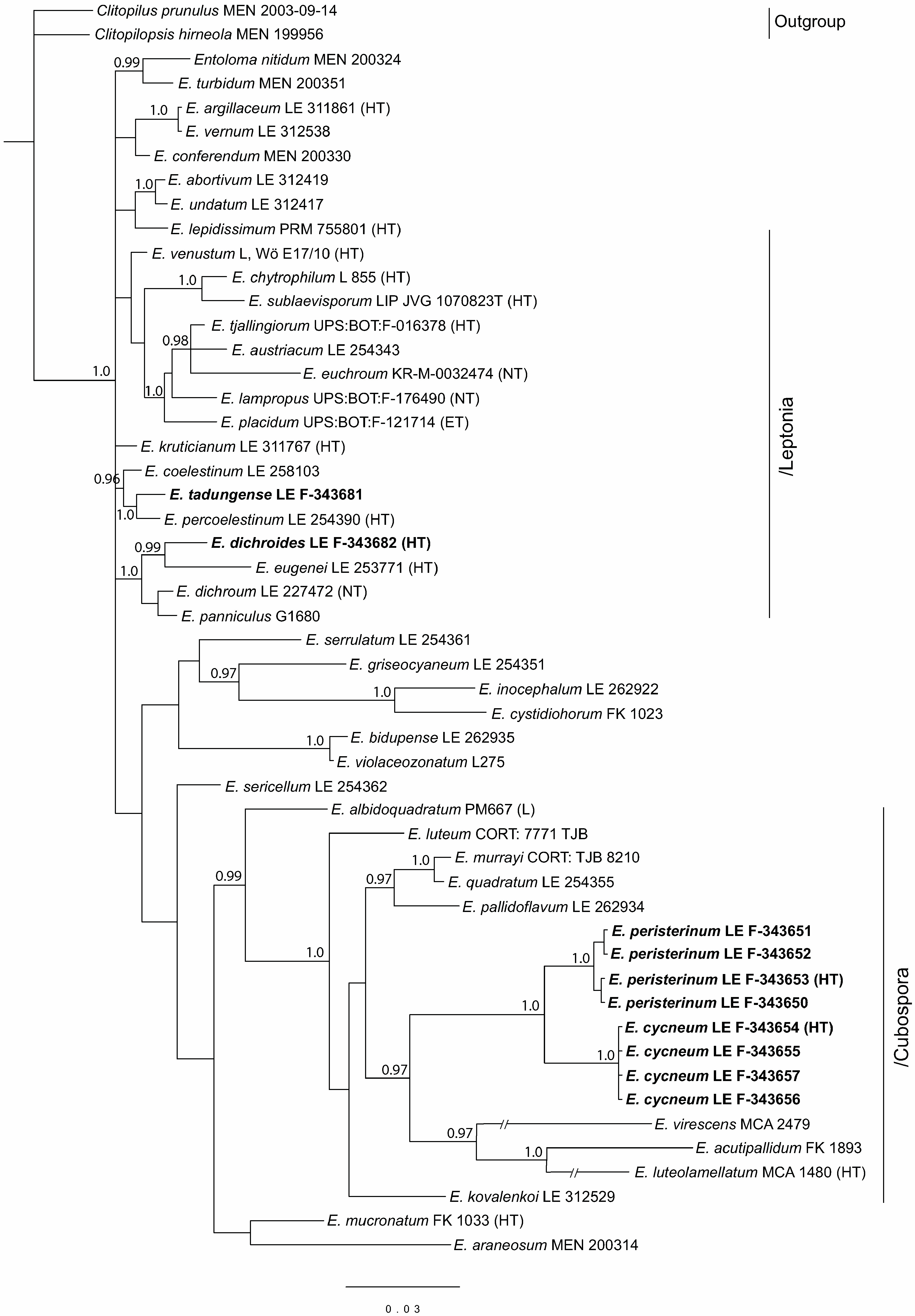
3.1. Phylogenetic Analysis
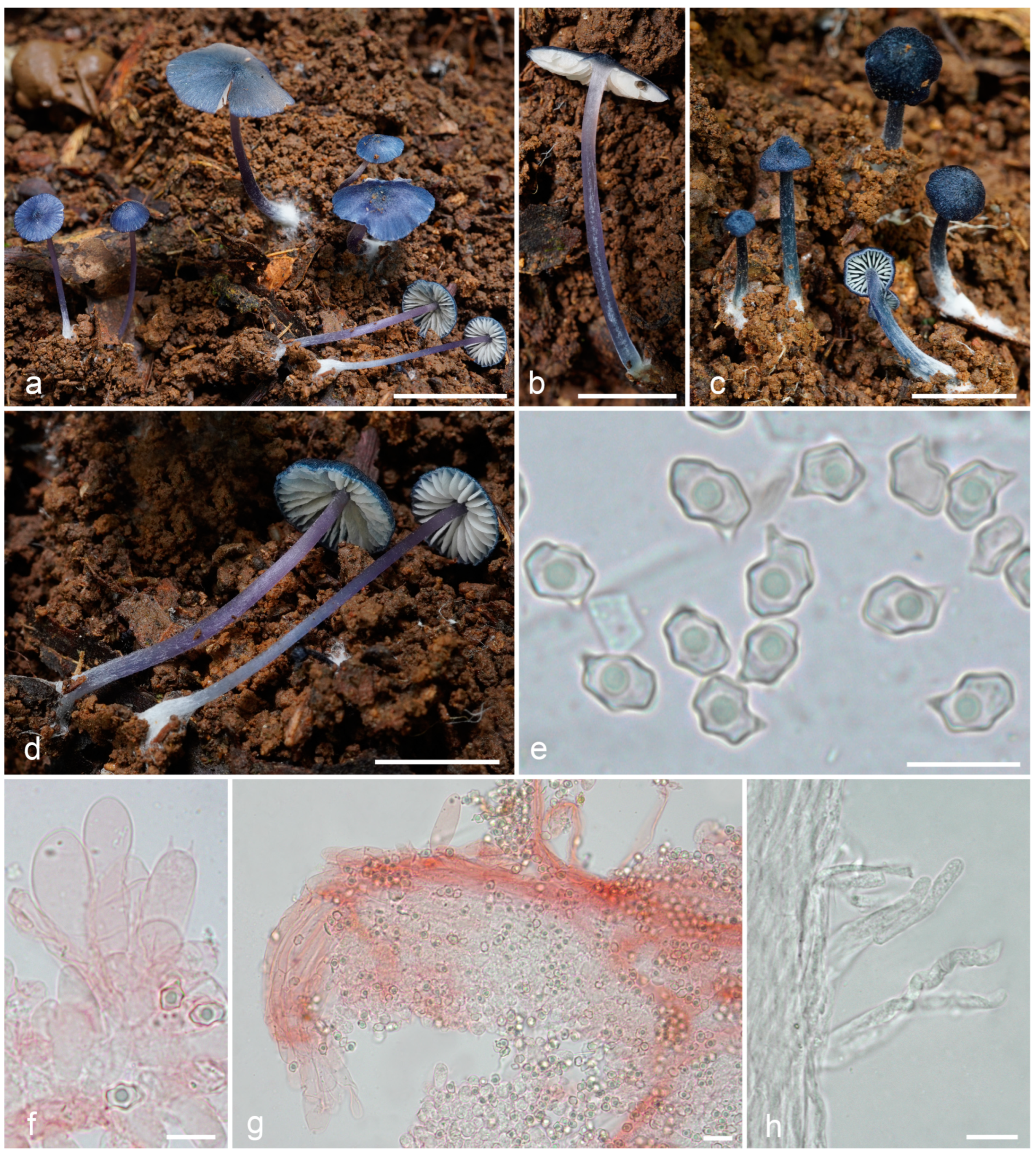
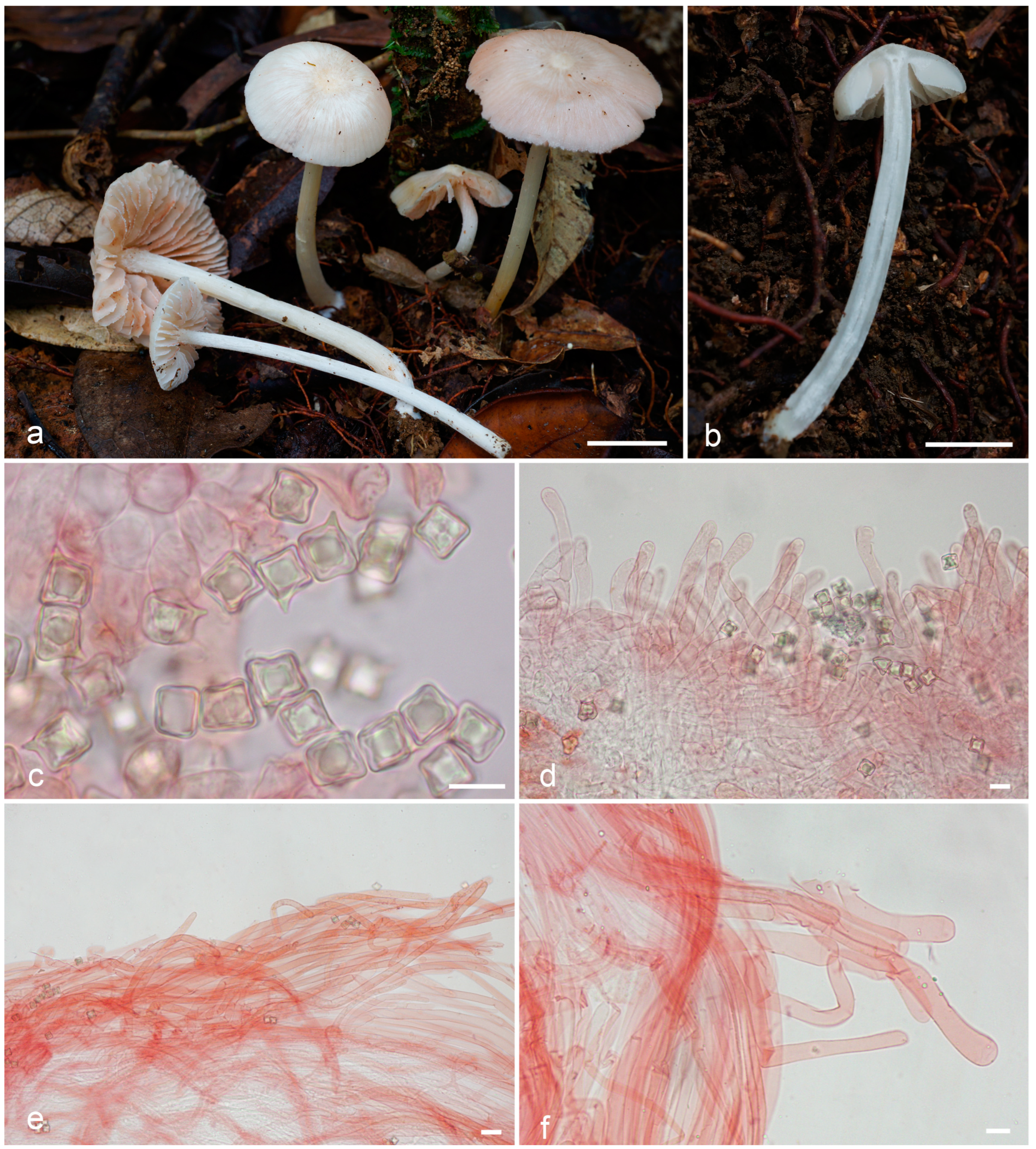
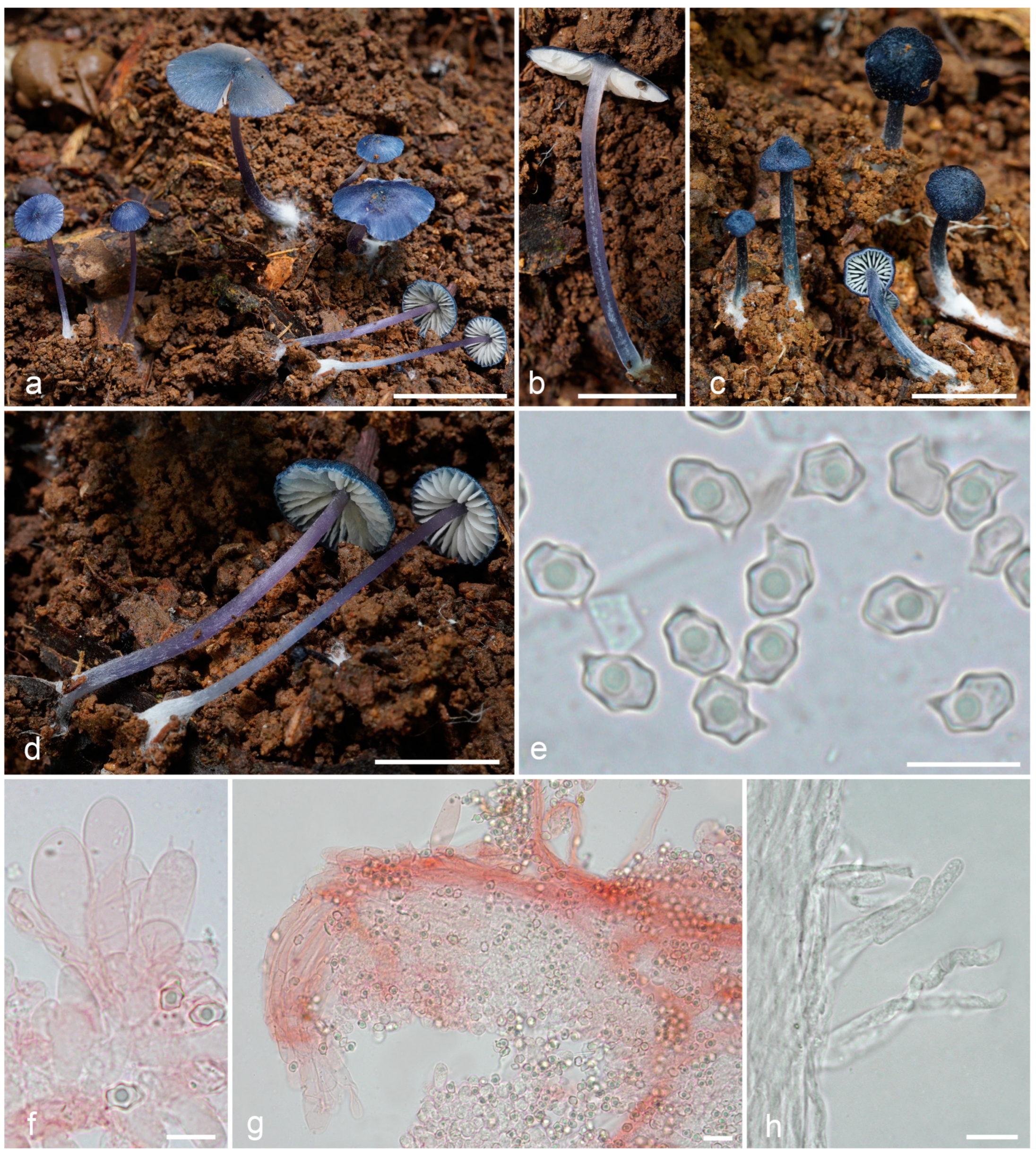
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pham, T.H.G.; Popov, E.S.; Morozova, O.V.; Nguyen, D.H. Basidiomycetous Macromycetes of the Tay Nguyen Plateau in Vietnam: The History of Investigation. Mikol. Fitopatol. 2018, 52, 306–318. [Google Scholar]
- Haelewaters, D.; Dima, B.; Abdel-Hafiz, B.I.I.; Abdel-Wahab, M.A.; Abul-Ezz, S.R.; Acar, I.; Aguirre-Acosta, E.; Aime, M.C.; Aldemir, S.; Ali, M.; et al. Fungal Systematics and Evolution 6. Sydowia 2020, 72, 231–356. [Google Scholar]
- Crous, P.W.; Osieck, E.R.; Jurjevi, Ž.; Boers, J.; Van Iperen, A.L.; Starink-Willemse, M.; Dima, B.; Balashov, S.; Bulgakov, T.S.; Johnston, P.R.; et al. Fungal Planet description sheets: 1284–1382. Persoonia 2021, 47, 178–374. [Google Scholar] [CrossRef]
- Morozova, O.; Popov, E.; Alexandrova, A.; Pham, T.H.G.; Noordeloos, M.E. Four new species of Entoloma (Entolomataceae, Agaricomycetes) subgenera Cyanula and Claudopus from Vietnam and their phylogenetic position. Phytotaxa 2022, 549, 1–21. [Google Scholar] [CrossRef]
- Co-David, D.; Langeveld, D.; Noordeloos, M.E. Molecular phylogeny and spore evolution of Entolomataceae. Persoonia 2009, 23, 147–176. [Google Scholar] [CrossRef]
- Baroni, T.J.; Matheny, P.B. A re-evaluation of gasteroid and cyphelloid species of Entolomataceae from eastern North America. Harv. Pap. Bot. 2011, 16, 293–310. [Google Scholar] [CrossRef]
- Karstedt, F.; Baroni, T.J.; Largent, D.L.; Capelari, M.; Bergemann, S.E. Phylogenetic and morphological analyses of species of the Entolomataceae (Agaricales, Basidiomycota) with cuboid basidiospores. Phytotaxa 2019, 391, 1–27. [Google Scholar] [CrossRef]
- Noordeloos, M.E.; Morozova, O.; Dima, B.; Reschke, K.; Jansen, G.M.; Brandrud, T.E.; Jordal, J.B.; Bendiksen, E.; Vila, J. Entoloma Sensu Lato. Subgenera Cyanula, Leptonia, Nolanea, Trichopilus, and the/Rhombisporum Clade. Fungi Europaei; Edizione Candusso: Alassio, Italy, 2022; Volume 5b, pp. 1–968. [Google Scholar]
- Noordeloos, M.E.; Jansen, G.M.; Jordal, J.B.; Morozova, O.; Weholt, Ø.; Brandrud, T.E.; Vila, J.; Karich, A.; Reschke, K.; Lorås, J.; et al. Molecular and morphological diversity in the /Rhombisporum clade of the genus Entoloma with a note on E. cocles. Mycol. Prog. 2022, 21, 48. [Google Scholar] [CrossRef]
- Romagnesi, H. Les Rhodophylles de Madagascar (Entoloma, Nolanea, Leptonia, Eccilia, Claudopus). Prodrome Flore Mycol. Madag. 1941, 2, 1–164. [Google Scholar]
- Morozova, O.V.; Popov, E.S.; Kovalenko, A.E. Studies on mycobiota of Vietnam. I. Genus Entoloma: New records and new species. Mikol. Fitopatol. 2012, 46, 184–200. [Google Scholar]
- He, X.-L.; Horak, E.; Li, T.-H.; Peng, W.-H.; Gan, B.-C. Two new cuboid-spored species of Entoloma s.l. (Agaricales, Entolomataceae) from Southern China. Cryptogam. Mycol. 2015, 36, 237–249. [Google Scholar] [CrossRef]
- Horak, E. On cuboid spored species of Entoloma. Sydowia 1976, 28, 171–236. [Google Scholar]
- Horak, E. Additions to ‘On cuboid spored species of Entoloma’. Sydowia 1977, 29, 289–299. [Google Scholar]
- Horak, E. Entoloma in South America. I. Sydowia 1978, 30, 40–110. [Google Scholar]
- Horak, E. Entoloma (Agaricales) in Indomalaya and Australasia. Nova Hedwig. Beih. 1980, 65, 1–352. [Google Scholar]
- Horak, E. Entoloma in South America. II. Sydowia 1982, 35, 75–99. [Google Scholar]
- Pegler, D.N. Agaric Flora of Sri Lanka. Kew Bull. Addit. Ser. 1986, 12, 1–519. [Google Scholar]
- Manimohan, P.; Joseph, A.V.; Leelavathy, K.M. The genus Entoloma in Kerala State, India. Mycol. Res. 1995, 99, 1083–1097. [Google Scholar] [CrossRef]
- Manimohan, P.; Noordeloos, M.E.; Dhanya, A.M. Studies on the genus Entoloma (Basidiomycetes, Agaricales) in Kerala State, India. Persoonia 2006, 19, 45–93. [Google Scholar]
- Noordeloos, M.E.; Morozova, O.V. New and noteworthy Entoloma species from the Primorsky Territory, Russian Far East. Mycotaxon 2010, 112, 231–255. [Google Scholar] [CrossRef]
- Romagnesi, H.; Gilles, G. Les Rhodophylles des forêts côtières du Gabon et de la Côte d’Ivoire. Nova Hedwig. Beih. 1979, 59, 1–649. [Google Scholar]
- Eyssartier, G.; Buyck, B.; Courtecuisse, R. New species and combinations in cuboid-spored Entoloma species from Madagascar. Mycol. Res. 2001, 105, 1144–1148. [Google Scholar] [CrossRef]
- Noordeloos, M.E.; Hausknecht, A. The genus Entoloma (Basidiomycetes, Agaricales) of the Mascarenes and Seychelles. Fungal Divers. 2007, 27, 111–144. [Google Scholar]
- Horak, E. Agaricales of New Zealand 1: Pluteaceae (Pluteus, Volvariella); Entolomataceae (Claudopus, Clitopilus, Entoloma, Pouzarella, Rhodocybe, Richoniella). Fungal Divers. Res. Ser. 2008, 19, 1–305. [Google Scholar]
- Noordeloos, M.E.; Gates, G.M. The Entolomataceae of Tasmania. Fungal Divers. Res. Ser. 2012, 22, 1–400. [Google Scholar] [CrossRef]
- Horak, E.; Cheype, J.-L. Entoloma acutopallidum et Entoloma pileofribrosum deux espèces nouvelles de la Guyane française. Bull. Trimest. Soc. Mycol. Fr. 2008, 124, 287–298. [Google Scholar]
- Karstedt, F.; Capelari, M. Inocephalus (Entolomataceae, Agaricales) from São Paulo State, Brazil. Nova Hedwig. 2013, 96, 279–308. [Google Scholar] [CrossRef]
- Karstedt, F.; Capelari, M. A new species of Entolomataceae with cuboidal basidiospores from the São Paulo Metropolitan Region, Brazil. Mycosphere 2015, 6, 69–73. [Google Scholar] [CrossRef]
- Pegler, D.N. Agaric flora of the Lesser Antilles. Kew Bull. Addit. Ser. 1983, 9, 1–668. [Google Scholar]
- Hesler, L.R. Entoloma in Southeastern North America. Nova Hedwig. Beih. 1967, 23, 1–196. [Google Scholar]
- Largent, D.L. Entolomatoid Fungi of the Western United States and Alaska; Mad River Press: Eureka, CA, USA, 1994; pp. 1–495. [Google Scholar]
- Largent, D.L. The genus Leptonia on the Pacific Coast of the United States including a study of North American types. Bibl. Mycol. 1977, 55, 1–286. [Google Scholar]
- Noordeloos, M.E. Entoloma subgenus Leptonia in northwestern Europe–I. Introduction and a revision of its section Leptonia. Persoonia 1982, 11, 451–471. [Google Scholar]
- Noordeloos, M.E. Entoloma Sensu Lato. Fungi Europaei; Giovanna Biella: Saronno, Italy, 1992; Volume 5, pp. 1–760. [Google Scholar]
- Noordeloos, M.E. Entoloma Sensu Lato. Fungi Europaei; Edizione Candusso: Alassio, Italy, 2004; Volume 5a, pp. 761–1378. [Google Scholar]
- Vidal, J.M.; Bellanger, J.-M.; Moreau, P.-A. Three new gasteroid species of the genus Entoloma found in Spain. Bol. Micol. FAMCAL 2016, 11, 53–78. [Google Scholar]
- Reschke, K.; Morozova, O.V.; Dima, B.; Dima, B.; Cooper, J.A.; Corriol, G.; Biketova, A.Y.; Piepenbring, M.; Noordeloos, M.E. Phylogeny, taxonomy, and character evolution in Entoloma subgenus Nolanea. Persoonia 2022, 49, 136–170. [Google Scholar] [CrossRef]
- Morozova, O.V.; Noordeloos, M.E.; Vila, J. Entoloma subgenus Leptonia in boreal-temperate Eurasia: Towards a phylogenetic species concept. Persoonia 2014, 32, 141–169. [Google Scholar] [CrossRef]
- New, M.; Lister, D.; Hulme, M.; Makin, I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 21, 1–25. [Google Scholar] [CrossRef]
- Tordoff, A.W.; Tran, Q.B.; Nguyen, D.T.; Le, M.H. (Eds.) Sourcebook of Existing and Proposed Protected Areas in Vietnam, 2nd ed.; CD-ROOM; Birdlife International in Indochina and Ministry of Agriculture and Rural Development: Hanoi, Vietnam, 2004; Available online: https://thiennhienviet.org.vn/sourcebook/source_book/index_EN.html (accessed on 15 December 2022).
- Morozova, O.V.; Noordeloos, M.E.; Popov, E.S.; Alexandrova, A.V. Three new species within the genus Entoloma (Basidiomycota, Agaricales) with clamped basidia and a serrulatum-type lamellae edge, and their phylogenetic position. Mycol. Prog. 2018, 17, 381–392. [Google Scholar] [CrossRef]
- Pham, T.H.G.; Morozova, O.V.; Alexandrova, A.V. Boletoid fungi (Boletaceae, Basidiomycota) of protected areas of Kon Tum Plateau (Central Highlands of Vietnam). Turczaninowia 2021, 24, 65–76. Available online: http://turczaninowia.asu.ru/article/view/10442 (accessed on 15 December 2022). [CrossRef]
- Nguyen, T.T. Study on structural characteristics, regeneration and determination of natural forest resilience in Ta Dung National Park, Dak Nong province. Summ. Rep. Sci. Technol. Top. 2021, 22–36. (In Vietnamese) [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; pp. 1–252. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-a sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Reschke, K.; Noordeloos, M.E.; Manz, C.; Hofmann, T.A.; Rodriguez-Cedeno, J.; Dima, B.; Piepenbring, M. Fungal diversity in the tropics: Entoloma spp. in Panama. Mycol. Progress 2022, 21, 93–145. [Google Scholar] [CrossRef]
- Morgado, L.N.; Noordeloos, M.E.; Lamoureux, Y.; Geml, J. Multi-gene phylogenetic analyses reveal species limits, phylogeographic patterns, and evolutionary histories of key morphological traits in Entoloma (Agaricales, Basidiomycota). Persoonia 2013, 31, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M.; Leroux, J.J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.W.J.; Heykoop, M.; et al. Fungal Planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef]
- Chen, Z.H.; Zhang, P. Atlas of Macrofungi in Hunan; Hunan Normal University Press: Changsha, China, 2019; pp. 1–426. (In Chinese) [Google Scholar]
- Varga, T.; Krizsán, K.; Földi, C.; Dima, B.; Sánchez-García, M.; Sánchez-Ramírez, S.; Szöllősi, G.J.; Szarkándi, J.G.; Papp, V.; Albert, L.; et al. Megaphylogeny resolves global patterns of mushroom evolution. Nat. Ecol. Evol. 2019, 3, 668–678. [Google Scholar] [CrossRef]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.M.; Ge, Z.W.; Yang, Z.L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 984–997. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008; p. 771. [Google Scholar] [CrossRef]
- Patouillard, N.; Demange, V. Nouvelles contributions’a la flore mycologique du Tonkin. Bull. Soc. Mycol. Fr. 1910, 26, 31–48. [Google Scholar]
- Patouillard, N. Quelques champignons du Tonkin. Bull. Soc. Mycol. Fr. 1913, 29, 206–228. [Google Scholar]
- Heim, R.; Malençon, G. Champignons du Tonkin recueillies par M. V. Demange. Ann. Cryptogam. Exot. 1928, 1, 58–74. [Google Scholar]
- Kiet, T.T. Preliminary checklist of macrofungi of Vietnam. Feddes Reper. 1998, 109, 257–277. [Google Scholar] [CrossRef]
- Le, T.C. (Ed.) Danh lục các loài thực vật Việt Nam: Tập 1 [List of Plant Species in Vietnam: Volume 1]; Agriculture Publishing House: Hanoi, Vietnam, 2001; pp. 1–1183. (In Vietnamese) [Google Scholar]
- Le, B.D. Nấm lớn ở Tây Nguyên [Macrofungi of the Central Highlands]; Science and Technics Publishing House: Hanoi, Vietnam, 2003; pp. 1–261. (In Vietnamese) [Google Scholar]
- Ngo, A. Sự đa dạng của khu hệ nấm lớn ở VQG Bạch Mã, tỉnh Thừa Thiên Huế [Diversity of macrofungi in Bach Ma National Park, Thua Thien Hue province. J. Res. Dev. 2006, 1, 46–52. [Google Scholar]
- Ngo, A.; Nguyen, T.K.C. Sự đa dạng nấm lớn khu bảo tồn thiên nhiên Phong Điền tỉnh Thừa Thiên Huế. Hội nghị Khoa học toàn quốc về Sinh thái và Tài nguyên sinh vật lần thứ 5. [Diversity of macrofungi in Phong Dien Nature Reserve, Thua Thien Hue Province]. In Proceedings of the 5th National Science Conference on Ecology and Biological Resources, Hanoi, Vietnam, 18 October 2013; pp. 346–370. (In Vietnamese). [Google Scholar]
- Kiet, T.T. Macrofungi of Vietnam. V. 1; Science and Technics Publishing House: Hanoi, Vietnam, 2011; pp. 1–314. (In Vietnamese) [Google Scholar]
- Kiet, T.T. Macrofungi of Vietnam. V. 2; Science and Technics Publishing House: Hanoi, Vietnam, 2012; pp. 1–413. (In Vietnamese) [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Hernández-Restrepo, M.; Sutton, D.A.; Acharya, K.; Groenewald, J.Z. Fungal Planet description sheets: 320–370. Persoonia 2015, 34, 167–266. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy GS, J.; Smith, D.; Groenewald, J.Z. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef]
- Morozova, O.V.; Noordeloos, M.E.; Popov, E.S.; Alexandrova, A.V. Nomenclatural novelties. Index Fungorum 2020, 459, 1. [Google Scholar]
- Dörfelt, H.; Kiet, T.T.; Berg, A. Neue Makromyceten-Kollektionen von Vietnam und deren systematische und okogeographische Bedeutung. Feddes Repert. 2004, 115, 164–177. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Voucher Number | Genbank Accession No. | References | ||
|---|---|---|---|---|---|---|
| ITS | LSU | tef | ||||
| Clitopilopsis hirneola (Fr.) Kühner (Clitopilus hirneolus (Fr.) Kühner & Romagn.) | Italy | MEN 199956 | — | GQ289211 | — | [5] |
| Clitopilus prunulus (Scop.) P. Kumm. | Belgium | MEN 2003-09-14 | — | GQ289149 | — | [5] |
| Entoloma abortivum (Berk. & M.A. Curtis) Donk | Russia: Far East | LE 312419 | MF476905 | MF487792 | — | [42] |
| E. acutipallidum E. Horak & Cheype | Brazil: Pará | SP: FK1893 | — | MG018325 | MH190147 | [7] |
| E. albidoquadratum Manim. & Noordel. | India: Kerala | PM667 (L) | — | GQ289151 | — | [5] |
| E. allochroum Noordel. | The Netherlands | v. Waveren, 29-07-1973, holotype (L) | KC898372 | — | — | [39] |
| E. altissimum (Massee) E. Horak | Vietnam | LE 262945 | MF476912 | — | — | [42] |
| E. argillaceum O.V. Morozova et al. | Russia: Caucasus | LE 311861, holotype | — | OL338531 | OL405537 | [38] |
| E. araneosum (Quél.) M.M. Moser | Belgium | MEN 200314 | KC710056 | GQ289153 | — | [5] |
| E. atricolor O.V. Morozova et al. | Vietnam | LE 295000, holotype | KY777496 | — | — | [42] |
| E. aurantiovirescens Reschke, Lotz-Winter & Noordel. | Panama | KaiR623, holotype | MZ611665 | — | — | [50] |
| E. austriacum Courtec. [E. tjallingiorum var. laricinum O.V. Morozova et al.] | Russia: Far East | LE 254343, holotype | KC898413 | KC898513 | — | [39] |
| E. bidupense O.V. Morozova & E.S. Popov | Vietnam | LE 262935 | — | NG_059265 | — | [42] |
| E. callichroum E. Horak & Noordel. | Switzerland | ZT 71/58, holotype | KC898350 | — | — | [39] |
| E. cervinum (Karstedt & Capelari) Blanco-Dios [Inocephalus cervinus Karstedt & Capelari] | Brazil | SP: FK 0940 | — | — | MH190138 | [7] |
| E. cetratum (Fr.) M.M. Moser | Russia: European part | LE 235480 | KC898450 | — | — | [39] |
| E. chalybaeum (Pers.) Noordel. | Russia: European part | LE 254353 | KC898445 | KC898500 | — | [39] |
| E. chytrophilum Wölfel, Noordel. & Dähncke | Spain: Canary Islands | L 855, holotype | KC898434 | KC898519 | — | [39] |
| E. coelestinum (Fr.) Hesler | Russia: Ural | LE 258103 | KC898362 | KC898524 | — | [39] |
| E. conferendum (Britzelm.) Noordel. | MEN 200330 | KC710055 | KC710133 | — | [51] | |
| E. cycneum O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343654, holotype | OQ779461 | OQ804518 | OQ779183 | this work |
| E. cycneum | Vietnam | LE F-343655 | OQ779463 | OQ804519 | OQ779182 | this work |
| E. cycneum | Vietnam | LE F-343656 | OQ779462 | OQ804521 | OQ779181 | this work |
| E. cycneum | Vietnam | LE F-343657 | OQ779464 | OQ804520 | OQ779184 | this work |
| E. cycneum | Vietnam | LE F-343658 | OQ779465 | — | OQ779185 | this work |
| E. cystidiophorum Dennis [Inocephalus cystidiophorus (Dennis) Karstedt & Capelari] | Brazil: São Paulo | SP: FK1023 | — | MG018319 | MH190140 | [7] |
| E. dennisii E. Horak [Inocephalus dennisii (E. Horak) Karstedt & Capelari] | Puerto Rico | CORT: 8263 TJB | — | — | MH190164 | [7] |
| E. dichroides O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343682 | OQ779472 | OQ804527 | — | this work |
| E. dichroum (Pers.) P. Kumm. | Russia: European part | LE 227472, neotype | KC898440 | — | — | [39] |
| E. dichroum | Russia: European part | LE 234260 | — | KC898527 | — | [39] |
| E. euchroum (Pers.) Donk | Russia: Caucasus | LE 262995 | KC898417 | KC898516 | — | [39] |
| E. eugenei Noordel. & O.V. Morozova | Russia: Primorsky Territory | LE 253771, holotype | KC898438 | KC898529 | — | [39] |
| E. griseocyaneum (Fr.) P. Kumm. | Russia: Caucasus | LE 254351 | KC898444 | KC898498 | — | [39] |
| E. inocephalum (Romagn.) Dennis | Vietnam | LE 262922 | KC898449 | MH259311 | MH190154 | [7,39] |
| E. kovalenkoi O.V. Morozova, E.S. Popov & A.V. Alexandrova | Vietnam | LE 312529 | OK257210 | OK257207 | OK256169 | [3] |
| E. kruticianum O.V. Morozova, M.Y. Dyakov, E.S. Popov & A.V. Alexandrova | Russia: European part | LE 311767, holotype | KU666558 | KU710222 | — | [52] |
| E. lampropus (Fr.) Hesler | Sweden | UPS:BOT:F-176490, neotype | KC898377 | KC898506 | — | [39] |
| E. lasium (Berk. & Broome) Noordel. & Co-David [Pouzarella lasia (Berk. & Broome) Largent & Abell-Davis] | Australia: Queensland | DLL9811 (BRI, CNS) | — | — | MG702641 | [7] |
| E. lepidissimum (Svrček) Noordel. | Czech Republic | PRM 755801, holotype | KC898364 | KC898532 | — | [39] |
| E. luteolamellatum (Largent & Aime) Blanco-Dios [Trichopilus luteolamellatus Largent & Aime] | Guyana | MCA 1480, holotype | — | MH190213 | MG702644 | [7] |
| E. luteum Peck [Inocephalus luteus (Peck) T.J. Baroni] | USA: New York | CORT: 7771 TJB | — | MH190212 | MH190161 | [7] |
| E. mucronatum (Karstedt & Capelari) Blanco-Dios [Inocephalus mucronatus Karstedt & Capelari] | Brazil: São Paulo | SP: FK1033, holotype | — | MH190174 | MH190141 | [7] |
| E. murrayi (Berk. & M.A. Curtis) Sacc. & P. Syd. | China | MHHNU 30602 | MK250917 | — | — | [53] |
| E. murrayi | USA | CORT: TJB 8210 | — | MH190193 | — | [7] |
| E. nitidum Quél. | Slovakia | MEN 200324 | KC710122 | GQ289175 | — | [5] |
| E. pallidoflavum(Henn. & E. Nyman) E. Horak | Vietnam | LE 262934 | OQ779469 | MH190183 | MH190155 | [7], this work |
| E. panniculus (Berk.) Sacc. | Australia | G1680 | — | MK278012 | — | [54] |
| E. percoelestinum O.V. Morozova, Noordel., Vila & Bulyonk. | Spain | LE 254390, holotype | KF745927 | KF745928 | — | [39] |
| E. peristerinum O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343653, holotype | OQ779466 | OQ804522 | OQ779188 | this work |
| E. peristerinum | Vietnam | LE F-343649 | OQ779468 | — | — | this work |
| E. peristerinum | Vietnam | LE F-343650 | OQ779467 | OQ804524 | OQ779186 | this work |
| E. peristerinum | Vietnam | LE F-343652 | — | OQ804525 | OQ779187 | this work |
| E. peristerinum | Vietnam | LE F-343651 | — | OQ804523 | OQ779189 | this work |
| E. placidum (Fr.) Noordel. | Sweden | UPS:BOT:F-121714, epitype, | KC898394 | KC898514 | — | [39] |
| E. plicatum (Largent) Blanco-Dios | USA | DAR: DLL10091 | — | — | MG702627 | [7] |
| E. prunuloides (Fr.) Quél. | USA | AFTOL-ID: 523 4765 TJB | — | — | DQ457633 | [55] |
| E. quadratum (Berk. & M.A. Curtis) E. Horak [Inocephalus quadratus (Berk. & M.A. Curtis) T.J. Baroni] | Russia: Far East | LE254355 | KC898452 | KC898504 | — | [39] |
| E. quadratum | USA: New York | CORT: TJB 8214 | — | — | MH190162 | [7] |
| E. sericellum (Fr.) P. Kumm. [Alboleptonia sericella (Fr.) Largent & R.G. Benedict] | Russia: Caucasus | LE 254362 | KC898453 | — | — | [39] |
| E. sericellum | USA: California | HSU: DLL9524 | — | — | MG702617 | [7] |
| E. sericellum | Belgium | MEN 200315 | — | GQ289190 | — | [5] |
| E. serrulatum (Fr.) Hesler | Russia: Caucasus | LE 254361 | KC898447 | KC898501 | — | [39] |
| E. sublaevisporum Vila, Noordel. & O.V. Morozova | Spain | LIP JVG 1070823T, holotype | KC898436 | KC898518 | — | [39] |
| E. tadungense O.V. Morozova et T.H.G. Pham | Vietnam | LE F-343680, holotype | OQ779470 | — | — | this work |
| E. tadungense | Vietnam | LE F-343681 | OQ779471 | OQ804526 | — | this work |
| E. tibiiforme (Largent & Aime) Blanco-Dios [Trichopilus tibiiformis Largent & Aime] | Guyana | MCA2426, holotypus (BRG, LSUM) | — | — | MG702645 | [7] |
| E. tjallingiorum Noordel. var. tjallingiorum | Sweden | UPS:BOT:F-016378, holotype | KC898412 | KC898509 | — | [39] |
| E. turbidum (Fr.) Quél. | Slovakia | MEN 200351 | KC710060 | GQ289201 | — | [5] |
| E. umbrophilum Noordel. & Hauskn. [Leptonia umbrophila (Noordel. & Hauskn.) Largent] | Australia: Queensland | DLL9766 (BRI, CNS) | — | — | MG702638 | [7] |
| E. undatum (Gillet) M.M. Moser | Russia: European part | LE 312417 | MF476910 | MF487801 | — | [42] |
| E. venustum Wölfel & F. Hampe | Germany | L, Wö E17/10, holotype | KC898355 | KC898523 | — | [39] |
| E. vernum S. Lundell | Russia: European part | LE 312538 | OL338282 | OL338537 | OL405539 | [38] |
| E. violaceozonatum Noordel. & Liiv | Estonia | L 275, holotype | KC898448 | KC898502 | — | [39] |
| E. virescens (Sacc.) E. Horak ex Courtec. [Inocephalus virescens (Berk. & M.A. Curtis) Largent & Abell-Davis] | MEL:2379813 | MF977981 | — | — | Genbank | |
| E. virescens | Guyana | MCA 2479 | — | GU384622 | MG702629 | [7] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morozova, O.; Pham, T.H.G. New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam. J. Fungi 2023, 9, 621. https://doi.org/10.3390/jof9060621
Morozova O, Pham THG. New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam. Journal of Fungi. 2023; 9(6):621. https://doi.org/10.3390/jof9060621
Chicago/Turabian StyleMorozova, Olga, and Thi Ha Giang Pham. 2023. "New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam" Journal of Fungi 9, no. 6: 621. https://doi.org/10.3390/jof9060621
APA StyleMorozova, O., & Pham, T. H. G. (2023). New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam. Journal of Fungi, 9(6), 621. https://doi.org/10.3390/jof9060621