
Expression of Subtelomeric lncRNAs Links Telomeres Dynamics to RNA Decay in S. cerevisiae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
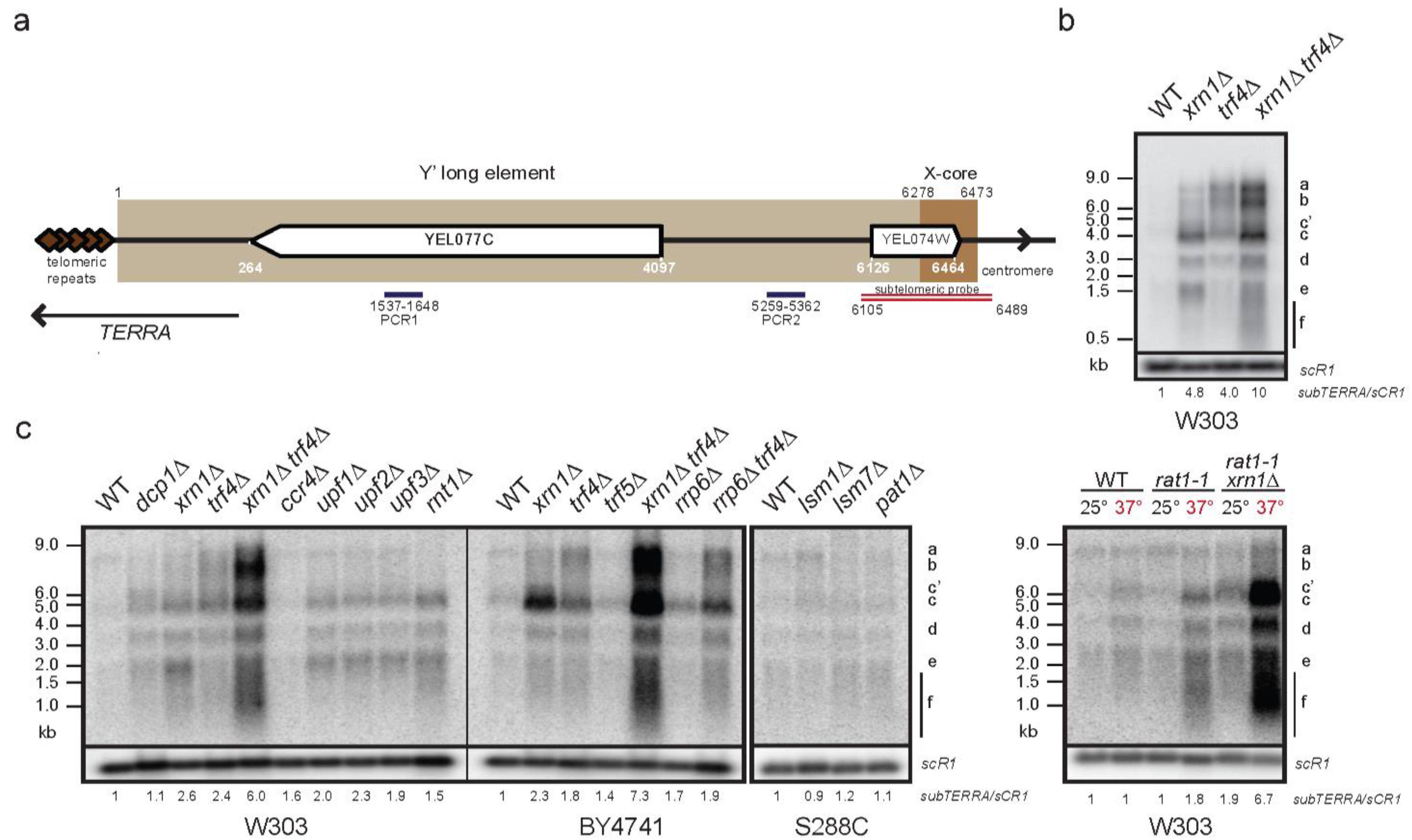
2.1. Characterization of the Subtelomeric Y′ ncRNAs (subTERRA)

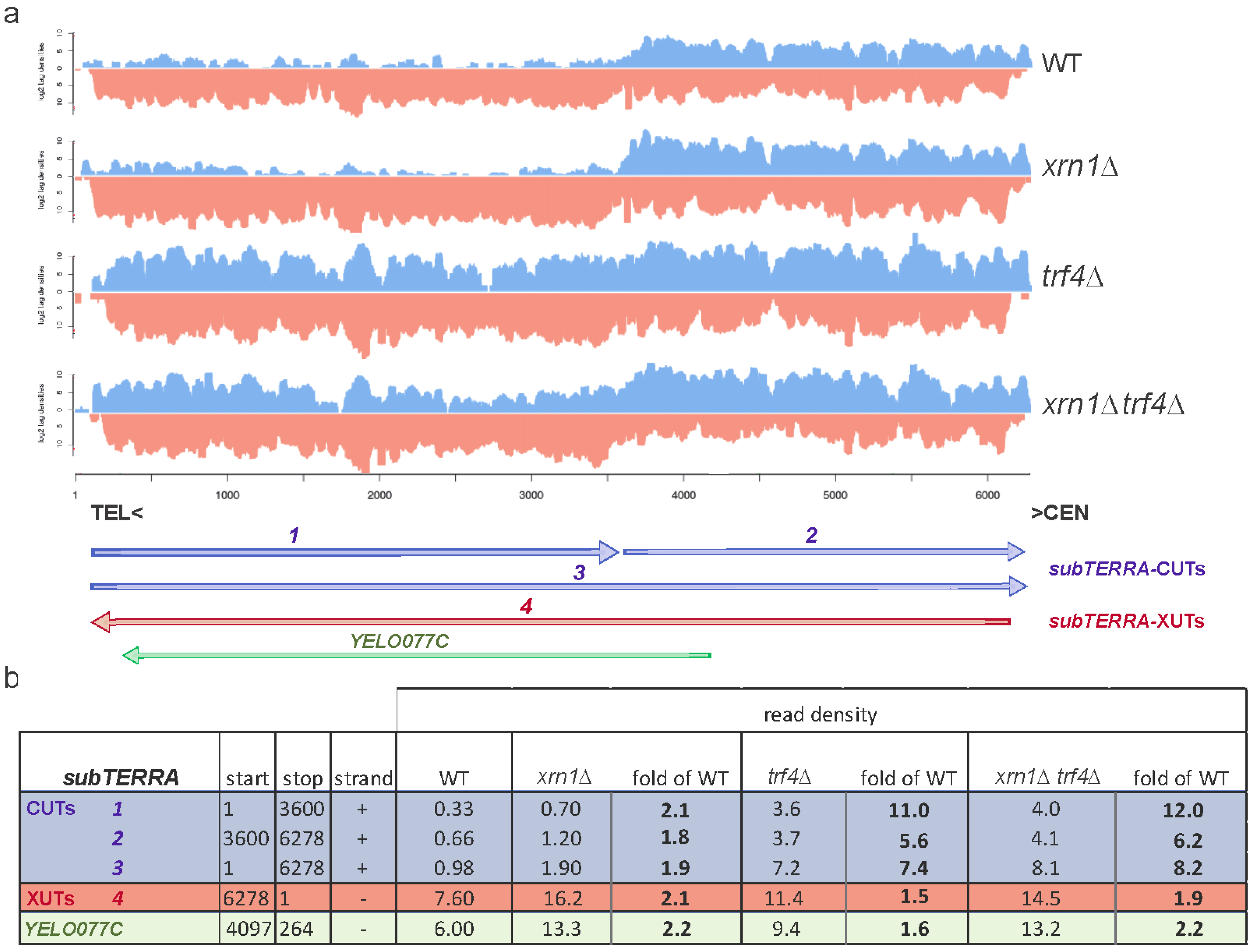
2.2. subTERRA-XUTs are Transcribed toward Telomeres and subTERRA-CUTs toward Centromeres

2.3. subTERRA Level is Mainly Regulated Post-Transcriptionally
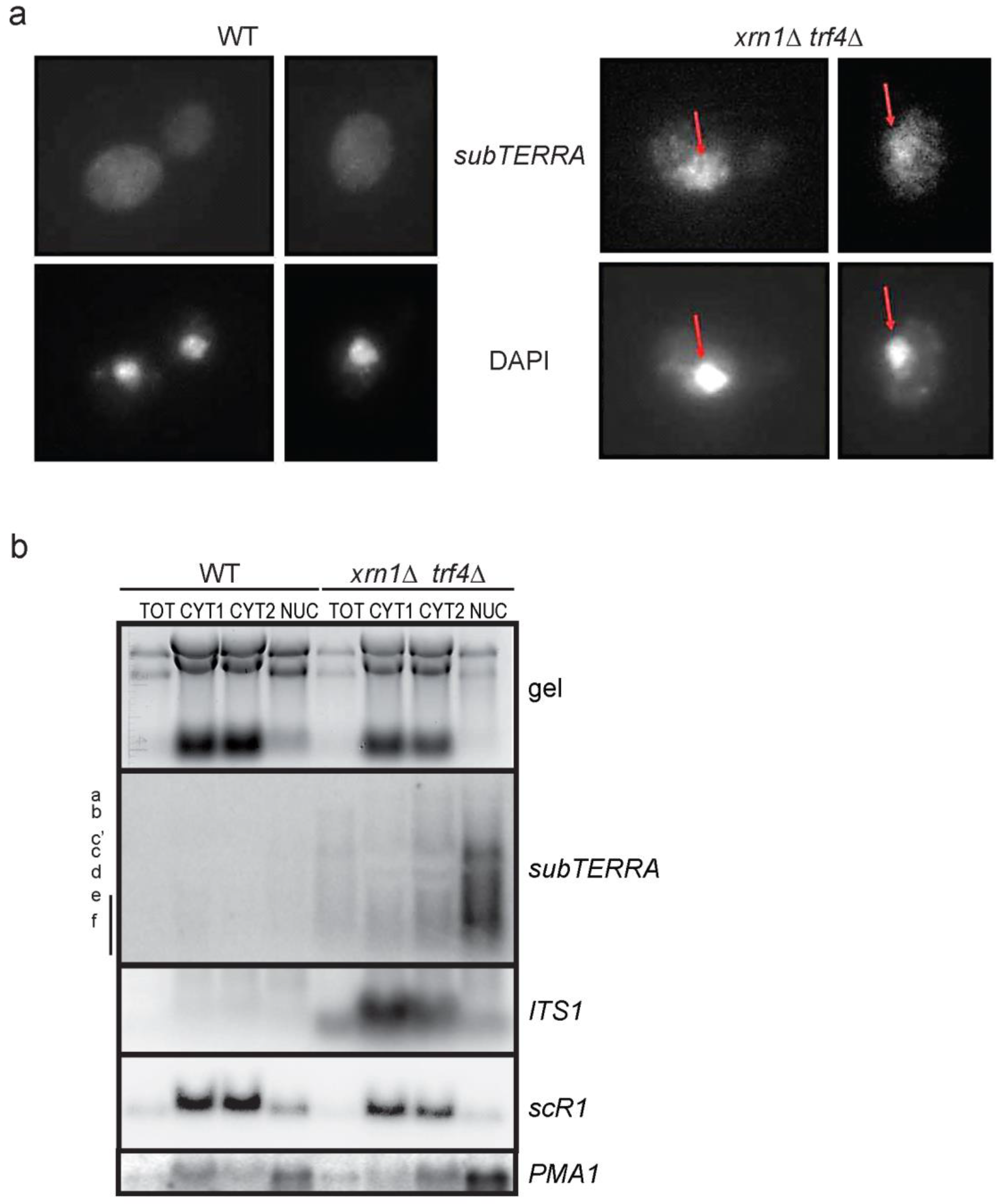
2.4. subTERRA Localizes to the Nucleus


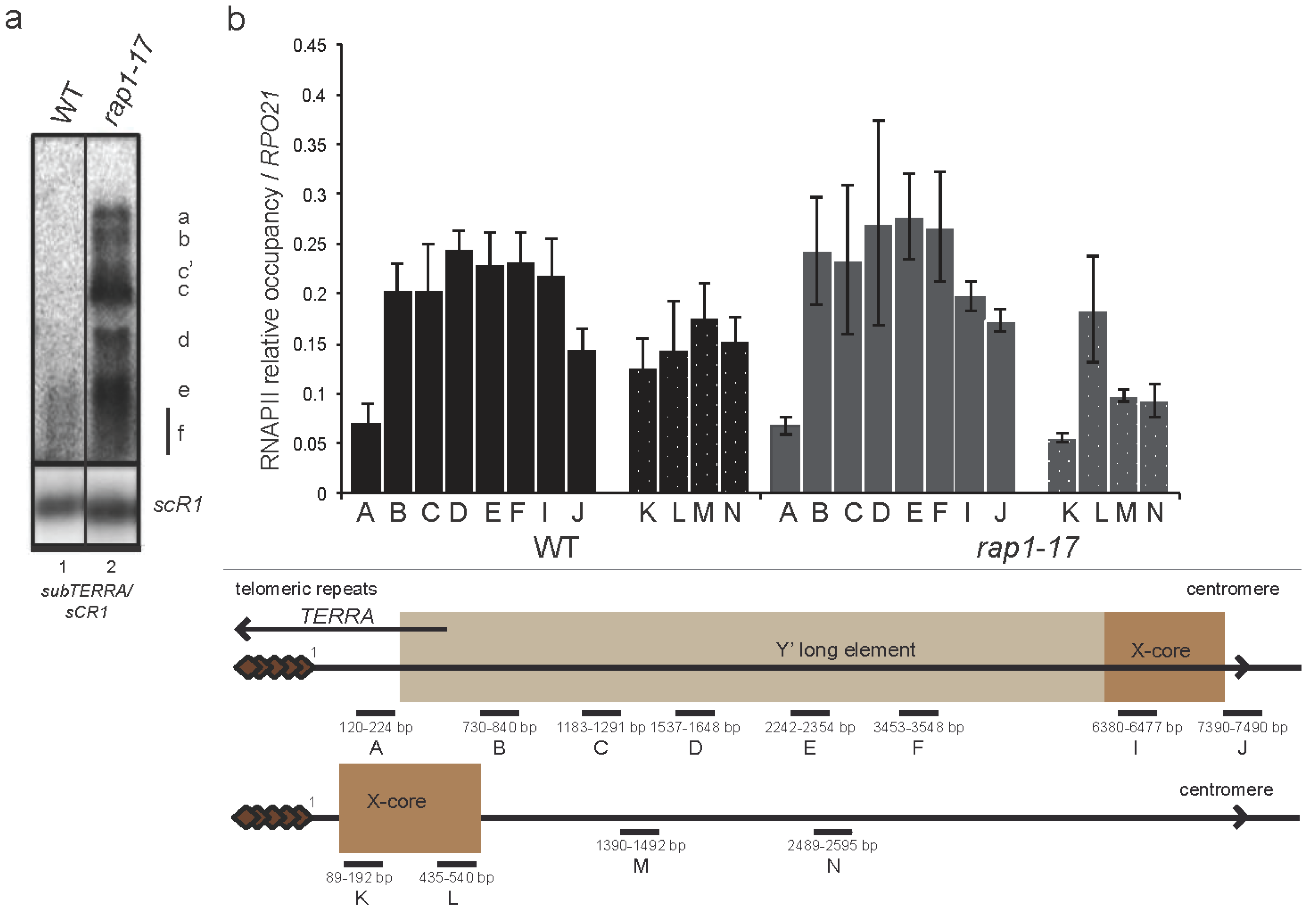
2.5. subTERRA Accumulates in rap1-17 Mutant
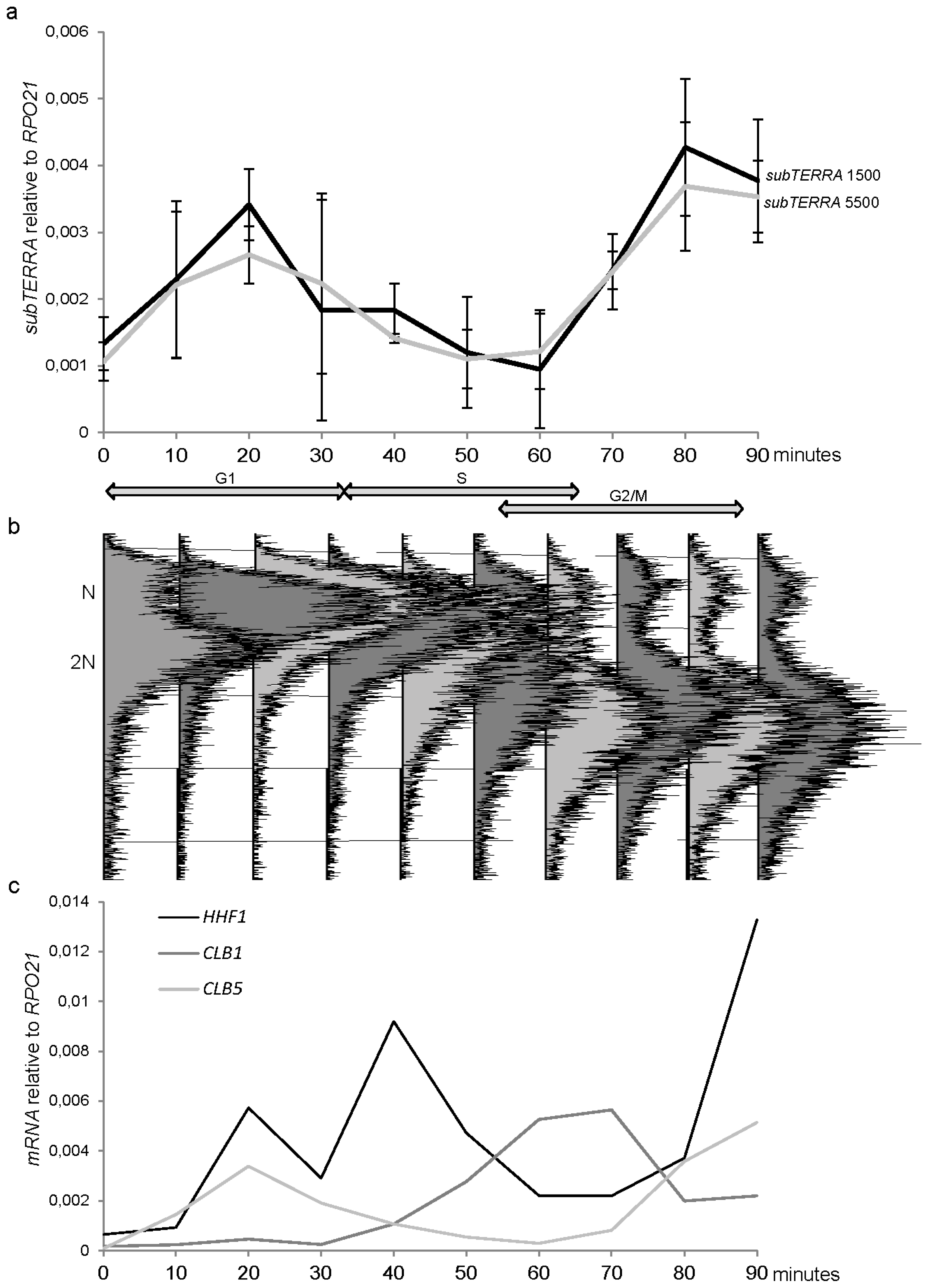
2.6. subTERRA Expression is Cell-Cycle Regulated


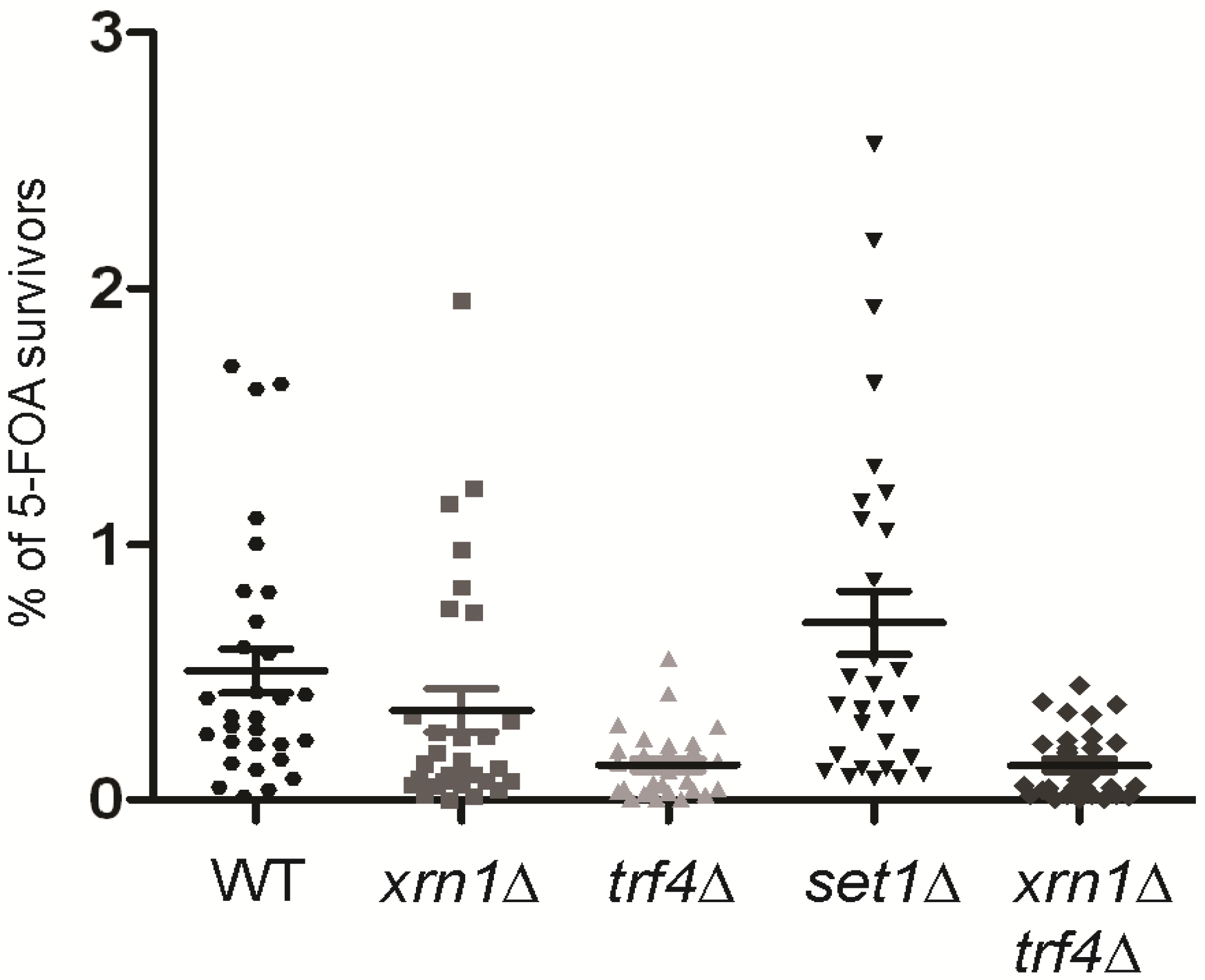
2.7. subTERRA-CUTs are Important for the Telomeric Silencing

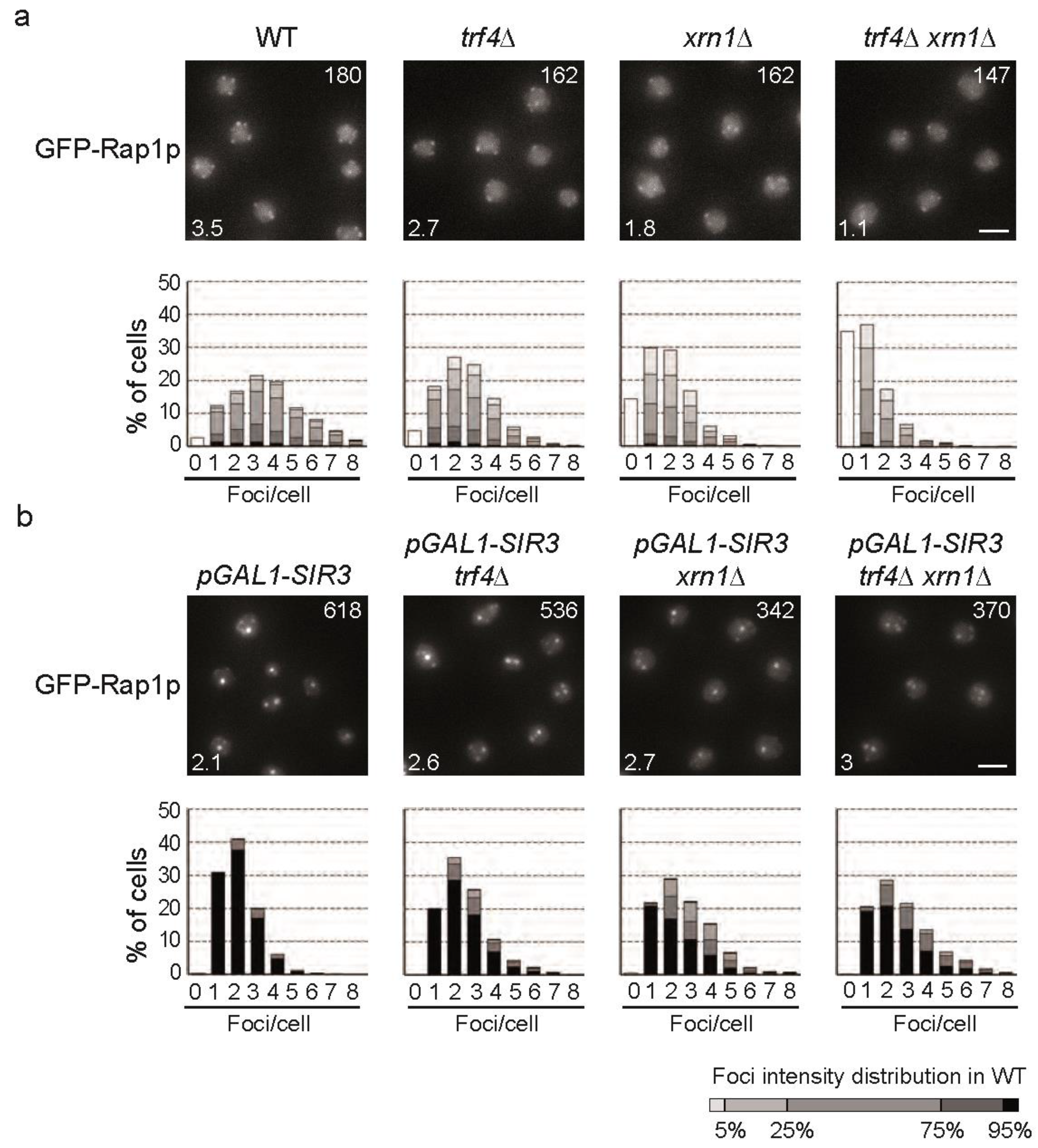
2.8. subTERRA-XUTs Accumulation Counteracts Telomeric Clustering

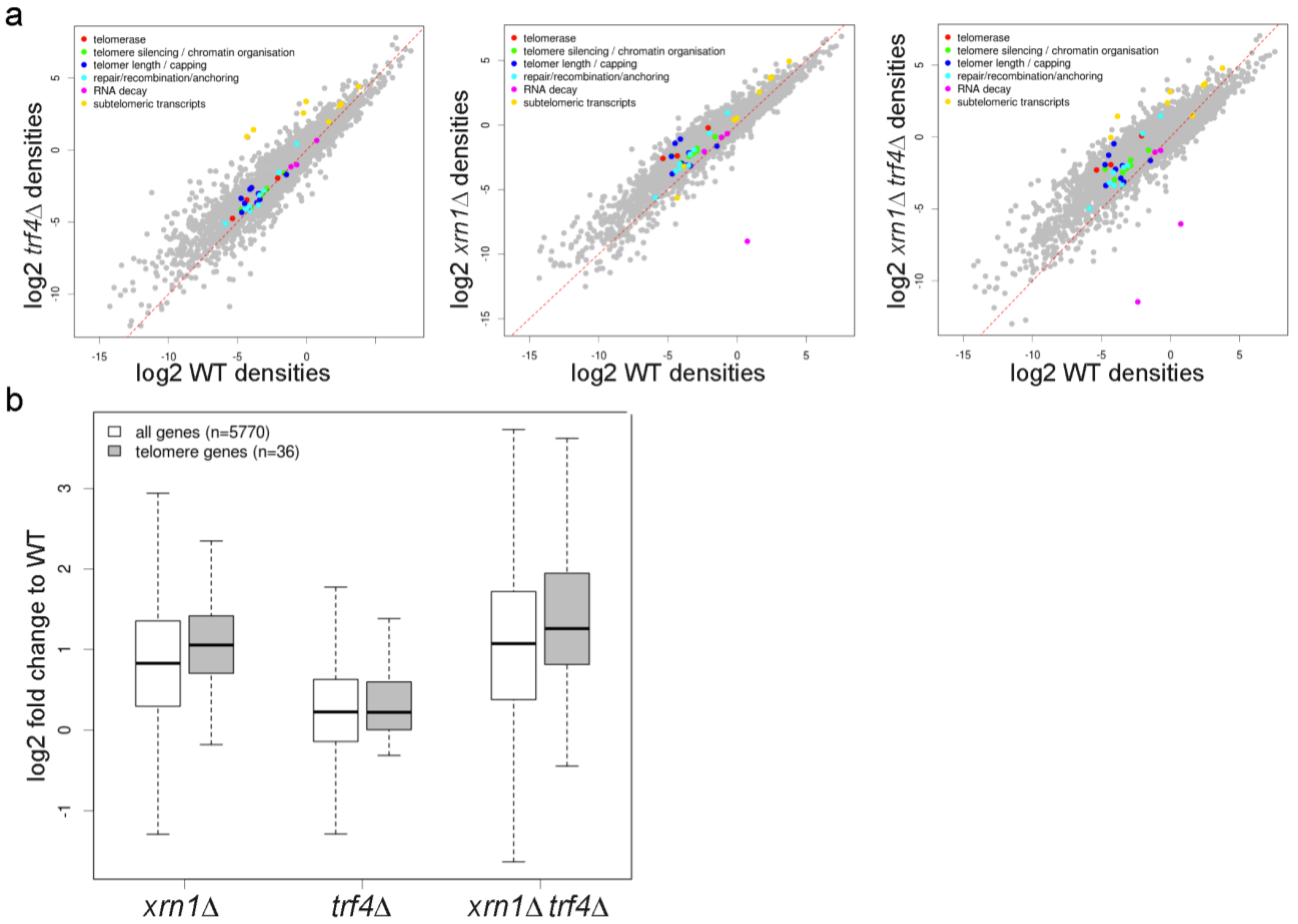
2.9. xrn1∆ and trf4∆ Mutations Change Global Expression Profiles but do not Affect Specifically Telomere Homeostasis Genes


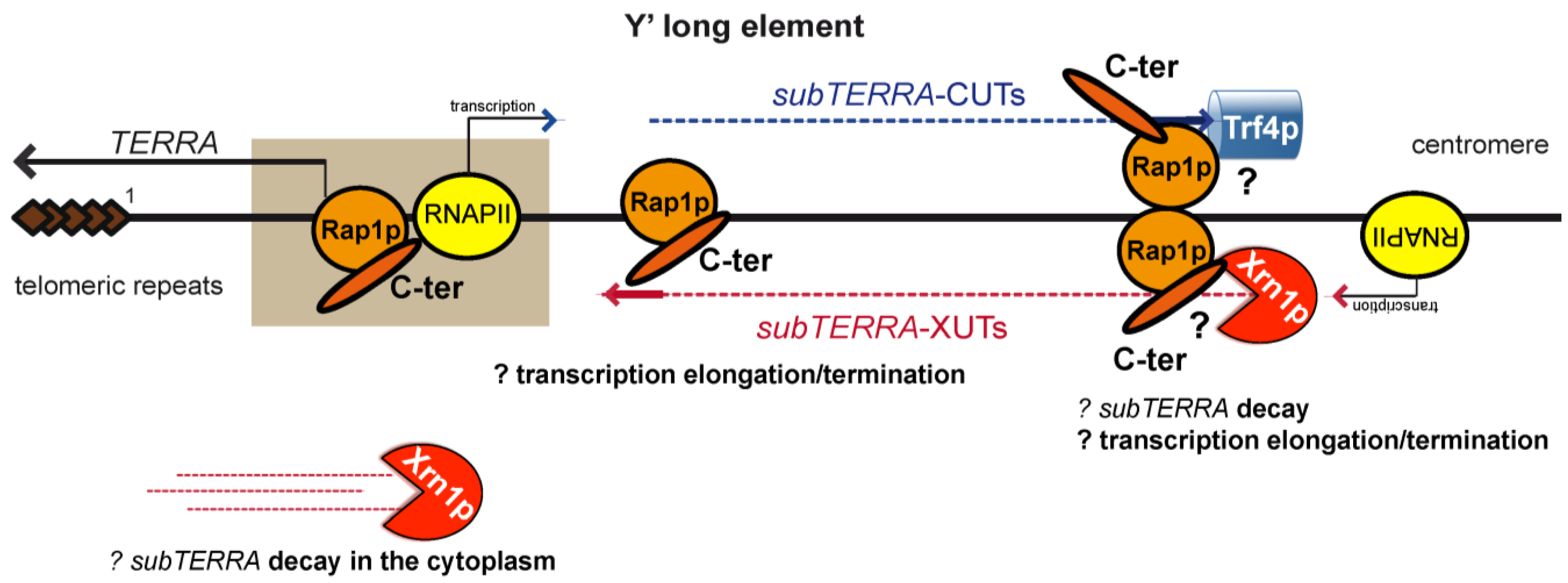
3. Discussion
3.1. Characterization and Subcellular Localization of subTERRA
3.2. subTERRA Expression and Mode of Action
3.3. Role of Rap1p in subTERRA Expression
4. Conclusions
5. Material and Methods
5.1. Yeast Strains and Plasmids
5.2. Media and Culture Conditions
5.3. RNA Extraction, poly(A)+ RNA Purification and Northern Blotting
5.4. Reverse Transcription and qPCR
5.5. RNA-seq
5.5.1. Preparation of Libraries
5.5.2. Data Mapping
5.5.3. Transcriptome Analysis and Normalization
5.6. Chromatin ImmunoPrecipitation (ChIP)
5.7. Fluorescence In Situ Hybridization (RNA-FISH)
5.8. Sub-Cellular Fractionation
5.9. Telomeric Clustering
5.10. Telomere Length Analysis
Supplementary Files
Supplementary File 1Supplementary File 2Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sandell, L.L.; Zakian, V.A. Loss of a yeast telomere: Arrest, recovery, and chromosome loss. Cell 1993, 75, 729–739. [Google Scholar] [CrossRef]
- D'Mello, N.P.; Jazwinski, S.M. Telomere length constancy during aging of saccharomyces cerevisiae. J. Bacteriol. 1991, 173, 6709–6713. [Google Scholar] [PubMed]
- Louis, E.J. The chromosome ends of saccharomyces cerevisiae. Yeast 1995, 11, 1553–1573. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.S.; Tye, B.K. A family of saccharomyces cerevisiae repetitive autonomously replicating sequences that have very similar genomic environments. J. Mol. Biol. 1983, 168, 505–523. [Google Scholar] [CrossRef]
- Chan, C.S.; Tye, B.K. Organization of DNA sequences and replication origins at yeast telomeres. Cell 1983, 33, 563–573. [Google Scholar] [CrossRef]
- Louis, E.J.; Haber, J.E. The structure and evolution of subtelomeric y' repeats in saccharomyces cerevisiae. Genetics 1992, 131, 559–574. [Google Scholar] [PubMed]
- Zakian, V.A.; Blanton, H.M.; Wetzel, L.; Dani, G.M. Size threshold for saccharomyces cerevisiae chromosomes: Generation of telocentric chromosomes from an unstable minichromosome. Mol. Cell Biol. 1986, 6, 925–932. [Google Scholar] [PubMed]
- Mak, H.C.; Pillus, L.; Ideker, T. Dynamic reprogramming of transcription factors to and from the subtelomere. Genome Res 2009, 19, 1014–1025. [Google Scholar] [CrossRef] [PubMed]
- Londono-Vallejo, J.A.; Wellinger, R.J. Telomeres and telomerase dance to the rhythm of the cell cycle. Trends Biochem. Sci. 2012, 37, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Gottschling, D.E.; Aparicio, O.M.; Billington, B.L.; Zakian, V.A. Position effect at s. Cerevisiae telomeres: Reversible repression of pol ii transcription. Cell 1990, 63, 751–762. [Google Scholar] [CrossRef]
- de Bruin, D.; Kantrow, S.M.; Liberatore, R.A.; Zakian, V.A. Telomere folding is required for the stable maintenance of telomere position effects in yeast. Mol. Cell Biol. 2000, 20, 7991–8000. [Google Scholar] [CrossRef] [PubMed]
- Guillemette, B.; Gaudreau, L. Reuniting the contrasting functions of h2a.Z. Biochem. Cell Biol. 2006, 84, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.M.; Brewer, B.J.; Reynolds, A.E.; Fangman, W.L. A yeast origin of replication is activated late in s phase. Cell 1991, 65, 507–515. [Google Scholar] [CrossRef]
- Bianchi, A.; Shore, D. Early replication of short telomeres in budding yeast. Cell 2007, 128, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Gilson, E.; Geli, V. How telomeres are replicated. Nat. Rev. Mol. Cell Biol. 2007, 8, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase activity in tetrahymena extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef]
- Lundblad, V.; Blackburn, E.H. An alternative pathway for yeast telomere maintenance rescues est1- senescence. Cell 1993, 73, 347–360. [Google Scholar] [CrossRef]
- Teng, S.C.; Zakian, V.A. Telomere-telomere recombination is an efficient bypass pathway for telomere maintenance in saccharomyces cerevisiae. Mol. Cell Biol. 1999, 19, 8083–8093. [Google Scholar] [PubMed]
- Teng, S.C.; Chang, J.; McCowan, B.; Zakian, V.A. Telomerase-independent lengthening of yeast telomeres occurs by an abrupt rad50p-dependent, rif-inhibited recombinational process. Mol. Cell 2000, 6, 947–952. [Google Scholar] [CrossRef]
- Dunham, M.A.; Neumann, A.A.; Fasching, C.L.; Reddel, R.R. Telomere maintenance by recombination in human cells. Nat. Genet 2000, 26, 447–450. [Google Scholar] [PubMed]
- Shore, D.; Nasmyth, K. Purification and cloning of a DNA binding protein from yeast that binds to both silencer and activator elements. Cell 1987, 51, 721–732. [Google Scholar] [CrossRef]
- Marcand, S.; Gilson, E.; Shore, D. A protein-counting mechanism for telomere length regulation in yeast. Science 1997, 275, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Moretti, P.; Freeman, K.; Coodly, L.; Shore, D. Evidence that a complex of sir proteins interacts with the silencer and telomere-binding protein rap1. Genes Dev. 1994, 8, 2257–2269. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Armstrong, C.M.; Kaeberlein, M.; Guarente, L. Transcriptional silencing and longevity protein sir2 is an nad-dependent histone deacetylase. Nature 2000, 403, 795–800. [Google Scholar] [PubMed]
- Aparicio, O.M.; Billington, B.L.; Gottschling, D.E. Modifiers of position effect are shared between telomeric and silent mating-type loci in s. Cerevisiae. Cell 1991, 66, 1279–1287. [Google Scholar] [CrossRef]
- Hecht, A.; Strahl-Bolsinger, S.; Grunstein, M. Spreading of transcriptional repressor sir3 from telomeric heterochromatin. Nature 1996, 383, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Rusche, L.N.; Kirchmaier, A.L.; Rine, J. The establishment, inheritance, and function of silenced chromatin in saccharomyces cerevisiae. Annu. Rev. Biochem. 2003, 72, 481–516. [Google Scholar] [CrossRef] [PubMed]
- van Leeuwen, F.; Gafken, P.R.; Gottschling, D.E. Dot1p modulates silencing in yeast by methylation of the nucleosome core. Cell 2002, 109, 745–756. [Google Scholar] [CrossRef]
- Santos-Rosa, H.; Bannister, A.J.; Dehe, P.M.; Geli, V.; Kouzarides, T. Methylation of h3 lysine 4 at euchromatin promotes sir3p association with heterochromatin. J. Biol. Chem. 2004, 279, 47506–47512. [Google Scholar] [CrossRef] [PubMed]
- Taddei, A.; Schober, H.; Gasser, S.M. The budding yeast nucleus. Cold Spring Harb. Perspect Biol. 2010, 2, a000612. [Google Scholar] [CrossRef] [PubMed]
- Gotta, M.; Laroche, T.; Formenton, A.; Maillet, L.; Scherthan, H.; Gasser, S.M. The clustering of telomeres and colocalization with rap1, sir3, and sir4 proteins in wild-type saccharomyces cerevisiae. J. Cell Biol. 1996, 134, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Taddei, A.; Hediger, F.; Neumann, F.R.; Bauer, C.; Gasser, S.M. Separation of silencing from perinuclear anchoring functions in yeast ku80, sir4 and esc1 proteins. Embo J. 2004, 23, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Laroche, T.; Martin, S.G.; Gotta, M.; Gorham, H.C.; Pryde, F.E.; Louis, E.J.; Gasser, S.M. Mutation of yeast ku genes disrupts the subnuclear organization of telomeres. Curr. Biol. 1998, 8, 653–656. [Google Scholar] [CrossRef]
- Galy, V.; Olivo-Marin, J.C.; Scherthan, H.; Doye, V.; Rascalou, N.; Nehrbass, U. Nuclear pore complexes in the organization of silent telomeric chromatin. Nature 2000, 403, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Ruault, M.; De Meyer, A.; Loiodice, I.; Taddei, A. Clustering heterochromatin: Sir3 promotes telomere clustering independently of silencing in yeast. J. Cell Biol. 2011, 192, 417–431. [Google Scholar] [CrossRef] [PubMed]
- Hoze, N.; Ruault, M.; Amoruso, C.; Taddei, A.; Holcman, D. Spatial telomere organization and clustering in yeast saccharomyces cerevisiae nucleus is generated by a random dynamics of aggregation-dissociation. Mol. Biol. Cell 2013, 24, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Therizols, P.; Duong, T.; Dujon, B.; Zimmer, C.; Fabre, E. Chromosome arm length and nuclear constraints determine the dynamic relationship of yeast subtelomeres. Proc. Natl. Acad. Sci. USA 2010, 107, 2025–2030. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally regulated transcription of mammalian telomeres by DNA-dependent rna polymerase ii. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric repeat containing rna and rna surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Luke, B.; Panza, A.; Redon, S.; Iglesias, N.; Li, Z.; Lingner, J. The rat1p 5' to 3' exonuclease degrades telomeric repeat-containing rna and promotes telomere elongation in saccharomyces cerevisiae. Mol. Cell 2008, 32, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Bah, A.; Gilson, E.; Wellinger, R.J. Telomerase is required to protect chromosomes with vertebrate-type t2ag3 3' ends in saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 27132–27138. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, N.; Redon, S.; Pfeiffer, V.; Dees, M.; Lingner, J.; Luke, B. Subtelomeric repetitive elements determine terra regulation by rap1/rif and rap1/sir complexes in yeast. EMBO Rep. 2011, 12, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. Terra rna binding to trf2 facilitates heterochromatin formation and orc recruitment at telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Berretta, J.; Pinskaya, M.; Morillon, A. A cryptic unstable transcript mediates transcriptional trans-silencing of the ty1 retrotransposon in s. Cerevisiae. Genes Dev. 2008, 22, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Camblong, J.; Iglesias, N.; Fickentscher, C.; Dieppois, G.; Stutz, F. Antisense rna stabilization induces transcriptional gene silencing via histone deacetylation in s. Cerevisiae. Cell 2007, 131, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Camblong, J.; Beyrouthy, N.; Guffanti, E.; Schlaepfer, G.; Steinmetz, L.M.; Stutz, F. Trans-acting antisense rnas mediate transcriptional gene cosuppression in s. Cerevisiae. Genes Dev. 2009, 23, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, E.L.; Chen, C.L.; d’Aubenton-Carafa, Y.; Gourvennec, S.; Kwapisz, M.; Roche, V.; Bertrand, C.; Silvain, M.; Legoix-Né, P.; Loeillet, S.; et al. Xuts are a class of xrn1-sensitive antisense regulatory non coding rna in yeast. Nature 2011, 475, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Gavalda, S.; Gallardo, M.; Luna, R.; Aguilera, A. R-loop mediated transcription-associated recombination in trf4delta mutants reveals new links between rna surveillance and genome integrity. PLoS ONE 2013, 8, e65541. [Google Scholar] [CrossRef] [PubMed]
- Manfrini, N.; Trovesi, C.; Wery, M.; Martina, M.; Cesena, D.; Descrimes, M.; Morillon, A.; d'Adda di Fagagna, F.; Longhese, M.P. Rna-processing proteins regulate mec1/atr activation by promoting generation of rpa-coated ssdna. EMBO Rep. 2015, 16, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Vasiljeva, L.; Kim, M.; Terzi, N.; Soares, L.M.; Buratowski, S. Transcription termination and rna degradation contribute to silencing of rna polymerase ii transcription within heterochromatin. Mol. Cell 2008, 29, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Houseley, J.; Kotovic, K.; El Hage, A.; Tollervey, D. Trf4 targets ncrnas from telomeric and rdna spacer regions and functions in rdna copy number control. EMBO J. 2007, 26, 4996–5006. [Google Scholar] [CrossRef] [PubMed]
- Danin-Kreiselman, M.; Lee, C.Y.; Chanfreau, G. Rnase iii-mediated degradation of unspliced pre-mrnas and lariat introns. Mol. Cell 2003, 11, 1279–1289. [Google Scholar] [CrossRef]
- Muhlrad, D.; Parker, R. Premature translational termination triggers mrna decapping. Nature 1994, 370, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Beelman, C.A.; Stevens, A.; Caponigro, G.; LaGrandeur, T.E.; Hatfield, L.; Fortner, D.M.; Parker, R. An essential component of the decapping enzyme required for normal rates of mrna turnover. Nature 1996, 382, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Castano, I.B.; Adams, C.; Vu, C.; Fitzhugh, D.; Christman, M.F. Structure/function analysis of the saccharomyces cerevisiae trf4/pol sigma DNA polymerase. Genetics 2002, 160, 381–391. [Google Scholar] [PubMed]
- Wyers, F.; Rougemaille, M.; Badis, G.; Rousselle, J.C.; Dufour, M.E.; Boulay, J.; Regnault, B.; Devaux, F.; Namane, A.; Seraphin, B.; et al. Cryptic pol ii transcripts are degraded by a nuclear quality control pathway involving a new poly(a) polymerase. Cell 2005, 121, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Neil, H.; Malabat, C.; d'Aubenton-Carafa, Y.; Xu, Z.; Steinmetz, L.M.; Jacquier, A. Widespread bidirectional promoters are the major source of cryptic transcripts in yeast. Nature 2009, 457, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Hayatsu, N.; Matsuura, A.; Ishikawa, F. Y'-help1, a DNA helicase encoded by the yeast subtelomeric y' element, is induced in survivors defective for telomerase. J. Biol. Chem. 1998, 273, 33360–33366. [Google Scholar] [CrossRef] [PubMed]
- Moy, T.I.; Silver, P.A. Nuclear export of the small ribosomal subunit requires the ran-gtpase cycle and certain nucleoporins. Genes Dev. 1999, 13, 2118–2133. [Google Scholar] [CrossRef] [PubMed]
- Shore, D.; Stillman, D.J.; Brand, A.H.; Nasmyth, K.A. Identification of silencer binding proteins from yeast: Possible roles in sir control and DNA replication. EMBO J. 1987, 6, 461–467. [Google Scholar] [PubMed]
- Pryde, F.E.; Louis, E.J. Limitations of silencing at native yeast telomeres. Embo J. 1999, 18, 2538–2550. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, M.P.; Luo, W.; Tsaponina, O.; Chabes, A.; Stillman, B. A common telomeric gene silencing assay is affected by nucleotide metabolism. Mol. Cell 2011, 42, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Lee, J.Y.; Abraham, V.; Chanoux, R.; Schoenly, K.A.; Johnson, F.B. Evidence that the s.Cerevisiae sgs1 protein facilitates recombinational repair of telomeres during senescence. Nucleic Acids Res. 2006, 34, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Schober, H.; Ferreira, H.; Kalck, V.; Gehlen, L.R.; Gasser, S.M. Yeast telomerase and the sun domain protein mps3 anchor telomeres and repress subtelomeric recombination. Genes Dev. 2009, 23, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Straatman, K.R.; Louis, E.J. Localization of telomeres and telomere-associated proteins in telomerase-negative saccharomyces cerevisiae. Chromosome Res. 2007, 15, 1033–1050. [Google Scholar] [CrossRef] [PubMed]
- Nagai, S.; Dubrana, K.; Tsai-Pflugfelder, M.; Davidson, M.B.; Roberts, T.M.; Brown, G.W.; Varela, E.; Hediger, F.; Gasser, S.M.; Krogan, N.J. Functional targeting of DNA damage to a nuclear pore-associated sumo-dependent ubiquitin ligase. Science 2008, 322, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Sheth, U.; Parker, R. Decapping and decay of messenger rna occur in cytoplasmic processing bodies. Science 2003, 300, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Houseley, J.; LaCava, J.; Tollervey, D. Rna-quality control by the exosome. Nat. Rev. Mol. Cell Biol. 2006, 7, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Switching and signaling at the telomere. Cell 2001, 106, 661–673. [Google Scholar] [CrossRef]
- Seto, A.G.; Zaug, A.J.; Sobel, S.G.; Wolin, S.L.; Cech, T.R. Saccharomyces cerevisiae telomerase is an sm small nuclear ribonucleoprotein particle. Nature 1999, 401, 177–180. [Google Scholar] [PubMed]
- Ferrezuelo, F.; Steiner, B.; Aldea, M.; Futcher, B. Biogenesis of yeast telomerase depends on the importin mtr10. Mol. Cell Biol. 2002, 22, 6046–6055. [Google Scholar] [CrossRef] [PubMed]
- Askree, S.H.; Yehuda, T.; Smolikov, S.; Gurevich, R.; Hawk, J.; Coker, C.; Krauskopf, A.; Kupiec, M.; McEachern, M.J. A genome-wide screen for saccharomyces cerevisiae deletion mutants that affect telomere length. Proc. Natl. Acad. Sci. USA 2004, 101, 8658–8663. [Google Scholar] [CrossRef] [PubMed]
- San Paolo, S.; Vanacova, S.; Schenk, L.; Scherrer, T.; Blank, D.; Keller, W.; Gerber, A.P. Distinct roles of non-canonical poly(a) polymerases in rna metabolism. PLoS Genet 2009, 5, e1000555. [Google Scholar] [CrossRef] [PubMed]
- Corda, Y.; Schramke, V.; Longhese, M.P.; Smokvina, T.; Paciotti, V.; Brevet, V.; Gilson, E.; Geli, V. Interaction between set1p and checkpoint protein mec3p in DNA repair and telomere functions. Nat. Genet 1999, 21, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, V.K.; Jones, C.I.; Newbury, S.F.; Green, P.J. Xrn 5' → 3' exoribonucleases: Structure, mechanisms and functions. Biochim. Biophys. Acta 2013, 1829, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Romero, C.A.; Chartrand, P. Telomeric noncoding rna terra is induced by telomere shortening to nucleate telomerase molecules at short telomeres. Mol. Cell 2013, 51, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, F.; Laterreur, N.; Cusanelli, E.; Ouenzar, F.; Querido, E.; Wellinger, R.J.; Chartrand, P. Live cell imaging of telomerase rna dynamics reveals cell cycle-dependent clustering of telomerase at elongating telomeres. Mol. Cell 2011, 44, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Porro, A.; Feuerhahn, S.; Reichenbach, P.; Lingner, J. Molecular dissection of telomeric repeat-containing rna biogenesis unveils the presence of distinct and multiple regulatory pathways. Mol. Cell Biol. 2010, 30, 4808–4817. [Google Scholar] [CrossRef] [PubMed]
- Marcand, S.; Brevet, V.; Mann, C.; Gilson, E. Cell cycle restriction of telomere elongation. Curr. Biol. 2000, 10, 487–490. [Google Scholar] [CrossRef]
- Wellinger, R.J.; Wolf, A.J.; Zakian, V.A. Origin activation and formation of single-strand TG1-3 tails occur sequentially in late S phase on a yeast linear plasmid. Mol. Cell. Biol. 1993, 7, 4057–4065. [Google Scholar]
- Ebrahimi, H.; Donaldson, A.D. Release of yeast telomeres from the nuclear periphery is triggered by replication and maintained by suppression of ku-mediated anchoring. Genes Dev. 2008, 22, 3363–3374. [Google Scholar] [CrossRef] [PubMed]
- Lundblad, V.; Szostak, J.W. A mutant with a defect in telomere elongation leads to senescence in yeast. Cell 1989, 57, 633–643. [Google Scholar] [CrossRef]
- Abdallah, P.; Luciano, P.; Runge, K.W.; Lisby, M.; Geli, V.; Gilson, E.; Teixeira, M.T. A two-step model for senescence triggered by a single critically short telomere. Nat. Cell Biol. 2009, 11, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Khadaroo, B.; Teixeira, M.T.; Luciano, P.; Eckert-Boulet, N.; Germann, S.M.; Simon, M.N.; Gallina, I.; Abdallah, P.; Gilson, E.; Geli, V.; et al. The DNA damage response at eroded telomeres and tethering to the nuclear pore complex. Nat. Cell Biol. 2009, 11, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, V.; Lingner, J. Terra promotes telomere shortening through exonuclease 1-mediated resection of chromosome ends. PLoS Genet 2012, 8, e1002747. [Google Scholar] [CrossRef] [PubMed]
- Maicher, A.; Lockhart, A.; Luke, B. Breaking new ground: Digging into terra function. Biochim. Biophys. Acta 2014, 1839, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Farnung, B.O.; Brun, C.M.; Arora, R.; Lorenzi, L.E.; Azzalin, C.M. Telomerase efficiently elongates highly transcribing telomeres in human cancer cells. PLoS ONE 2012, 7, e35714. [Google Scholar] [CrossRef] [PubMed]
- Maicher, A.; Kastner, L.; Dees, M.; Luke, B. Deregulated telomere transcription causes replication-dependent telomere shortening and promotes cellular senescence. Nucleic Acids Res. 2012, 40, 6649–6659. [Google Scholar] [CrossRef] [PubMed]
- Balk, B.; Dees, M.; Bender, K.; Luke, B. The differential processing of telomeres in response to increased telomeric transcription and rna-DNA hybrid accumulation. RNA Biol. 2014, 11, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. Rnaseh1 regulates terra-telomeric DNA hybrids and telomere maintenance in alt tumour cells. Nat. Commun. 2014, 5, 5220. [Google Scholar] [CrossRef] [PubMed]
- Fallet, E.; Jolivet, P.; Soudet, J.; Lisby, M.; Gilson, E.; Teixeira, M.T. Length-dependent processing of telomeres in the absence of telomerase. Nucleic Acids Res. 2014, 42, 3648–3665. [Google Scholar] [CrossRef] [PubMed]
- Lickwar, C.R.; Mueller, F.; Hanlon, S.E.; McNally, J.G.; Lieb, J.D. Genome-wide protein-DNA binding dynamics suggest a molecular clutch for transcription factor function. Nature 2012, 484, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Bregman, A.; Avraham-Kelbert, M.; Barkai, O.; Duek, L.; Guterman, A.; Choder, M. Promoter elements regulate cytoplasmic mrna decay. Cell 2011, 147, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.J.; Rothstein, R. Elevated recombination rates in transcriptionally active DNA. Cell 1989, 56, 619–630. [Google Scholar] [CrossRef]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical pcr-based gene deletion and modification in saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Janke, C.; Magiera, M.M.; Rathfelder, N.; Taxis, C.; Reber, S.; Maekawa, H.; Moreno-Borchart, A.; Doenges, G.; Schwob, E.; Schiebel, E.; et al. A versatile toolbox for pcr-based tagging of yeast genes: New fluorescent proteins, more markers and promoter substitution cassettes. Yeast 2004, 21, 947–962. [Google Scholar] [CrossRef] [PubMed]
- SGD. Available online: http://www.yeastgenome.org/ (accessed on 25 September 2011).
- Wery, M.; Descrimes, M.; Vogt, N.; Dallongeville, A.-S.; Gautheret, D.; Morillon, A. Double-stranded RNA formation protects lncRNA from NMD-mediated degradation. Moll. Cell. (under review).
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Kwapisz, M.; Wery, M.; Despres, D.; Ghavi-Helm, Y.; Soutourina, J.; Thuriaux, P.; Lacroute, F. Mutations of rna polymerase ii activate key genes of the nucleoside triphosphate biosynthetic pathways. EMBO J. 2008, 27, 2411–2421. [Google Scholar] [CrossRef] [PubMed]
- Louis, E.J.; Borts, R.H. A complete set of marked telomeres in saccharomyces cerevisiae for physical mapping and cloning. Genetics 1995, 139, 125–136. [Google Scholar] [PubMed]
- Gomes, P.; Sampaio-Marques, B.; Ludovico, P.; Rodrigues, F.; Leao, C. Low auxotrophy-complementing amino acid concentrations reduce yeast chronological life span. Mech. Ageing Dev. 2007, 128, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Cosnier, B.; Kwapisz, M.; Hatin, I.; Namy, O.; Hermann-Le Denmat, S.; Morillon, A.; Rousset, J.P.; Fabret, C. A viable hypomorphic allele of the essential imp3 gene reveals novel protein functions in saccharomyces cerevisiae. PLoS ONE 2011, 6, e19500. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.W. Rat1p and xrn1p are functionally interchangeable exoribonucleases that are restricted to and required in the nucleus and cytoplasm, respectively. Mol. Cell Biol. 1997, 17, 6122–6130. [Google Scholar] [PubMed]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [PubMed]
- He, F.; Li, X.; Spatrick, P.; Casillo, R.; Dong, S.; Jacobson, A. Genome-wide analysis of mrnas regulated by the nonsense-mediated and 5' to 3' mrna decay pathways in yeast. Mol. Cell 2003, 12, 1439–1452. [Google Scholar] [CrossRef]
- Egecioglu, D.E.; Henras, A.K.; Chanfreau, G.F. Contributions of trf4p- and trf5p-dependent polyadenylation to the processing and degradative functions of the yeast nuclear exosome. RNA 2006, 12, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Marcand, S.; Brevet, V.; Gilson, E. Progressive cis-inhibition of telomerase upon telomere elongation. EMBO J. 1999, 18, 3509–3519. [Google Scholar] [CrossRef] [PubMed]
- Bouveret, E.; Rigaut, G.; Shevchenko, A.; Wilm, M.; Seraphin, B. A sm-like protein complex that participates in mrna degradation. EMBO J. 2000, 19, 1661–1671. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwapisz, M.; Ruault, M.; Van Dijk, E.; Gourvennec, S.; Descrimes, M.; Taddei, A.; Morillon, A. Expression of Subtelomeric lncRNAs Links Telomeres Dynamics to RNA Decay in S. cerevisiae. Non-Coding RNA 2015, 1, 94-126. https://doi.org/10.3390/ncrna1020094
Kwapisz M, Ruault M, Van Dijk E, Gourvennec S, Descrimes M, Taddei A, Morillon A. Expression of Subtelomeric lncRNAs Links Telomeres Dynamics to RNA Decay in S. cerevisiae. Non-Coding RNA. 2015; 1(2):94-126. https://doi.org/10.3390/ncrna1020094
Chicago/Turabian StyleKwapisz, Marta, Myriam Ruault, Erwin Van Dijk, Stephanie Gourvennec, Marc Descrimes, Angela Taddei, and Antonin Morillon. 2015. "Expression of Subtelomeric lncRNAs Links Telomeres Dynamics to RNA Decay in S. cerevisiae" Non-Coding RNA 1, no. 2: 94-126. https://doi.org/10.3390/ncrna1020094
APA StyleKwapisz, M., Ruault, M., Van Dijk, E., Gourvennec, S., Descrimes, M., Taddei, A., & Morillon, A. (2015). Expression of Subtelomeric lncRNAs Links Telomeres Dynamics to RNA Decay in S. cerevisiae. Non-Coding RNA, 1(2), 94-126. https://doi.org/10.3390/ncrna1020094