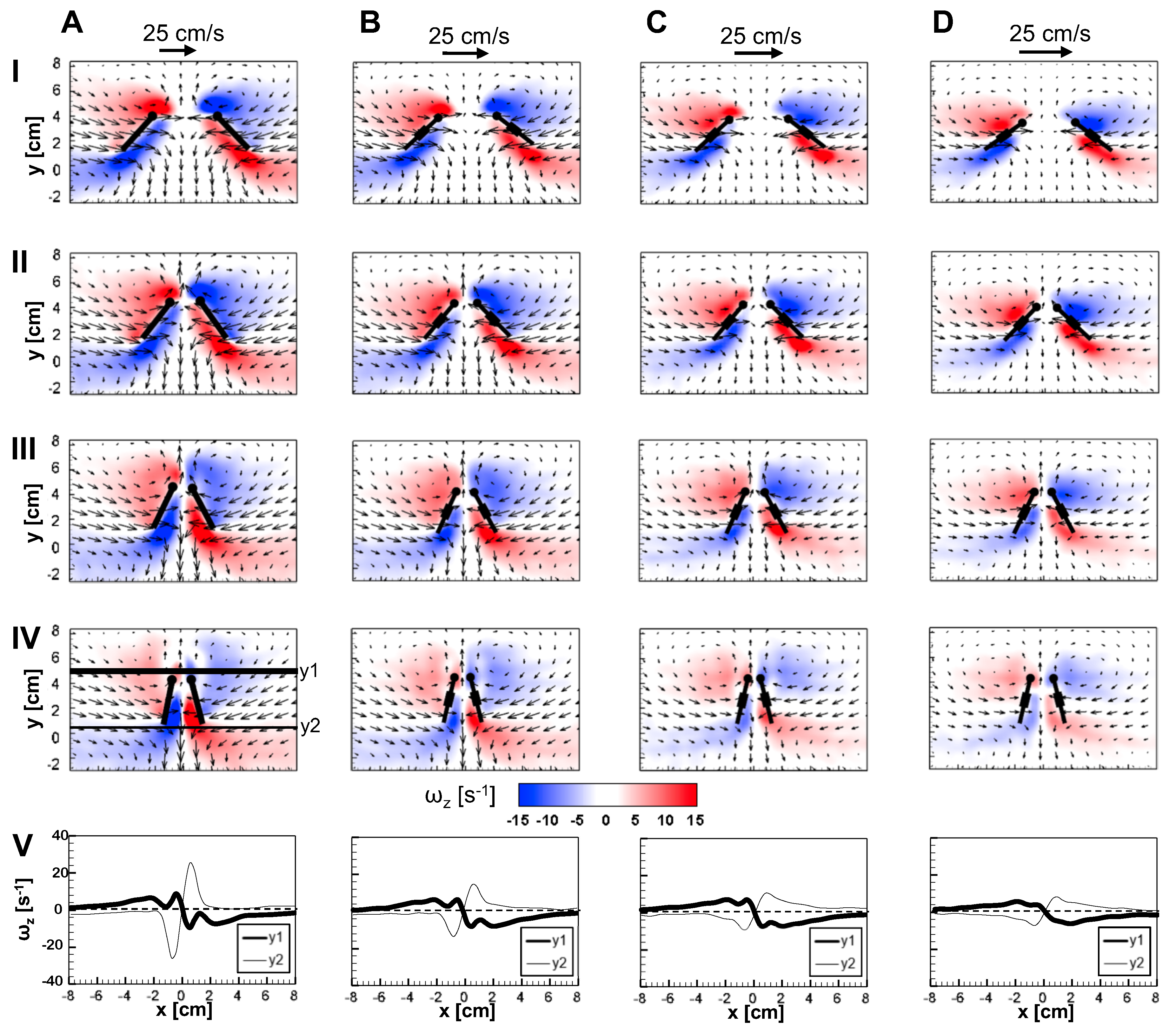
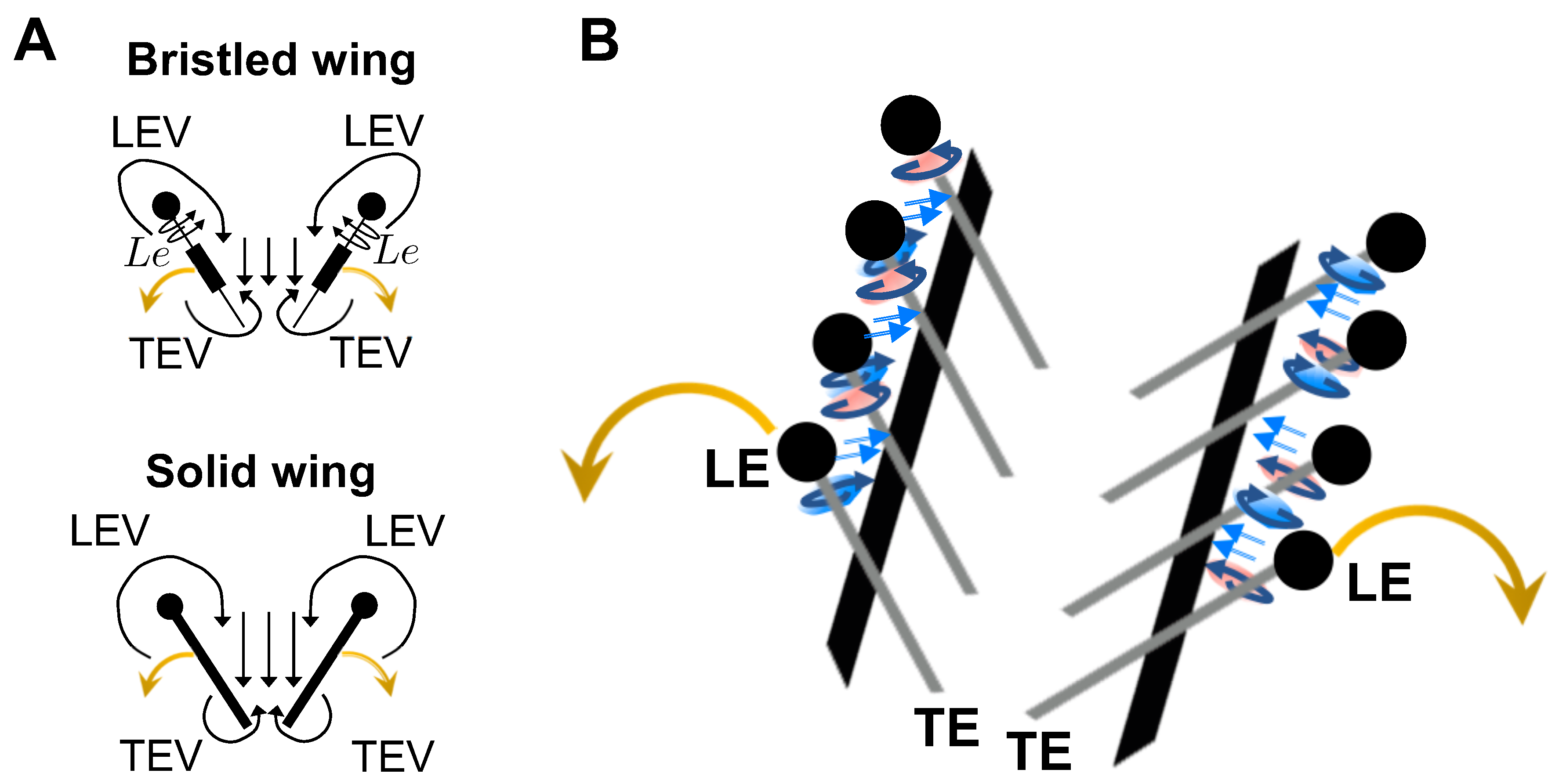
3.1. Chordwise Flow in Clap and Fling: LEV and TEV
Figure 9 and
Figure 10 show 2D velocity vector fields (
) overlaid on out-of-plane vorticity (
) contours at
= 10 for solid and bristled wing models at 4 time points in clap and fling. During clap half-stroke, as both the wings rotated about the leading edge and translated to come together, we observed the formation of both LEV and TEV. For all bristled wing models, we observed that the LEV and TEV were diffused throughout the wing chord (
Figure 9B–D; row V of
Figure 9). This was in direct contrast to the solid wing, where the LEV and TEV were observed to be concentrated near the leading and trailing edges. With increasing clap angle from 12.5% to 62.5%, both the LEV and TEV were observed to grow in both the solid and bristled wing models. With further increase in clap angle (from 62.5% to 87.5%), the strengths of both the LEV and TEV were observed to decrease. A downward jet was observed from the cavity between the trailing edges.
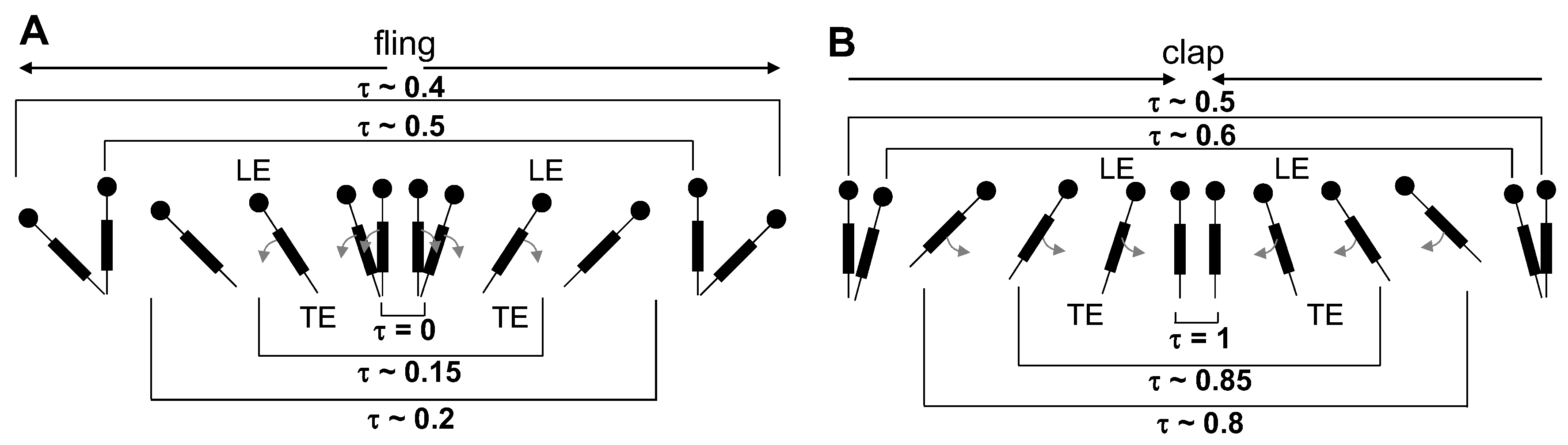
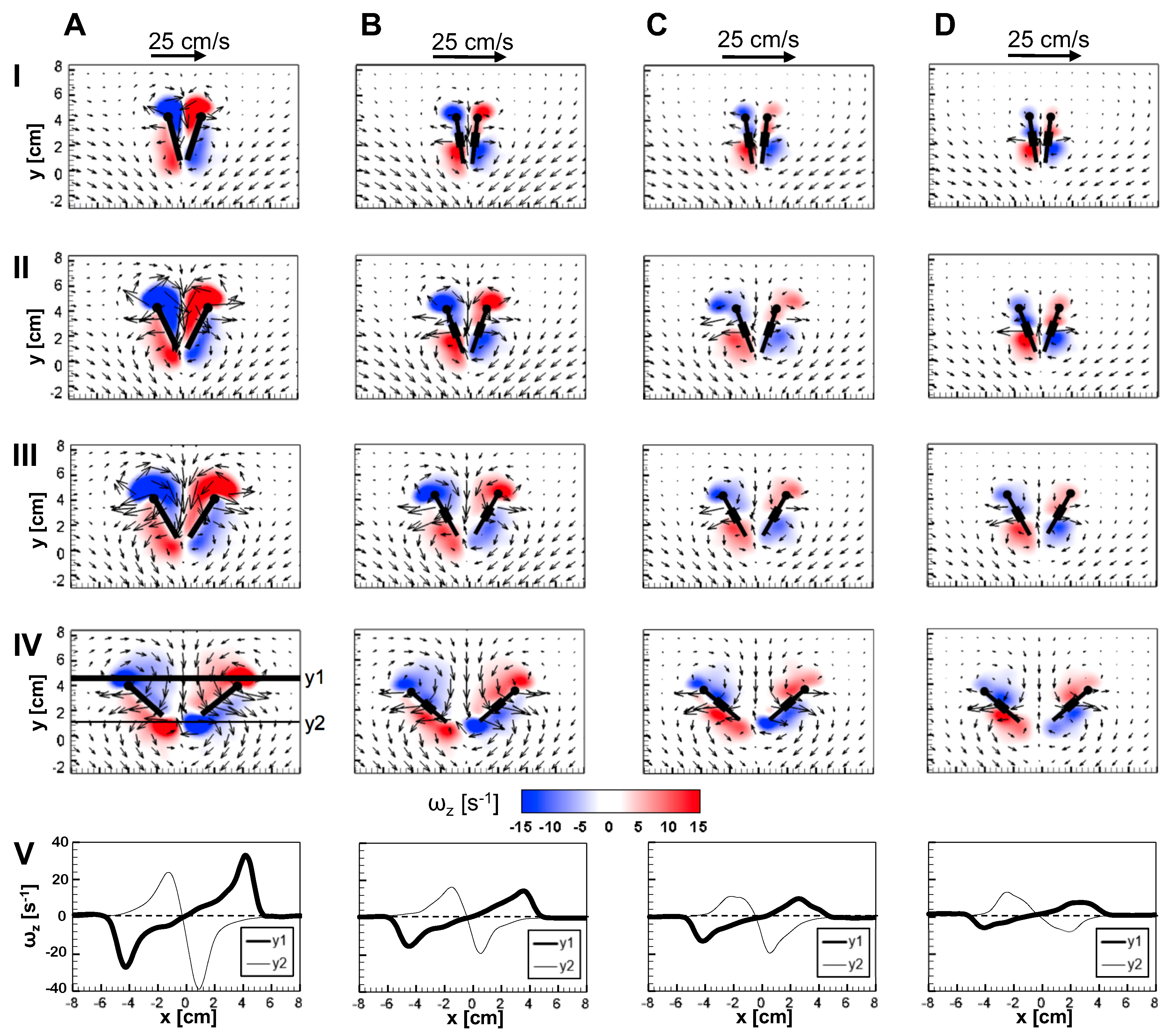
The flow generated by wing rotation during fling using the robotic model developed in this study was qualitatively similar to the observations reported by Lighthill [
27] and Maxworthy [
24]. During fling, as the wings rotated about their trailing edges, two large LEVs formed over each wing at 25% fling angle (
Figure 10). Weak circulating flow with diffuse vorticity was seen at the trailing edges. With increasing fling angle from 50% to 75%, the LEV grew stronger as compared to the TEV in both the solid and bristled wing models. Suction of fluid into the cavity formed between the wings in fling was observed. The onset of wing translation at 50% fling time (
= 1.1 in
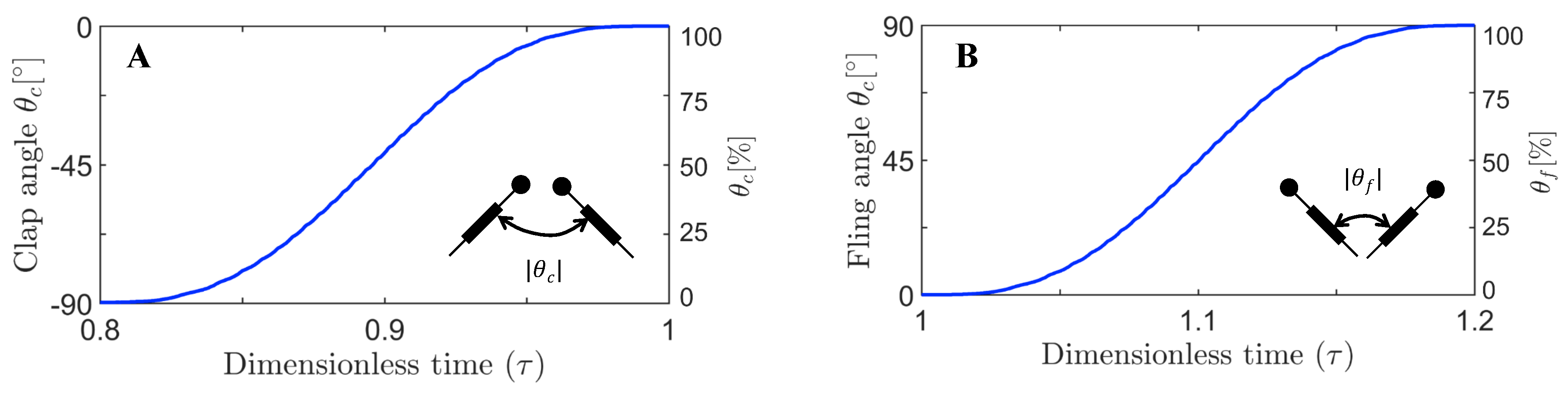
Figure 3), equivalent to roughly ∼45% fling angle (
Figure 5B), did not noticeably enhance vorticity near the TE of solid wing until at 100% angle (
Figure 10A). However, the translation motion was found to make the flow through the chord more diffuse for bristled wing models (
Figure 10B–D).
The velocities generated by the bristled wing pair were markedly smaller when compared to the solid wing pair, showing generation of weaker chordwise flow by bristled wings. Throughout clap and fling, both the LEV and TEV were weaker and highly diffuse for the bristled wing when compared to those of the solid wing (row V in
Figure 9 and
Figure 10). Within the bristled wing models, both the LEV and TEV strengths (proportional to
) were found to decrease with increasing spacing (W/S) between the bristles. Unlike solid wings, vorticity near the TE of the bristled wing model with largest inter-bristle spacing (W/S = 0.23) was close in magnitude to LE vorticity (see row V of
Figure 9 and
Figure 10). Symmetric LEV-TEV has been previously noted to not be conducive for lift generation at low
[
9].
3.2. Force Generation during Clap and Fling
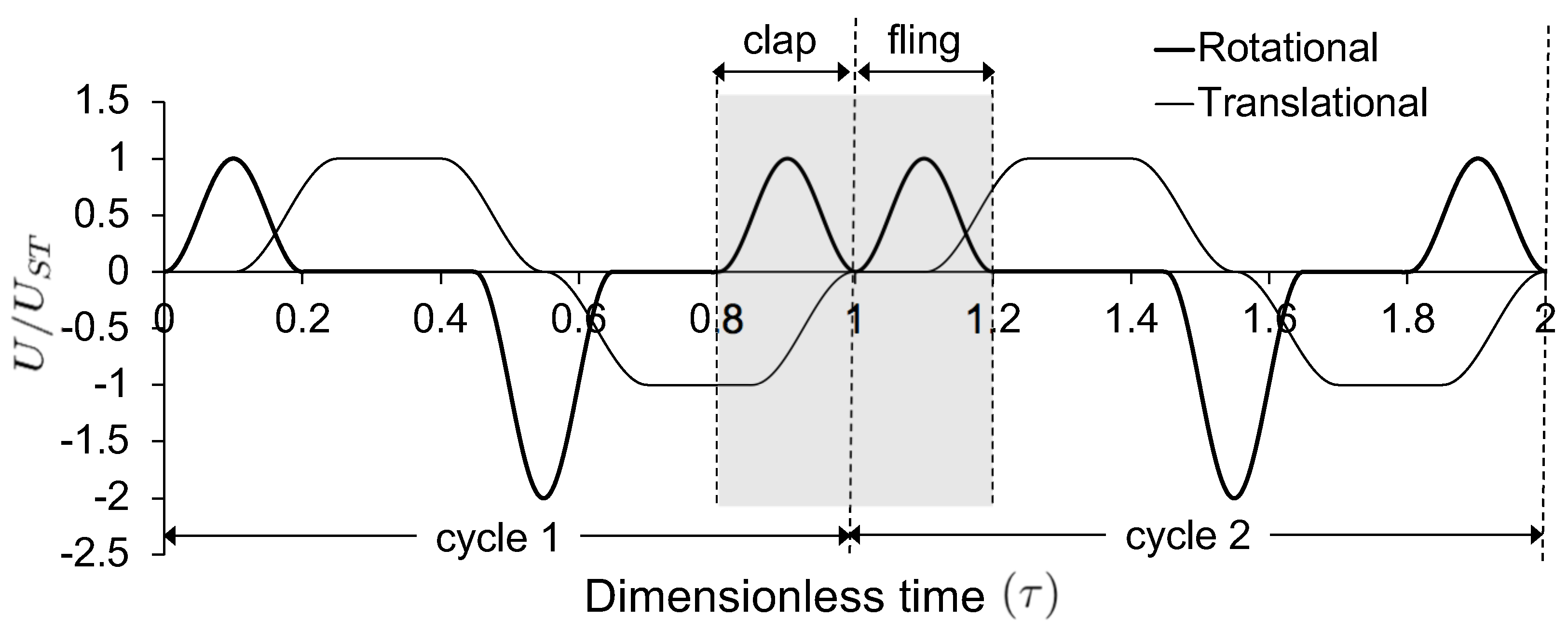
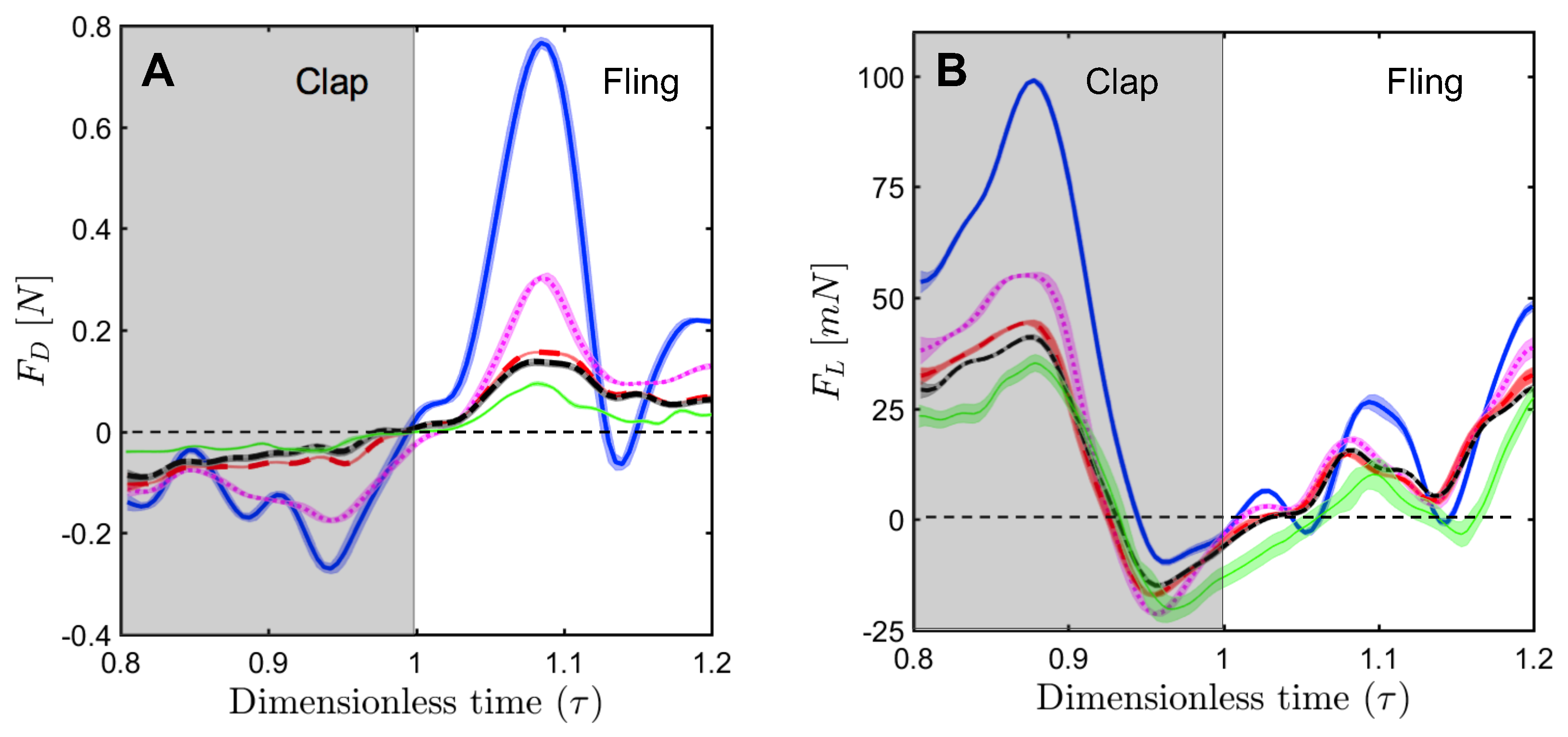
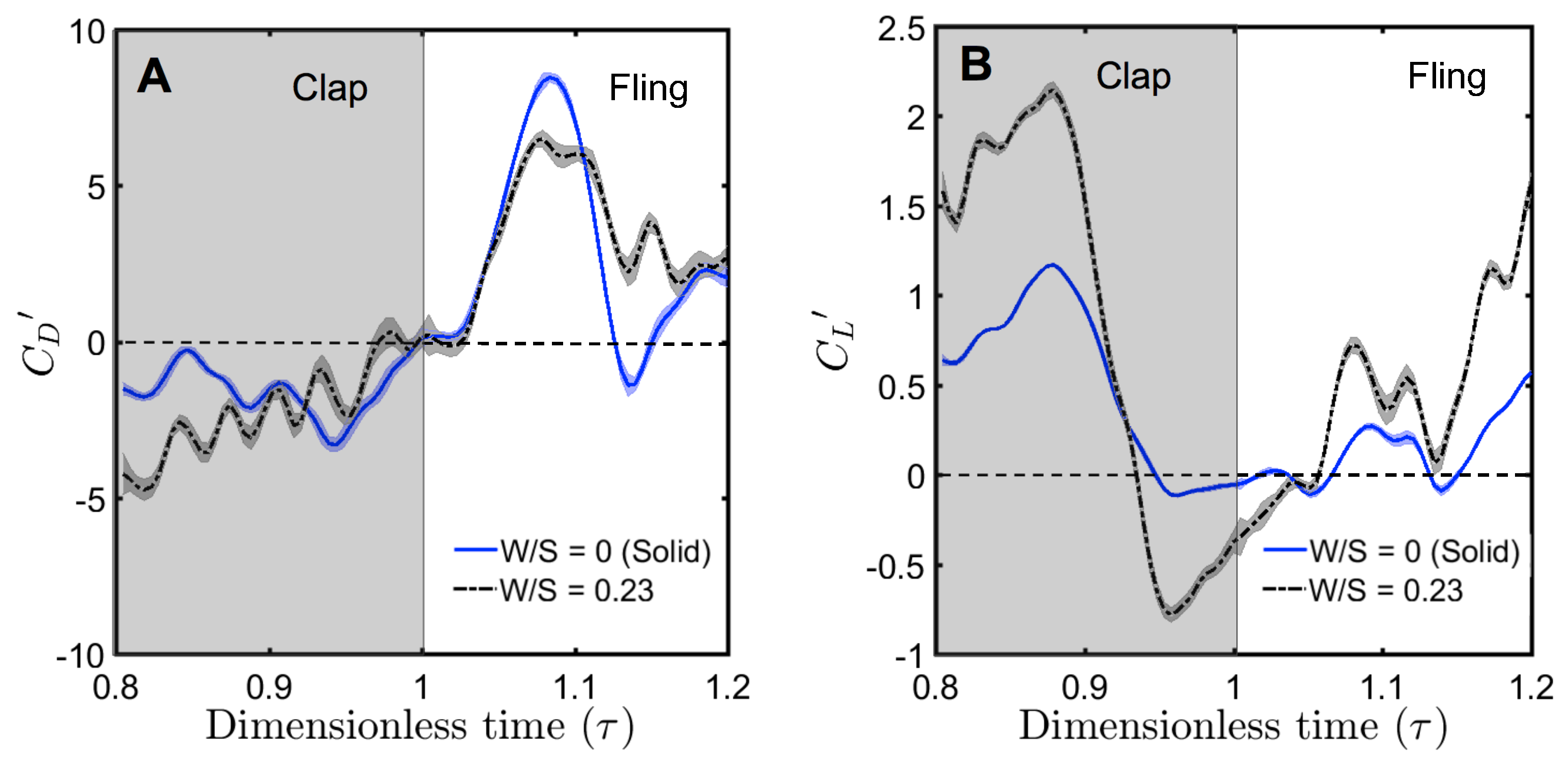
For a solid wing model at
= 10, the drag coefficient reached maximum value during fling half-stroke (
Figure 11A) at around
≈ 1.08 (40% angle in fling half-stroke, see
Figure 5B). This is prior to the start of overlapping translational motion of the wings at
= 1.1 (50% angle in fling half-stroke;
Figure 5B), as seen in the motion profile prescribed to drive wing motion (
Figure 3). This increase in the drag force could potentially be due to added mass effects at the start of fling half-stroke [
36]. In contrast to the solid wing, lower fluctuations were observed in the time-variation of drag coefficients of all bristled wing models (
Figure 11A). This could be due to flow passing through the bristles instead of over the wing. However, for the bristled wing with least spacing (W/S = 0.05), we found the drag force to peak at similar angular position (40% angle in fling half-stroke) to solid wing model, but was smaller in magnitude.
Comparing the drag coefficients between solid and bristled wing models (
Figure 11A) in fling at
= 10, roughly 2–3 times reduction in
was observed across all the bristled wing models. This suggests that bristles can reduce the forces needed to fling the wings, which is supported by findings of previous computational studies [
13,
14]. Another point of interest in
Figure 11A is a small amount of negative drag for solid wing at
≈ 1.14. At this time point, rotational deceleration of the wings occurred simultaneous with the onset of translational acceleration (
Figure 3). Rotational deceleration started prior to this time point (
= 1.1) and decreased drag force as well as TEV strength (proportional to TEV vorticity) across all wing models (
Figure 10). However, the sudden onset of translational acceleration at
= 1.1 was nearly coincident with negative drag force production in the solid wing. This suggests that added mass effects expected with this sudden onset of translational motion could be responsible for the negative drag. Interestingly, no negative drag force was observed at this time point for any of the bristled wing models. With further progress in fling half-stroke (
= 1.14–1.2), translational acceleration increased the drag coefficients of all models. This is supported by TR-PIV visualization of chordwise flow (
Figure 10), where TEV increased in vorticity at the end of fling due to translational acceleration.
Negative drag force during clap indicates that the force was acting opposite in direction to that of fling. During clap, drag force peaked at about
= 0.94 (
Figure 11A). This was prior to the wings coming close together, where the fluid between the wings is expected to resist the wings from coming in close proximity. During both clap and fling, peak drag coefficients were found to decrease with increasing inter-bristle spacing (W/S).
In contrast to the drag reduction seen in the bristled wings, lift generated by solid and bristled wings during clap and fling at
= 10 was roughly in the same order of magnitude (
Figure 11B). The lift force generated was found to reach peak value during clap at around
= 0.88 (40% angle in clap half-stroke;
Figure 5A) for all the wing models. This could be potentially due to wing–wake interaction during stroke reversal (90 degrees rotation before start of clap half-stroke ;
≈ 0.4–0.7 in
Figure 3 and
Figure 4). This is also prior to start of decelerating portion of translational motion at
= 0.9 (50% angle in clap half-stroke in
Figure 5A). During clap, the lift coefficient was observed to decrease from solid to bristled wing model of least spacing (W/S = 0.05) by a small amount. A sudden drop in lift was observed at the end of the clap (
Figure 11B) due to decreased vorticity at the leading edge (
Figure 9). In addition, a small amount of negative lift was observed at the end of clap. This could be due to deceleration of wings and flow induced by the other wing, as both wings come close together. During most of the fling half-stroke, lift coefficient was found to only change slightly from solid to any bristled wing model. A small amount of negative lift was observed during fling for the solid wing at
= 10 (
Figure 11B). Little to no negative lift was observed in all bristled wing models during fling at
= 10, showing a modest improvement from the solid wing purely via the inclusion of bristles. Peak value of lift coefficient at
= 10 was slightly lower for the bristled wing as compared to the solid wing (
Figure 11B). Increasing spacing between the bristles (W/S) did not noticeably affect the lift coefficients, as compared to the impact of changing W/S on the drag coefficients (
Figure 11A).
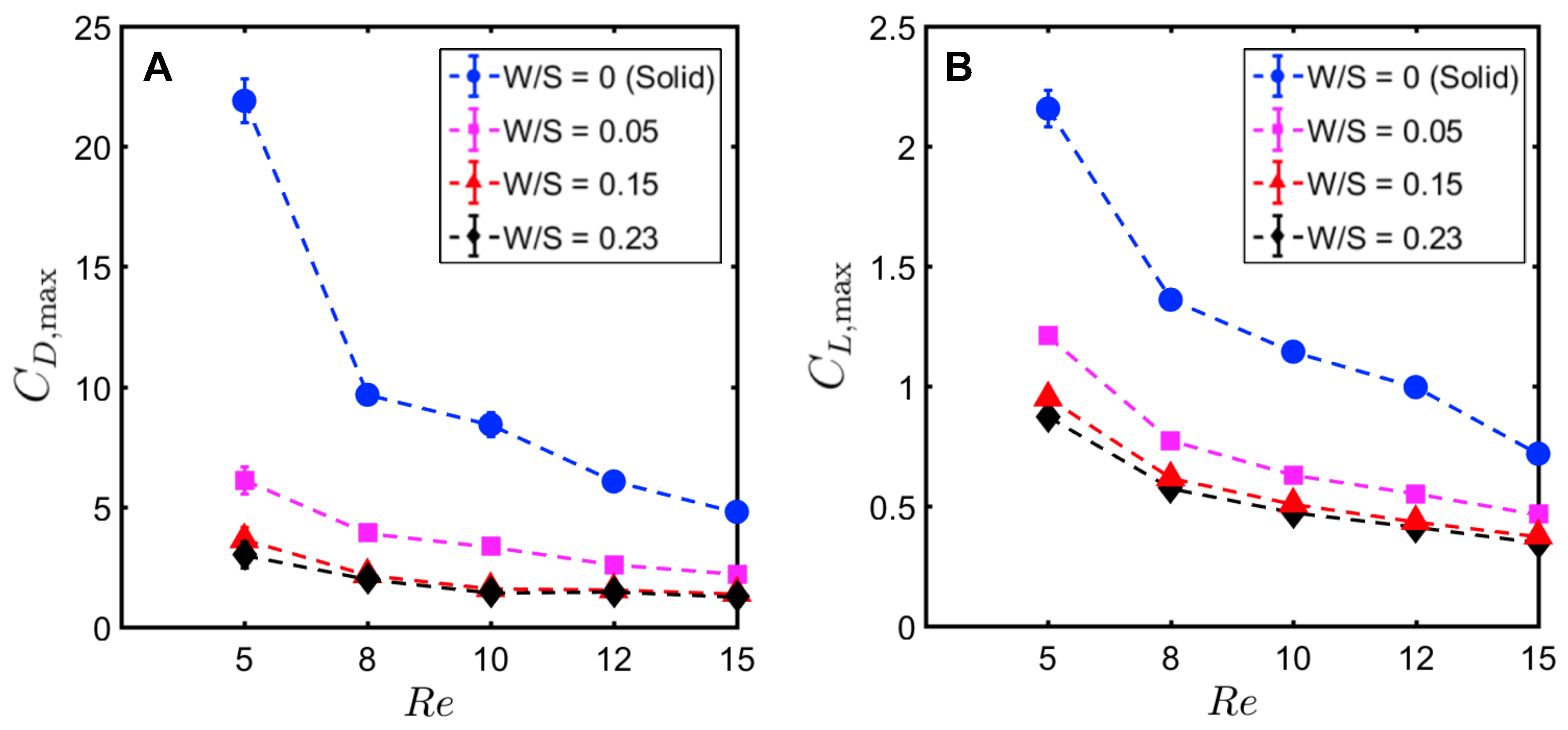
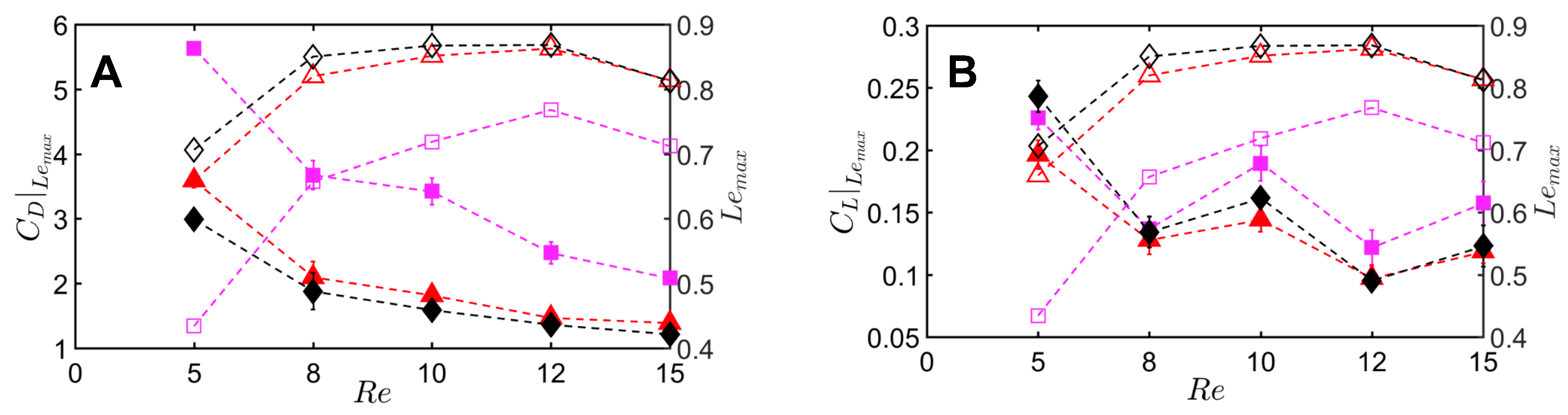
Peak drag (
) and lift coefficients (
) were calculated from the absolute values of drag (
) and lift coefficients (
) throughout the clap and fling cycle. Peak drag and lift coefficients during clap and fling of the solid wing model were nearly doubled when decreasing
from 10 to 5 (
Figure 12A,B). This shows the inherent challenge with generating sufficient lift relative to drag at
∼
(1), corresponding to the lowermost limit of flapping flight by insects [
13,
14,
28]. Within
range of 5–10, all bristled wing models (W/S = 0.05, 0.15, 0.23) showed an order of magnitude reduction in peak drag coefficient when compared to the solid wing. Looking at only bristled wing models in the
range of 5–10, large reduction in drag was found with increasing W/S from 0.05 to 0.15. With further increase in spacing between the bristles (W/S = 0.15 to W/S = 0.23), the peak drag coefficients did not noticeably change. A similar trend was observed for peak lift coefficient of bristled wings in
range of 5–10. Increasing
from 10 to 15 did not show a marked impact in peak force coefficients of solid and bristled wings. However, the reduction in peak lift was smaller compared to reduction in peak drag (
Figure 12A,B) when comparing solid and bristled wings. Both peak force coefficients decreased with increasing
from 5 to 15 irrespective of W/S (
Figure 12A,B). Decrease in peak force coefficients with increasing
was most pronounced for the solid wing (
Figure 12A,B). Overall, the inclusion of bristles rendered the peak force coefficients to only slightly vary with
in the range of 5–15. The most pronounced impact of the bristles in terms of large drag reduction (over solid wings) was observed in
range of 5–10, corresponding to the biological
range of flapping flight in tiny insects that posess bristled wings [
14].
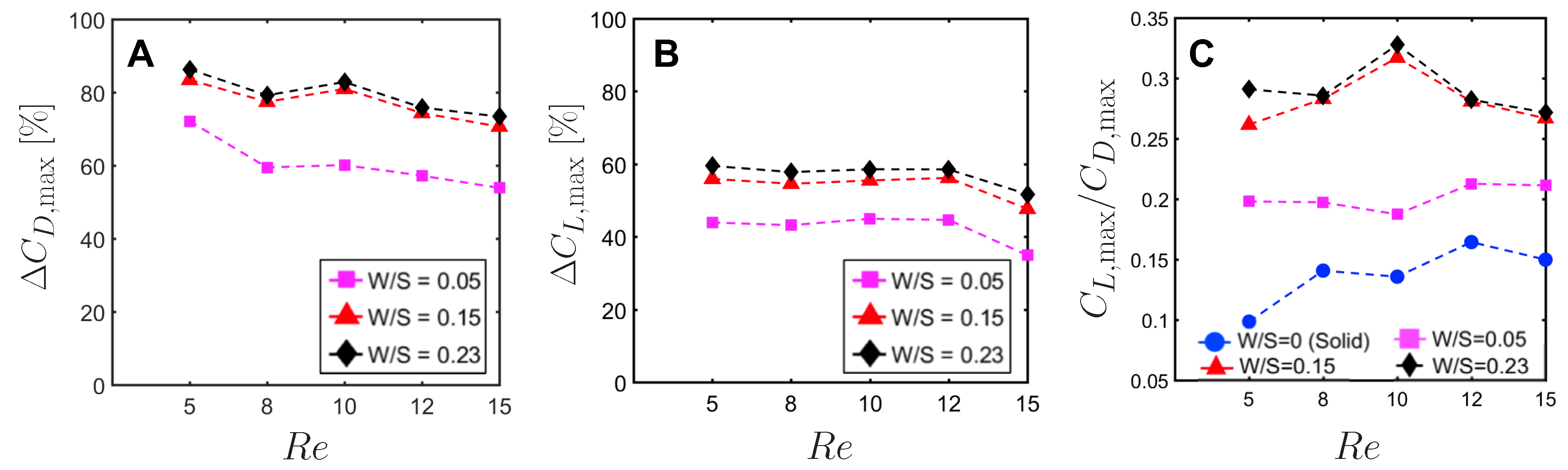
To quantify the reduction in peak drag and lift coefficients between a bristled wing pair (non-zero W/S) to the solid wing pair (W/S = 0), we calculated
and
using Equation (
10). Across the range of
tested, bristled wings reduced peak drag coefficient by 50%–90% and peak lift coefficient by 35%–60% when compared to the corresponding peak coefficients of the solid wing model (
Figure 13A,B). Thus, the inclusion of bristles can disproportionally affect lift and drag generated during clap and fling—even for the smallest W/S model examined here (W/S = 0.05). Across
ranging from 5 to 15, peak lift to peak drag ratios (
Figure 13C) were larger in all the bristled wing models when compared to the solid wing model. While peak lift to peak drag ratio increased with increasing non-dimensional inter-bristle spacing (W/S) from 0.05 to 0.15, the ratio was nearly unaffected when W/S was further increased to 0.23. These results demonstrate that bristled wings can provide aerodynamic benefits over solid wings for
ranging between 5 and 15 via: (a) augmentation in peak lift to peak drag ratio; and (b) disproportionally larger drag reduction than lift reduction.
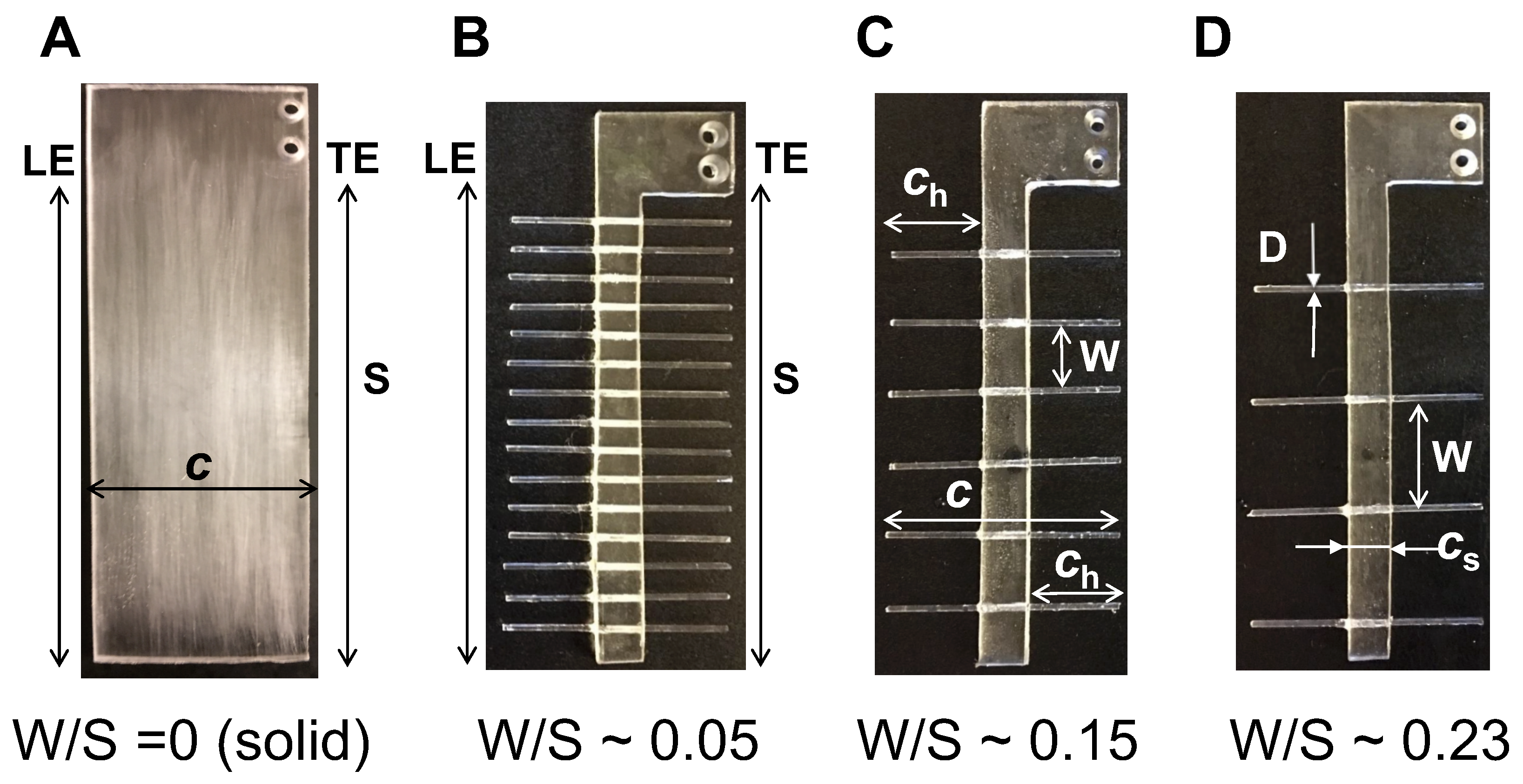
From the above results, we observe that ∼55% increase in gap width (W) relative to span (S) from 0.15 to 0.23 produces small changes in the peak force coefficients. To examine the relative importance of even a sparse number of bristles (
n = 8 in W/S = 0.23 model) on aerodynamic force generation, we acquired force measurements on a membrane-only model with no bristles at
= 10. The membrane-only model can be viewed as the limiting case of W/S > 1. The time-variation of lift and drag forces of the membrane-only model (
= 8.5 mm and span S = 90 mm as in
Figure 1) was observed to follow a similar trend as the other wing models in this study (
Figure A3). For the membrane-only model, peak drag was observed at
≈ 1.08 (40% angle in fling half-stroke;
Figure 5B) and peak lift occurred at
≈ 0.88 (40% angle in clap half-stroke;
Figure 5A)—both similar to the other wing models (
Figure A3). However, the magnitudes of both lift and drag forces were lower for the membrane-only model when compared to the sparsest bristled wing model (W/S = 0.23). In addition, the peak lift to peak drag ratio was roughly 53% lower in the membrane-only model (
= 0.16) when compared to the sparsest bristled wing of W/S = 0.23 (
= 0.3). This suggests that having even a small number of bristles for a given span (as in W/S = 0.23), such that W/S < 1, does indeed impact force generation during clap and fling at
∼
(10). More experiments are needed on bristled wing models with 0.23 < W/S < 1 to identify the precise ratio of gap width to span (W/S) and number of bristles (
n) where peak force coefficients are essentially unchanged.
When force coefficients for the bristled wing were calculated using the scaled (or reduced) surface area given by Equation (
11), a reduction in bristled wing drag coefficient (
) was observed as compared to the solid wing (
Figure A4A) during pure wing rotation in fling (
≈ 1–1.1,
Figure 5B). This was similar to what was observed when using identical (or characteristic) surface areas between both wing models (
Figure 11A). The scaled drag coefficient of the bristled wing (
in
Figure A4A) increased when translation and rotation started to overlap (
≈ 1.1) until
≈ 1.12, after which no difference was observed between the bristled and solid wings. However, the scaled lift coefficients of the bristled wing (
in
Figure A4B) were larger than the solid model for most of the fling half-stroke (
1.05 in
Figure A4). This is in exact contrast to the minor reduction in lift coefficient of the bristled wing when using identical surface areas (
Figure 11B). Examining dimensional lift forces for this particular test condition (
Figure A3B) shows that calculation of lift coefficient using reduced surface area can incorrectly suggest that lift augmentation occurred in bristled wings during fling as compared to the solid wing.
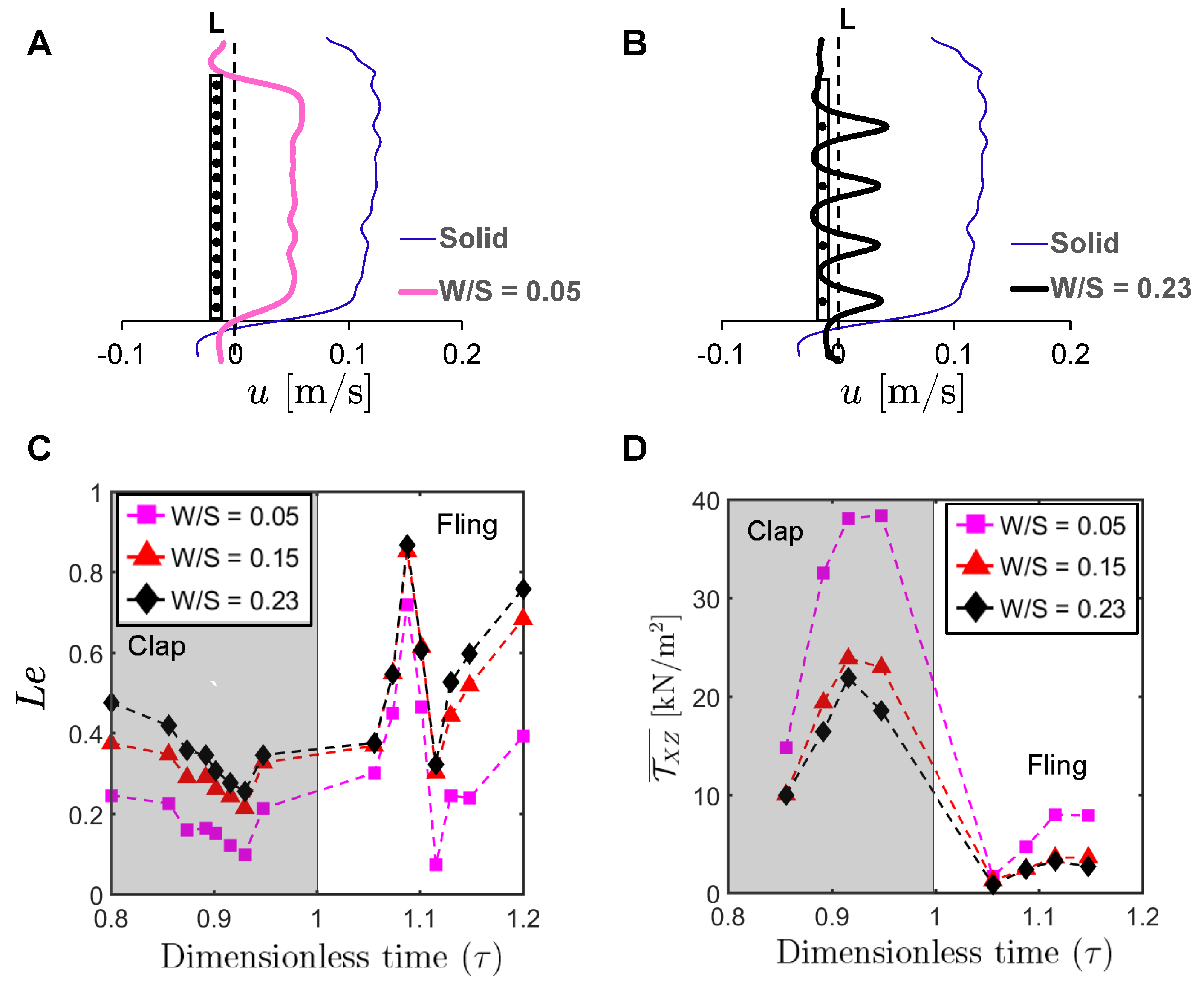
3.3. Inter-Bristle Flow during Clap and Fling: Leakiness
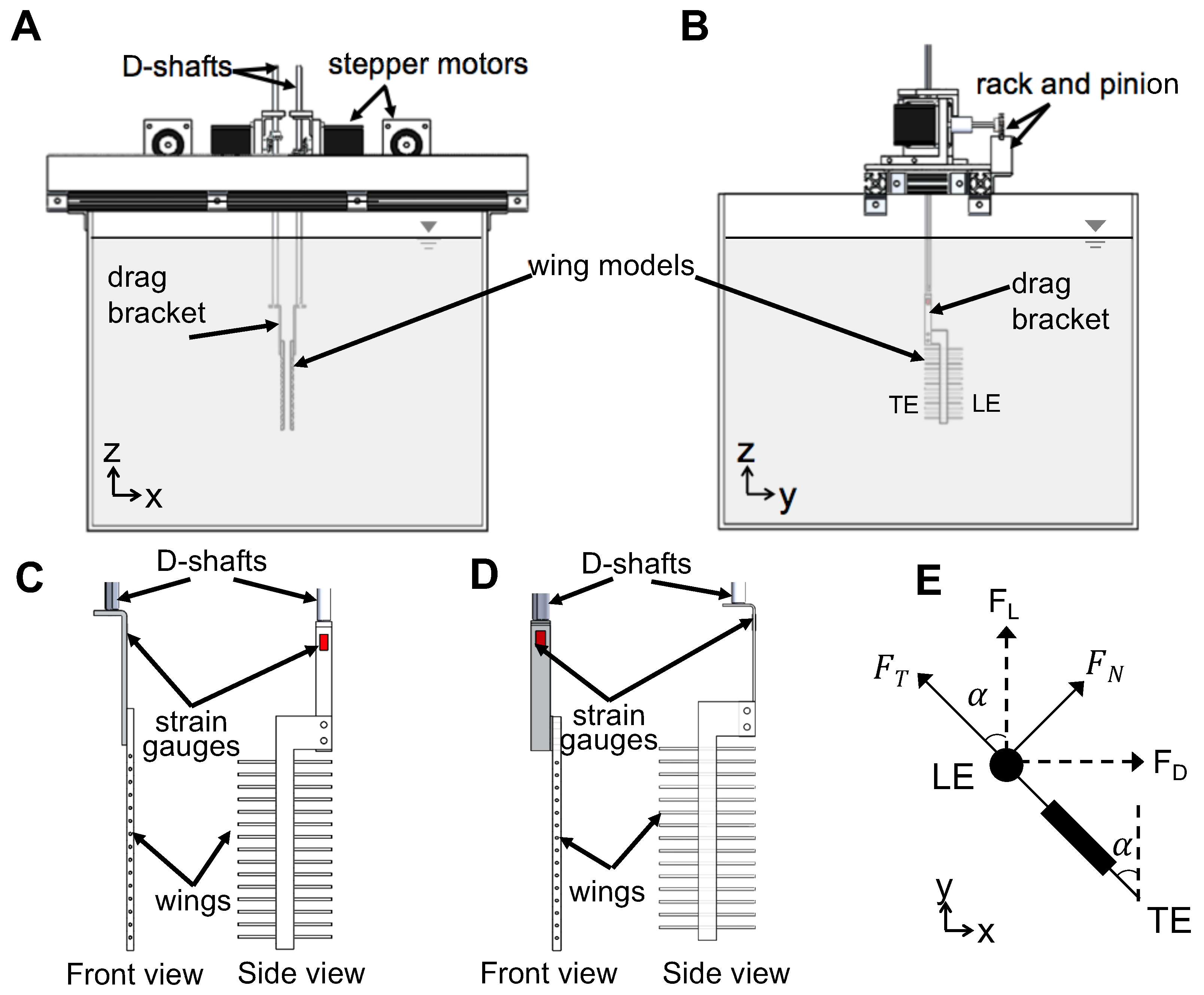
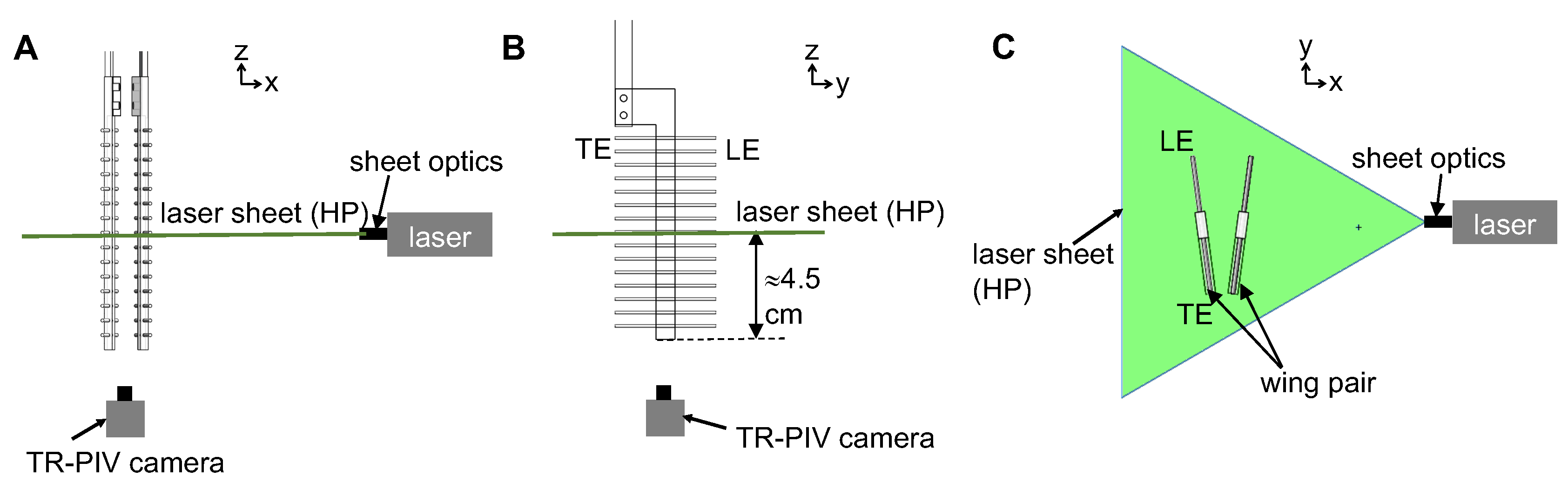
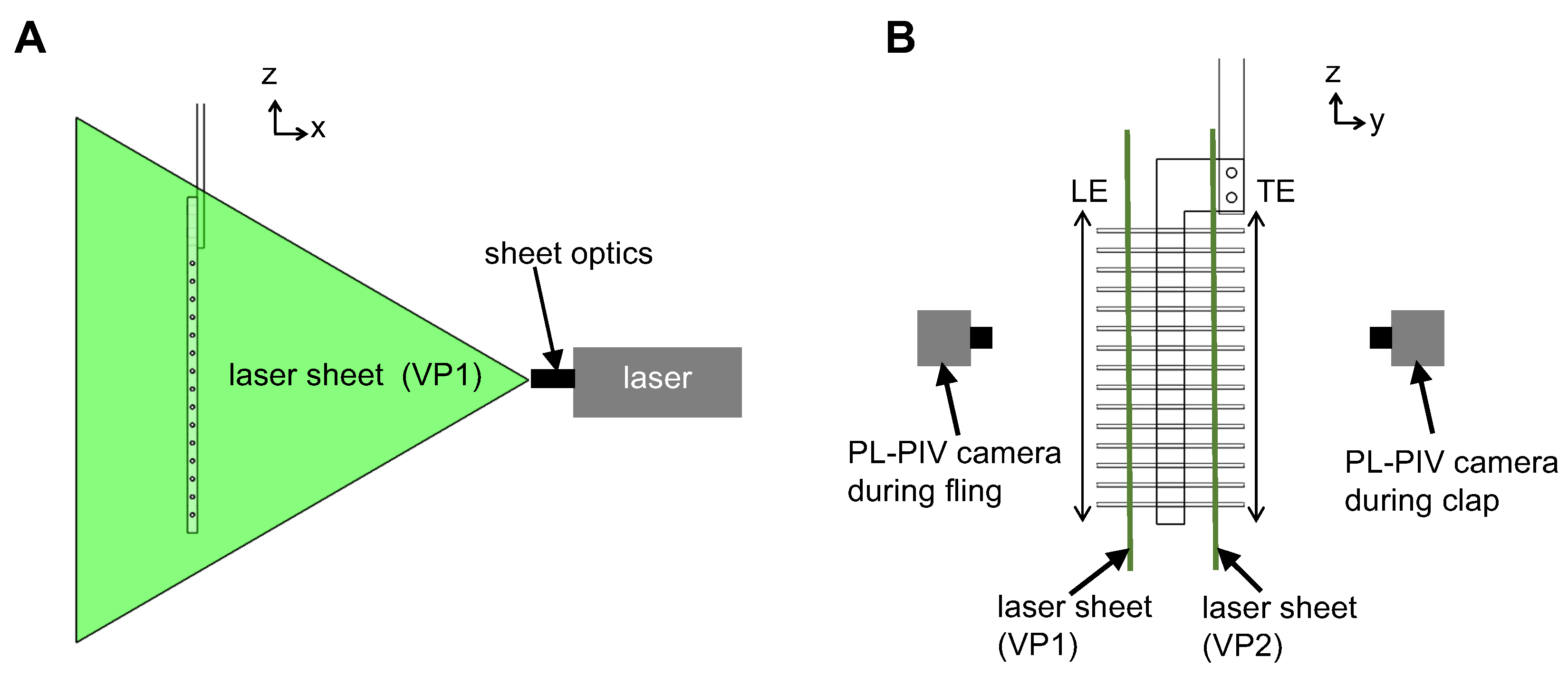
To examine the physical mechanism underlying drag reduction by inclusion of bristles, we examined flow fields in spanwise planes VP1 and VP2 (
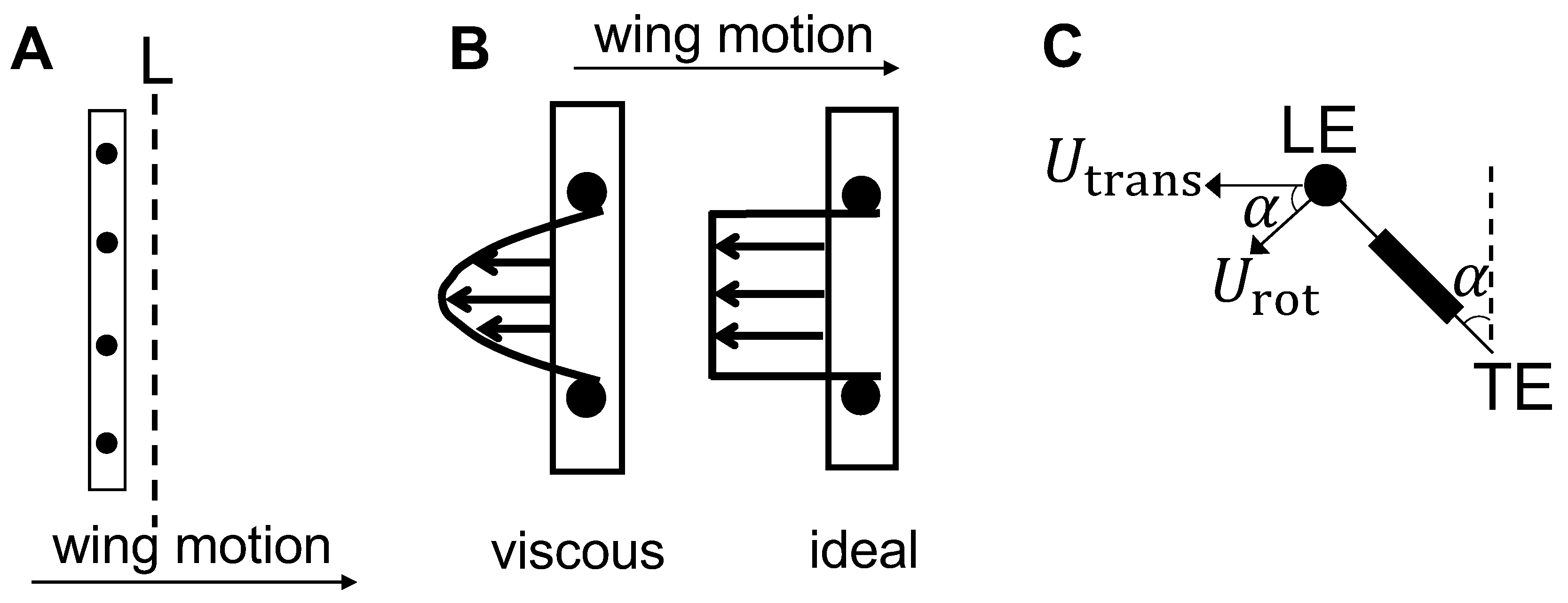
Figure 7B) via PL-PIV. Horizontal velocity profiles were extracted at line “L”, located at a distance of about 10% chord in the same direction as wing motion (
Figure 8A), for
= 37.5% and
= 10 (
Figure 14A,B). Velocities decreased for the bristled wing models with smallest and largest gap widths (W/S = 0.05 and 0.23) when compared to the solid wing, suggesting that flow leaked through the gaps in these bristled wing models. The spanwise distribution of horizontal velocity in the bristled wing model with least spacing (W/S = 0.05) followed the same trend as that of the solid wing (
Figure 14A). In contrast, recirculating flow with both negative and positive velocities were observed in the bristled wing model with the largest spacing (W/S = 0.23;
Figure 14B). Fluid leakage through the gaps between bristles was quantified using leakiness (
) calculated from Equations (
12)–(
16). Irrespective of W/S,
showed similar variation across the clap and fling cycle for
= 10 (
Figure 14C).
varied between 0.1 and 0.5 in clap and between 0.1 and 0.9 in fling. The time point where peak
was observed for
= 10 (
= 37.5% in
Figure 5B;
≈ 1.08) remained invariant with changing spacing between the bristles (W/S).
Despite the ∼55% increase in W/S,
of W/S = 0.15 and W/S = 0.23 models were quite similar at
= 10 (
Figure 14C). It is intuitive to expect that the W/S = 0.23 model should permit a larger volume of fluid to leak through the bristles when compared to the W/S = 0.15 model with a smaller gap width (W). However, this expectation does not necessitate that
of W/S = 0.23 model also be greater than
of W/S = 0.15 model. As per Equation (
12),
is the ratio of flow rate leaked by a bristled wing model under viscous conditions (
) to the theoretical/ideal flow rate that would be leaked by the same wing model in an inviscid flow (
). A small change in
, such as between W/S = 0.15 and 0.23 models at
= 10 (
Figure 14C), can therefore be interpreted as both models leaking fluid by roughly the same fraction relative to their individual ideal volumetric flow rate.
While
was calculated from PL-PIV data,
depends on the bristled wing design and prescribed wing kinematics as per Equation (
13). To maintain the same wing span and bristle diameter across all bristled wing models, W was increased in our models by lowering the number of bristles (
n) based on Equation (
1). The ideal volumetric flow rate, given by Equation (
13), would increase with decreasing
n. As
is identical across all wings for a specific value of
, it will cancel out when considering ratio of
between two bristled wing models of different W/S. Thus, Equation (
13) can be used to give the following ratios for a specific value of Re:
= 84.8/82.2 = 1.03 (3% increase in
). In contrast,
= 82.2/70.5 = 1.17 (17% increase in
); and
= 84.8/70.5 = 1.20 (20% increase in
). As
is inversely related to
as per Equation (
12),
of W/S = 0.15 and 0.23 bristled wing models need not show as much of a difference as compared to the differences in
of: W/S = 0.05 and 0.15 models, or W/S = 0.05 and 0.23 models. When W/S was increased from 0.15 to 0.23, the expected increase in
(due to larger inter-bristle spacing) in W/S = 0.23 was most likely countered by a small increase in
in W/S = 0.23—resulting in only a minimal increase in
as per Equation (
12). In this context, it is important to note that the definition of
used in this study was originally proposed by Cheer and Koehl [
34] to quantify leakage flow through an array of two cylinders. Despite use of the same definition to quantify
through arrays of multiple cylinders (as in this study), the close similarity in
between W/S = 0.15 and W/S = 0.23 wing models was also mirrored by the similarity of peak force coefficients of the same two wing models (
Figure 12).
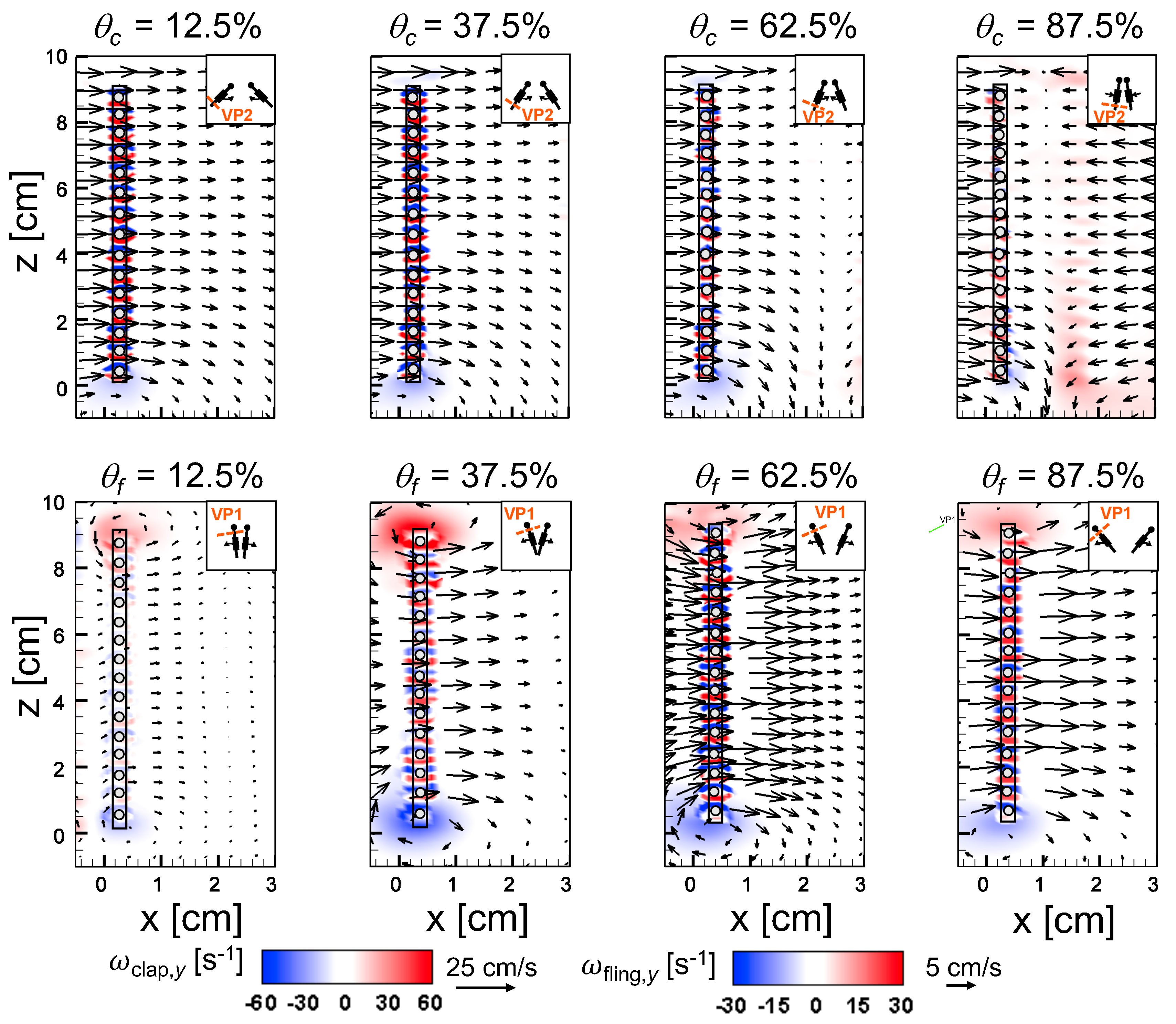
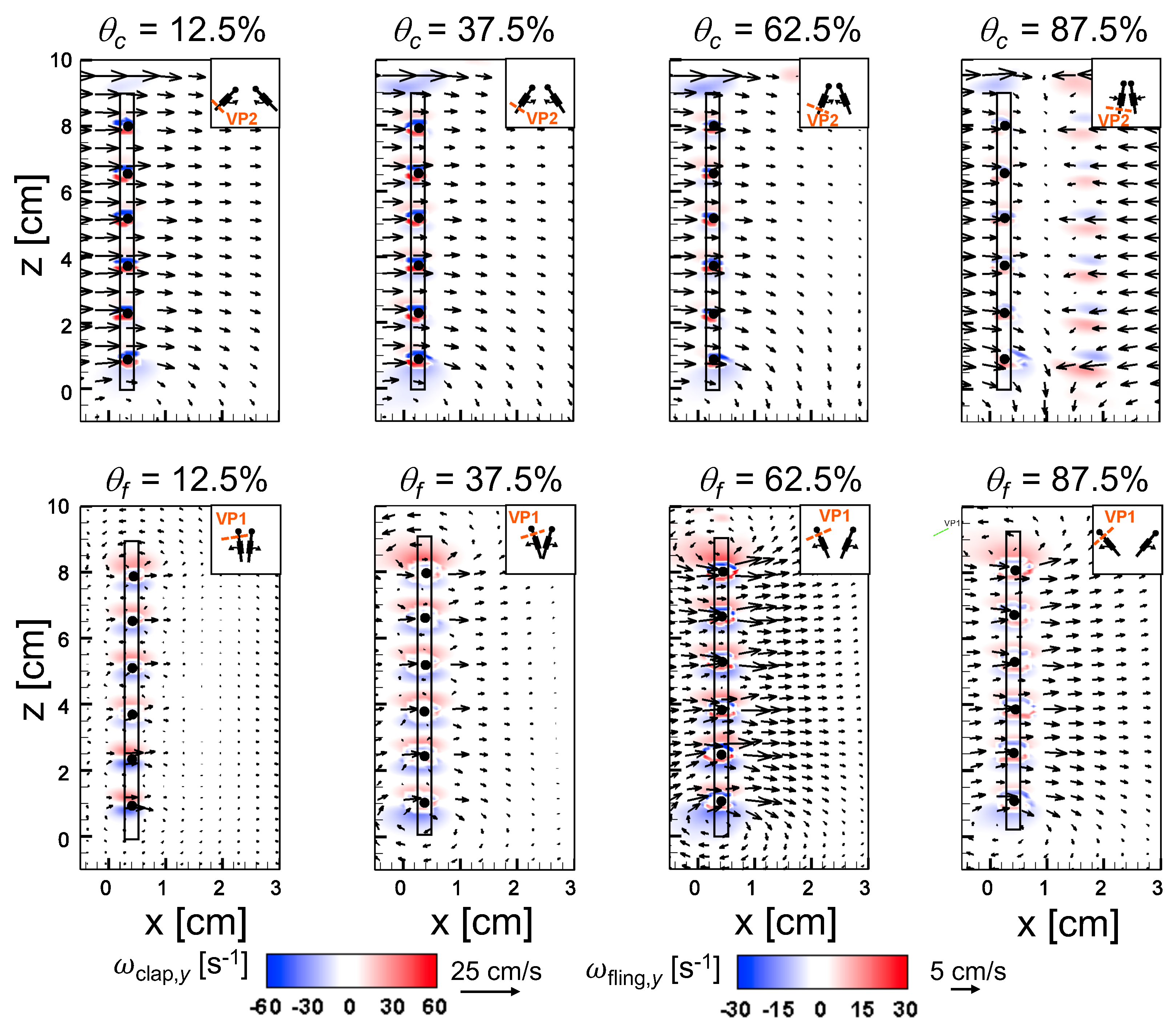
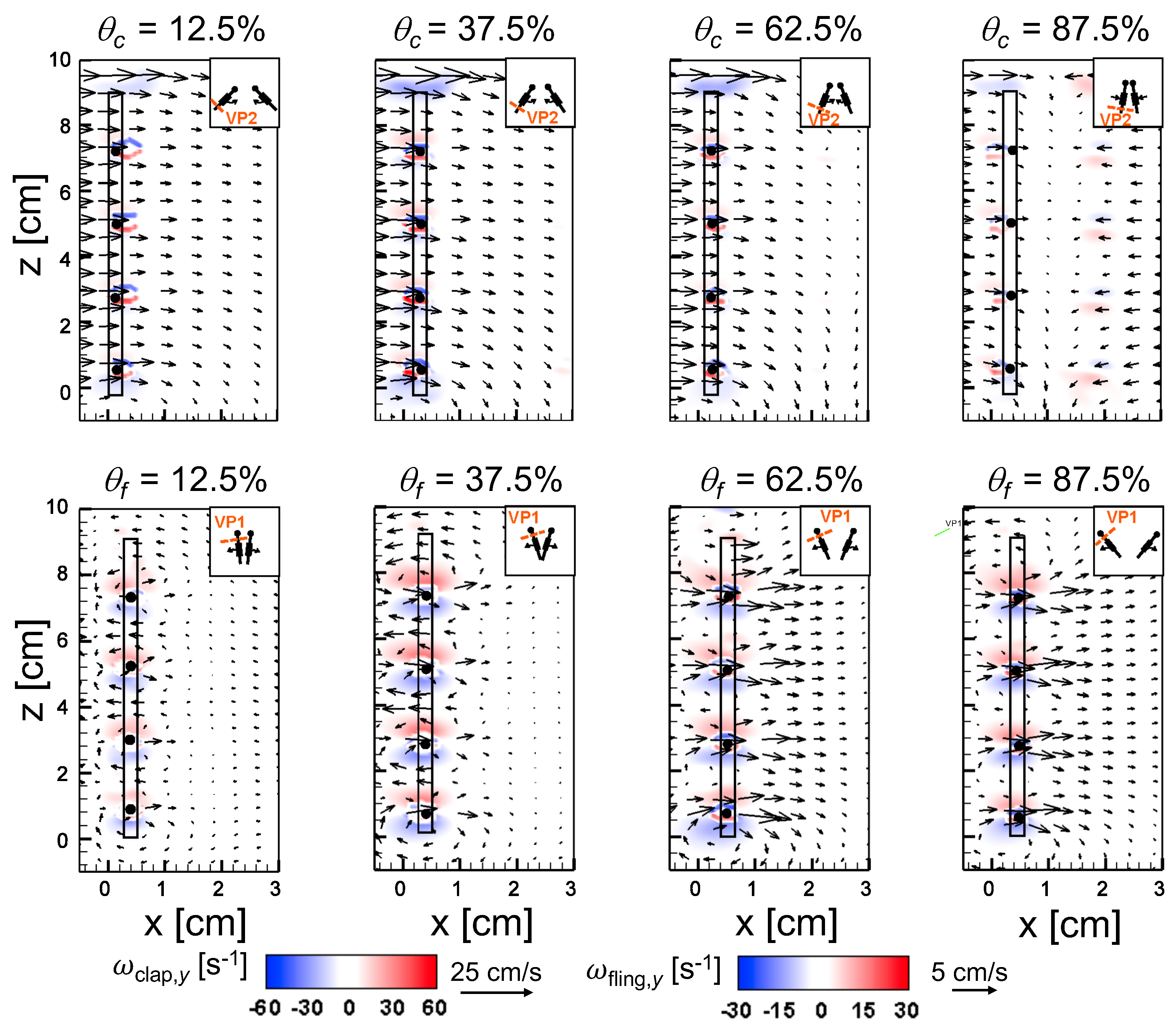
An examination of the inter-bristle vorticity and velocity fields during clap and fling (
Figure 15,
Figure 16 and
Figure 17) shows that high-shear regions are formed around the bristles at
= 10. For all bristled wing models, shear layers around bristles was observed and was of significant value throughout the clap half-stroke. During fling, the shear layers around bristles were found to be minimum for
< 37.5%. The vorticity of these shear layers increased until
= 62.5% and decreased for
> 62.5% of fling half-stroke. This was coincident with the time points where peak leakiness (
= 37.5% fling angle,
≈ 1.08) and minimum leakiness (
= 62.5% fling angle;
≈ 1.12) were observed. This suggests that rotation and translation can impact leakiness in opposite manner, such that pure rotation in fling increases leakiness while onset of translation decreases leakiness and aids in increasing vorticity of shear flow around the bristles. With increase in spacing between the bristles (W/S) from 0.05 to 0.23, the vorticity of shear layers formed around the bristles was found to decrease during both clap and fling for all
and
(
Figure 15,
Figure 16 and
Figure 17).
For bristled wing model of W/S = 0.23, we observed the flow to recirculate around the bristles for
< 62.5% fling angle (
Figure 17 and
Figure A5C), while this phenomenon was absent during clap. In addition, the magnified inter-bristle flow field for W/S = 0.15 at
= 37.5% (
Figure A5B) showed the onset of small amount of recirculation around each bristle. However, the regions of recirculating flow were found to be quite small for W/S = 0.15 due to large shear layers (
Figure A5B) formed around neighboring bristles. These shear layers could further reduce the effective gaps between bristles and thereby limit the extent of recirculation and reverse flow through the bristles. For W/S = 0.05, larger shear layers without any recirculation were observed around the bristles when compared to W/S = 0.15 and 0.23 (
Figure A5A). These factors could potentially contribute to lower leakiness in W/S = 0.05 as compared to other two bristled wing models of larger W/S.A larger gap between adjacent bristles (as in W/S = 0.23) would favor the development of recirculation regions around bristles. However, increasing the gap to such an extent that there is no interaction between shear layers of adjacent bristles may not favor the development of recirculation. Comparing the W/S = 0.23 model with the limiting case of membrane-only wing model with no bristles (
Figure A3), we can observe that interactions between the shear layers formed around adjacent bristles can have a non-negligible impact on forces generated during clap and fling. In the case of W/S = 0.23 model, it appears that the gap width was large enough for more volume of flow to leak through than W/S = 0.05, but not so large so that shear layers of adjacent bristles in W/S = 0.23 did not interact. If the latter were true, we would not expect: (a) recirculation of flow around bristles in W/S = 0.23 model; and (b) there to be any appreciable variation in forces generated by W/S = 0.23 and membrane-only models (contradicted by
Figure A3). Although W/S = 0.05 favors the formation of high-shear regions around the bristles on account of the small gap width, there is not adequate gap in between interacting shear layers to permit flow to recirculate.
To quantify the strength of the shear layers formed around the bristles, we calculated average shear stress
using Equation (
17) at the same location “L” where
was determined (
Figure 8A). Irrespective of W/S, average shear stress was larger in clap half-stroke as compared to fling at
= 10 (
Figure 14D). The average shear stress decreased with increasing W/S. The average shear stress for W/S = 0.15 and 0.23 models showed only a small amount of change, similar to
for these two wing models (
Figure 14C).
Comparing
Figure 14C,D for a particular bristled wing model, it is clear that leakiness is inversely related to the average shear stress. This reinforces our previous suggestion that interacting high-shear regions formed around the bristles (e.g., W/S = 0.05 in
Figure 15 and
Figure A5A) can generate “inter-bristle blockage”, lowering the effective gap through which fluid can leak through, which consequently lowers
. The recirculating flow around the bristles for W/S = 0.15 and 0.23 wing models (
Figure 16 and
Figure 17) started to dissipate after
37.5% fling angle (
≈ 1.08). This time point was just prior to start of translation motion during fling (
≈ 1.1;
Figure 5B). The shear layers around the bristles were found to increase in strength with the onset of translation motion, as evidenced by an increase in
for
1.1 in
Figure 14D. This was particularly evident during clap (100% overlap of translation and rotation), where the shear layers around the bristles were stronger than during fling (
Figure 14D). The high-shear flow around the bristles thus directly contribute d to decrease in
. While pure rotation in fling increased
due to lower shear stress near the wing bristles (
= 1–1.1 in
Figure 14C,D), combining translational motion with rotation in clap increased shear stress and lowered
(
= 0.8–1.0 in
Figure 14C,D).



























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}