
The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling

Abstract
:1. Introduction
2. Biogenesis of Housekeeping and Regulatory ncRNAs
2.1. Transfer RNA Derived Fragments (tRFs)
2.2. Small Interfering RNAs (siRNAs)
2.3. MicroRNAs (miRNAs)
2.4. Long Non-Coding RNAs (lncRNAs)
2.5. Circular RNAs (circRNAs)
2.6. piwi-Interacting RNAs (piRNAs)
3. Role of ncRNAs in Plant Growth and Development
3.1. Root Growth and Development
3.2. Leaf Development
3.3. Seed, Endosperm, and Seed Nutrient Development
4. Role of ncRNAs in Various Abiotic Stress Resilience
4.1. Drought Stress
4.2. Salt Stress
4.3. Flooding/Waterlogging/Hypoxia Stress
4.4. Heat and Cold Stress
4.5. Combined Stresses
5. Conclusions and Future Perspectives

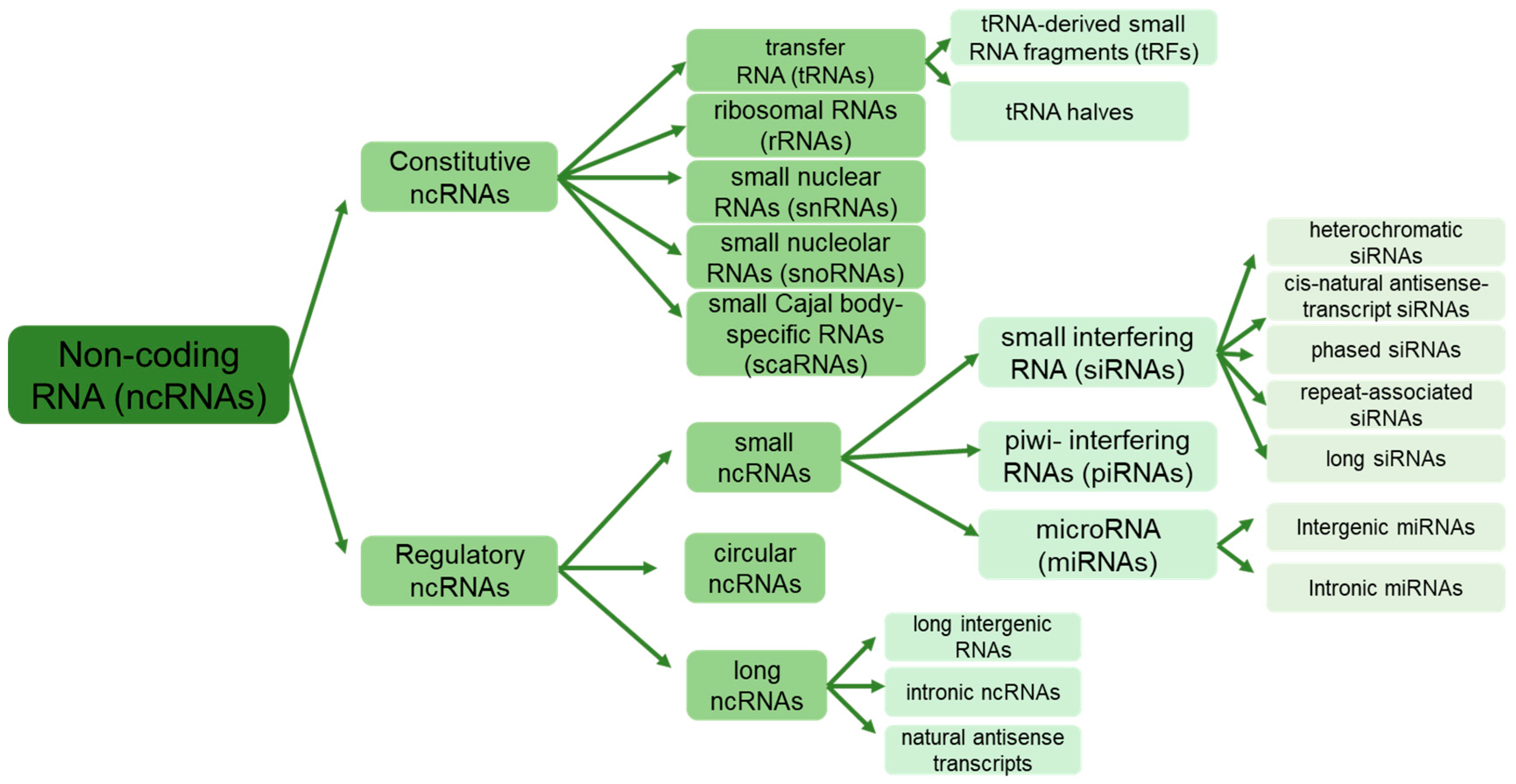
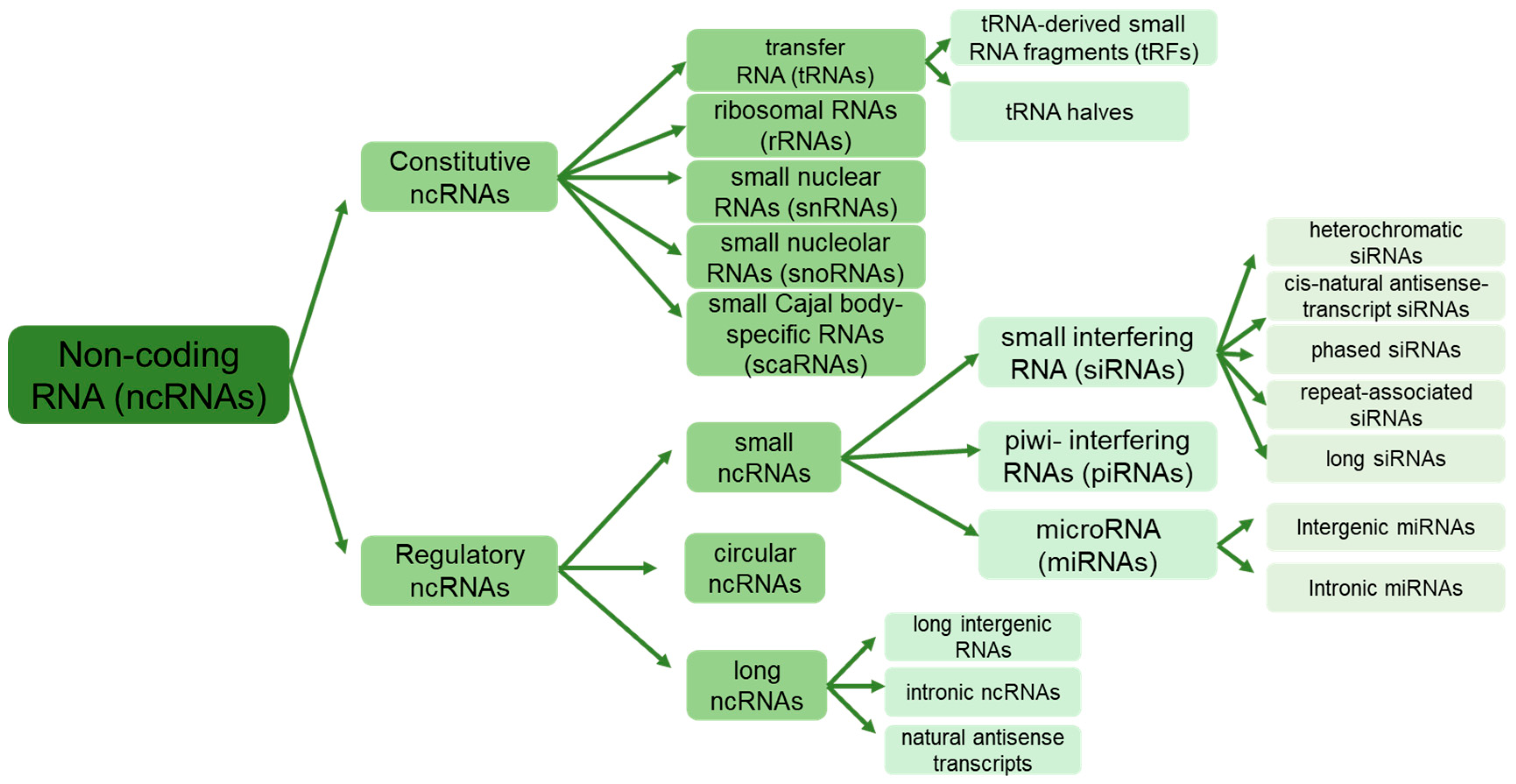{kind=link}
| Trait | siRNAs | Reference |
|---|---|---|
| Root development | TAS3-siRNA (ARFs) | Marin et al. [84] |
| TAS3-siRNA (leafbladeless1) | Gautam et al. [85] | |
| Leaf development | TAS3-siRNA (ARF3) | Chitwoodet al. [103] |
| TAS3-siRNA | Schwab et al. [104] | |
| TAS3-siRNA (leafbladeless1) | Dotto et al. [105] | |
| Seed, endosperm, nutrient development | maternal p4-siRNAs | Lu et al. [130] Yuan et al. [131] |
| Drought stress | Dehydration induced siRNAs | Jung et al. [161] |
| Yao et al. [162] | ||
| Salt stress | OsSIDP301 (stress-induced DUF1644 protein) | Ge et al. [182] |
| Response to ABA and Salt 1 | Ren et al. [183] | |
| OsSKL2 and OsASR | Jiang et al. [184] | |
| NATs of SRO5 and P5CDH | Borsani et al. [185] | |
| Flooding/hypoxia | hypoxia-induced tasiRNA | Moldovan et al. [194] |
| Heat stress | HEAT-INDUCED TAS1 TARGET1 | Li et al. [207] |
| Phased siRNA | Pokhrel et al. [208] | |
| Cold stress | cold-induced tasiRNA | Kume et al. [209] |
| miRNAs and corresponding targets | ||
| Root development | miR165/166-RLD1/2 | Gautam et al. [85] |
| miR156-SPL10 | Barrera-Rojas et al. [86] | |
| miR165-SHR | Carlsbecker et al. [87] | |
| Leaf development | miR319-TCP | Bresso et al. [106] |
| miR390-TAS3 | Husbands et al. [108] | |
| miR396-GRF | Omidbakhshfard et al. [124] | |
| Seed, endosperm, nutrient development | miR156, miR166, miR172, miR319, miR396 | Khemka et al. [132] |
| miR156-SPL14/SPL16 | Miao et al. [133] | |
| Drought stress | miR169-NFYA5 | Li et al. [128] |
| miR408a-Laccase | Jiao et al. [163] | |
| miRNA-mediated drought-responsive regulatory network | Balyan et al. [165] | |
| miR169-NFYA3 | Ni et al. [167] | |
| Salt stress | osa-miR12477-LAO | Parmar et al. [180] |
| Flooding/hypoxia | miR156/SPL | Feyissa et al. [199] |
| Heat stress | miR164-NAC | Tsai et al. [218] |
| Cold stress | miR319-TCP | Thiebaut et al. [225] |
| miR319-OsPCF6 and OsTCP21 | Wang et al. [226] | |
| lncRNAs | ||
| Root development | APOLO | Ariel et al. [89] Moison et al. [91] |
| lncWOX11a | Ran et al. [97] | |
| lncWOX5 | Qi et al. [100] | |
| Leaf development | TWISTED LEAF | Liu et al. [110] |
| age-related lncRNAs | Kim et al. [111] | |
| Seed, endosperm, nutrient development | LAIR | Wang et al. [134] |
| Drought stress | DRIR | Qin et al. [171] Dong et al. [172] |
| Salt stress | Salt-responsive DE-lncRNAs | Mansuri et al. [177] Mansuri et al. [187] |
| Genome-wide lncRNAs in Medicago truncatula | Wang et al. [188] | |
| Flooding/hypoxia | DE-lncRNAs in response to waterlogging stress | Yu et al. [196] Kęska et al. [202] |
| Heat stress | TCONS_00048391 and TCONS_00010856 | Wang et al. [235] |
| Cold stress | CIL1 | Liu et al. [61] |
| circRNAs | ||
| Root development | DE-circRNAs | Xu et al. [95] |
| Leaf development | circ-AT1G29965, circ-AT4G08300, circ-AT5G18590 | Liu et al. [112] |
| Seed, endosperm, nutrient development | DE-circRNAs participate in fatty acid metabolism | Zhou et al. [135] |
| Drought stress | Circular RNA profiling | Zhang et al. [173] |
| circGORK | Ache et al. [174] Becker et al. [175] | |
| Salt stress | circ_000260, circ_001362, and circ_001730 circRNA Chr10:20,345,844|20,346,873 | Liu et al. [189] Yin et al. [190] |
| Heat stress | 1583 heat-specific | Pan et al. [212] |
| Cold stress | 163 chilling responsive 1830 low temperatures induced | Zuo et al. [211] Yang et al. [210] |
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Hajieghrari, B.; Farrokhi, N. Plant RNA-Mediated Gene Regulatory Network. Genomics 2022, 114, 409–442. [Google Scholar] [CrossRef] [PubMed]
- Giambruno, R.; Mihailovich, M.; Bonaldi, T. Mass Spectrometry-Based Proteomics to Unveil the Non-Coding RNA World. Front. Mol. Biosci. 2018, 5, 90. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.M.; Chen, Y.Q. Principles and Innovative Technologies for Decrypting Noncoding RNAs: From Discovery and Functional Prediction to Clinical Application. J. Hematol. Oncol. 2020, 13, 109. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Zhu, W.; Sun, M.; Shi, L. Bioinformatics Analysis of Long Non-Coding RNA and Related Diseases: An Overview. Front. Genet. 2021, 12, 813873. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F. Is Junk DNA Bunk? A Critique of ENCODE. Proc. Natl. Acad. Sci. USA 2013, 110, 5294–5300. [Google Scholar] [CrossRef]
- Patra, G.K.; Gupta, D.; Rout, G.R.; Panda, S.K. Role of Long Non Coding RNA in Plants under Abiotic and Biotic Stresses. Plant Physiol. Biochem. 2023, 194, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and Their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef] [PubMed]
- Wajahat, M.; Bracken, C.P.; Orang, A. Emerging Functions for Snornas and Snorna-derived Fragments. Int. J. Mol. Sci. 2021, 22, 10193. [Google Scholar] [CrossRef]
- Alves, C.S.; Nogueira, F.T.S. Plant Small RNA World Growing Bigger: TRNA-Derived Fragments, Longstanding Players in Regulatory Processes. Front. Mol. Biosci. 2021, 8, 638911. [Google Scholar] [CrossRef]
- Waititu, J.K.; Zhang, C.; Liu, J.; Wang, H. Plant Non-Coding Rnas: Origin, Biogenesis, Mode of Action and Their Roles in Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 8401. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Fu, X.D. Non-Coding RNA: A New Frontier in Regulatory Biology. Natl. Sci. Rev. 2014, 1, 190–204. [Google Scholar] [CrossRef]
- Domínguez-Rosas, E.; Hernández-Oñate, M.Á.; Fernandez-Valverde, S.-L.; Tiznado-Hernández, M.E. Plant Long Non-Coding RNAs: Identification and Analysis to Unveil Their Physiological Functions. Front. Plant Sci. 2023, 14, 1275399. [Google Scholar] [CrossRef]
- Zhao, W.; Chu, S.; Jiao, Y. Present Scenario of Circular RNAs (CircRNAs) in Plants. Front. Plant Sci. 2019, 10, 379. [Google Scholar] [CrossRef]
- Zhang, P.; Li, S.; Chen, M. Characterization and Function of Circular RNAs in Plants. Front. Mol. Biosci. 2020, 7, 91. [Google Scholar] [CrossRef]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional Gene Regulation by Long Noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef]
- Sebastian-Delacruz, M.; Gonzalez-Moro, I.; Olazagoitia-Garmendia, A.; Castellanos-Rubio, A.; Santin, I. Non-Coding RNA The Role of LncRNAs in Gene Expression Regulation through MRNA Stabilization. Non-Coding RNA 2021, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, Y.; Zhu, W.; Liu, J.; Cheng, F. Non-Coding RNAs Fine-Tune the Balance between Plant Growth and Abiotic Stress Tolerance. Front. Plant Sci. 2022, 13, 965745. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Sze, C.C.; Li, M.W.; Lam, H.M. Roles of Non-Coding RNAs in the Hormonal and Nutritional Regulation in Nodulation and Nitrogen Fixation. Front. Plant Sci. 2022, 13, 997037. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, W.; Zhao, M.; Li, Z.; Wang, G.L.; Liu, W.; Liang, C. Transcriptome Analysis Reveals a LncRNA-MiRNA-MRNA Regulatory Network in OsRpp30-Mediated Disease Resistance in Rice. BMC Genom. 2023, 24, 643. [Google Scholar] [CrossRef] [PubMed]
- Hüttenhofer, A.; Vogel, J. Experimental Approaches to Identify Non-Coding RNAs. Nucleic Acids Res. 2006, 34, 635–646. [Google Scholar] [CrossRef]
- Singh, U.; Khemka, N.; Rajkumar, M.S.; Garg, R.; Jain, M. PLncPRO for Prediction of Long Non-Coding RNAs (LncRNAs) in Plants and Its Application for Discovery of Abiotic Stress-Responsive LncRNAs in Rice and Chickpea. Nucleic Acids Res. 2017, 45, e183. [Google Scholar] [CrossRef] [PubMed]
- Bhogireddy, S.; Mangrauthia, S.K.; Kumar, R.; Pandey, A.K.; Singh, S.; Jain, A.; Budak, H.; Varshney, R.K.; Kudapa, H. Regulatory Non-Coding RNAs: A New Frontier in Regulation of Plant Biology. Funct. Integr. Genom. 2021, 21, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Wang, X.; Duan, J.; Ma, J. Rhizobial TRNA-Derived Small RNAs Are Signal Molecules Regulating Plant Nodulation. Science 2019, 365, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Martinez, G.; Choudury, S.G.; Slotkin, R.K. TRNA-Derived Small RNAs Target Transposable Element Transcripts. Nucleic Acids Res. 2017, 45, 5142–5152. [Google Scholar] [CrossRef]
- Voinnet, O. Origin, Biogenesis, and Activity of Plant MicroRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef]
- Brosnan, C.A.; Voinnet, O. Cell-to-Cell and Long-Distance SiRNA Movement in Plants: Mechanisms and Biological Implications. Curr. Opin. Plant Biol. 2011, 14, 580–587. [Google Scholar] [CrossRef]
- Patil, V.S.; Zhou, R.; Rana, T.M. Gene Regulation by Non-Coding RNAs. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 16–32. [Google Scholar] [CrossRef]
- Mallory, A.C.; Vaucheret, H. Functions of Micrornas and Related Small a Rnas in Plants. Nat. Genet. 2006, 38, S31–S36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xia, J.; Lii, Y.E.; Barrera-Figueroa, B.E.; Zhou, X.; Gao, S.; Lu, L.; Niu, D.; Chen, Z.; Leung, C.; et al. Genome-Wide Analysis of Plant Nat-SiRNAs Reveals Insights into Their Distribution, Biogenesis and Function. Genome Biol. 2012, 13, R20. [Google Scholar] [CrossRef]
- Sanan-Mishra, N.; Abdul Kader Jailani, A.; Mandal, B.; Mukherjee, S.K. Secondary SiRNAs in Plants: Biosynthesis, Various Functions, and Applications in Virology. Front. Plant Sci. 2021, 12, 610283. [Google Scholar] [CrossRef]
- Zhang, X.; Lii, Y.; Wu, Z.; Polishko, A.; Zhang, H.; Chinnusamy, V.; Lonardi, S.; Zhu, J.K.; Liu, R.; Jin, H. Mechanisms of Small RNA Generation from Cis-NATs in Response to Environmental and Developmental Cues. Mol. Plant 2013, 6, 704–715. [Google Scholar] [CrossRef]
- Liu, Y.; Teng, C.; Xia, R.; Meyers, B.C. PhasiRNAs in Plants: Their Biogenesis, Genic Sources, and Roles in Stress Responses, Development, and Reproduction. Plant Cell 2020, 32, 3059–3080. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Muhammad, S.; Cao, M.; Wu, L. Biogenesis and Regulatory Hierarchy of Phased Small Interfering RNAs in Plants. Plant Biotechnol. J. 2018, 16, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Tao, X.; Li, M.; Gao, M.; Chen, J.; Zhou, N.; Mei, G.; Fang, L.; Ding, L.; Zhou, B.; et al. Role of PhasiRNAs from Two Distinct Phasing Frames of GhMYB2 Loci in Cis- Gene Regulation in the Cotton Genome. BMC Plant Biol. 2020, 20, 219. [Google Scholar] [CrossRef] [PubMed]
- Barber, W.T.; Zhang, W.; Win, H.; Varala, K.K.; Dorweiler, J.E.; Hudson, M.E.; Moose, S.P. Repeat Associated Small RNAs Vary among Parents and Following Hybridization in Maize. Proc. Natl. Acad. Sci. USA 2012, 109, 10444–10449. [Google Scholar] [CrossRef] [PubMed]
- Katiyar-Agarwal, S.; Gao, S.; Vivian-Smith, A.; Jin, H. A Novel Class of Bacteria-Induced Small RNAs in Arabidopsis. Genes Dev. 2007, 21, 3123–3134. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. MicroRNAs and Developmental Timing. Curr. Opin. Genet. Dev. 2011, 21, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of MiRNAs and SiRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Eulalio, A.; Huntzinger, E.; Izaurralde, E. Getting to the Root of MiRNA-Mediated Gene Silencing. Cell 2008, 132, 9–14. [Google Scholar] [CrossRef]
- Yadav, A.; Sanyal, I.; Rai, S.P.; Lata, C. An Overview on MiRNA-Encoded Peptides in Plant Biology Research. Genomics 2021, 113, 2385–2391. [Google Scholar] [CrossRef]
- Feng, Y.Z.; Zhu, Q.F.; Xue, J.; Chen, P.; Yu, Y. Shining in the Dark: The Big World of Small Peptides in Plants. aBIOTECH 2023, 4, 238–256. [Google Scholar] [CrossRef]
- Lauressergues, D.; Couzigou, J.M.; San Clemente, H.; Martinez, Y.; Dunand, C.; Bécard, G.; Combier, J.P. Primary Transcripts of MicroRNAs Encode Regulatory Peptides. Nature 2015, 520, 90–93. [Google Scholar] [CrossRef]
- Chen, Q.J.; Deng, B.H.; Gao, J.; Zhao, Z.Y.; Chen, Z.L.; Song, S.R.; Wang, L.; Zhao, L.P.; Xu, W.P.; Zhang, C.X.; et al. A Mirna-Encoded Small Peptide, Vvi-MiPEP171d1, Regulates Adventitious Root Formation. Plant Physiol. 2020, 183, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Badola, P.K.; Bhatia, C.; Sharma, D.; Trivedi, P.K. Primary Transcript of MiR858 Encodes Regulatory Peptide and Controls Flavonoid Biosynthesis and Development in Arabidopsis. Nat. Plants 2020, 6, 1262–1274. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Stellwag, E.J. Identification of Soybean MicroRNAs and Their Targets. Planta 2008, 229, 161–182. [Google Scholar] [CrossRef]
- Yang, T.; Wang, Y.; Teotia, S.; Wang, Z.; Shi, C.; Sun, H.; Gu, Y.; Zhang, Z.; Tang, G. The Interaction between MiR160 and MiR165/166 in the Control of Leaf Development and Drought Tolerance in Arabidopsis. Sci. Rep. 2019, 9, 2832. [Google Scholar] [CrossRef]
- Dong, Q.; Hu, B.; Zhang, C. MicroRNAs and Their Roles in Plant Development. Front. Plant Sci. 2022, 13, 824240. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.V.; Chekanova, J.A. Long Noncoding RNAs in Plants. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2017; Volume 1008, pp. 133–154. [Google Scholar]
- Wierzbicki, A.T.; Blevins, T.; Swiezewski, S. Annual Review of Plant Biology Long Noncoding RNAs in Plants. Annu. Rev. Plant Biol. 2021, 72, 2021. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhu, Q.H.; Kaufmann, K. Long Non-Coding RNAs in Plants: Emerging Modulators of Gene Activity in Development and Stress Responses. Planta 2020, 252, 92. [Google Scholar] [CrossRef]
- Bouba, I.; Kang, Q.; Luan, Y.S.; Meng, J. Predicting MiRNA-LncRNA Interactions and Recognizing Their Regulatory Roles in Stress Response of Plants. Math. Biosci. 2019, 312, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, A.; Yu, B.; Li, S. Interplay between MiRNAs and LncRNAs: Mode of Action and Biological Roles in Plant Development and Stress Adaptation. Comput. Struct. Biotechnol. J. 2021, 19, 2567–2574. [Google Scholar] [CrossRef]
- Kouchi, H.; Hata, S. Isolation and Characterization of Novel Nodulin CDNAs Representing Genes Expressed at Early Stages of Soybean Nodule Development. Mol. Gen. Genet. 1993, 238, 106–119. [Google Scholar] [CrossRef]
- Waseem, M.; Liu, Y.; Xia, R. Long Non-Coding RNAs, the Dark Matter: An Emerging Regulatory Component in Plants. Int. J. Mol. Sci. 2021, 22, 86. [Google Scholar] [CrossRef] [PubMed]
- Kindgren, P.; Ard, R.; Ivanov, M.; Marquardt, S. Transcriptional Read-through of the Long Non-Coding RNA SVALKA Governs Plant Cold Acclimation. Nat. Commun. 2018, 9, 4561. [Google Scholar] [CrossRef] [PubMed]
- Fedak, H.; Palusinska, M.; Krzyczmonik, K.; Brzezniak, L.; Yatusevich, R.; Pietras, Z.; Kaczanowski, S.; Swiezewski, S. Control of Seed Dormancy in Arabidopsis by a Cis-Acting Noncoding Antisense Transcript. Proc. Natl. Acad. Sci. USA 2016, 113, E7846–E7855. [Google Scholar] [CrossRef]
- Heo, J.B.; Sung, S. Vernalization-Mediated Epigenetic Silencing by a Long Intronic Noncoding RNA. Science 2011, 1197349, 331. [Google Scholar] [CrossRef]
- Kim, D.H.; Sung, S. Vernalization-Triggered Intragenic Chromatin Loop Formation by Long Noncoding RNAs. Dev. Cell 2017, 40, 302–312.e4. [Google Scholar] [CrossRef]
- Hawkes, E.J.; Hennelly, S.P.; Novikova, I.V.; Irwin, J.A.; Dean, C.; Sanbonmatsu, K.Y. COOLAIR Antisense RNAs Form Evolutionarily Conserved Elaborate Secondary Structures. Cell Rep. 2016, 16, 3087–3096. [Google Scholar] [CrossRef]
- Liu, G.; Liu, F.; Wang, Y.; Liu, X. A Novel Long Noncoding RNA CIL1 Enhances Cold Stress Tolerance in Arabidopsis. Plant Sci. 2022, 323, 111370. [Google Scholar] [CrossRef]
- Cui, J. Editorial: LncRNA in Plants: Function, Mechanisms and Applications. Front. Plant Sci. 2023, 14, 1238185. [Google Scholar] [CrossRef]
- Wen, J.; Lease, K.A.; Walker, J.C. DVL, a Novel Class of Small Polypeptides: Overexpression Alters Arabidopsis Development. Plant J. 2004, 37, 668–677. [Google Scholar] [CrossRef]
- Narita, N.N.; Moore, S.; Horiguchi, G.; Kubo, M.; Demura, T.; Fukuda, H.; Goodrich, J.; Tsukaya, H. Overexpression of a Novel Small Peptide Rotundifolia4 Decreases Cell Proliferation and Alters Leaf Shape in Arabidopsis Thaliana. Plant J. 2004, 38, 699–713. [Google Scholar] [CrossRef]
- Chilley, P.M.; Casson, S.A.; Tarkowski, P.; Hawkins, N.; Wang, K.L.C.; Hussey, P.J.; Beale, M.; Ecker, J.R.; Sandberg, G.K.; Lindsey, K. The POLARIS Peptide of Arabidopsis Regulates Auxin Transport Root Growth via Effects on Ethylene Signaling. Plant Cell 2006, 18, 3058–3072. [Google Scholar] [CrossRef]
- Lin, X.; Lin, W.; Ku, Y.S.; Wong, F.L.; Li, M.W.; Lam, H.M.; Ngai, S.M.; Chan, T.F. Analysis of Soybean Long Non-Coding RNAs Reveals a Subset of Small Peptide-Coding Transcripts1[OPEN]. Plant Physiol. 2020, 182, 1359–1374. [Google Scholar] [CrossRef]
- Zhao, X.; Lan, Y.; Chen, D. Exploring Long Non-Coding RNA Networks from Single Cell Omics Data. Comput. Struct. Biotechnol. J. 2022, 20, 4381–4389. [Google Scholar] [CrossRef]
- He, Z.; Luo, Y.; Zhou, X.; Zhu, T.; Lan, Y.; Chen, D. ScPlantDB: A Comprehensive Database for Exploring Cell Types and Markers of Plant Cell Atlases. Nucleic Acids Res. 2023, 52, D1629–D1638. [Google Scholar] [CrossRef]
- Salzman, J. Circular RNA Expression: Its Potential Regulation and Function. Trends Genet. 2016, 32, 309–316. [Google Scholar] [CrossRef]
- Patop, I.L.; Wüst, S.; Kadener, S. Past, Present, and Future of Circ RNA s. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Liu, S.; Guo, X.Y.; Shang, Q.J.; Gao, P. The Biogenesis, Biological Functions and Modification of Circular RNAs. Exp. Mol. Pathol. 2023, 131, 104861. [Google Scholar] [CrossRef]
- Feng, X.Y.; Zhu, S.X.; Pu, K.J.; Huang, H.J.; Chen, Y.Q.; Wang, W.T. New Insight into CircRNAs: Characterization, Strategies, and Biomedical Applications. Exp. Hematol. Oncol. 2023, 12, 91. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, Y.; Jia, E.; Liu, Z.; Pan, M.; Bai, Y.; Zhao, X.; Ge, Q. Comparative Analysis of Circular RNA Enrichment Methods. RNA Biol. 2022, 19, 55–67. [Google Scholar] [CrossRef]
- Long, F.; Lin, Z.; Li, L.; Ma, M.; Lu, Z.; Jing, L.; Li, X.; Lin, C. Comprehensive Landscape and Future Perspectives of Circular RNAs in Colorectal Cancer. Mol. Cancer 2021, 20, 26. [Google Scholar] [CrossRef]
- Ye, C.Y.; Zhang, X.; Chu, Q.; Liu, C.; Yu, Y.; Jiang, W.; Zhu, Q.H.; Fan, L.; Guo, L. Full-Length Sequence Assembly Reveals Circular RNAs with Diverse Non-GT/AG Splicing Signals in Rice. RNA Biol. 2017, 14, 1055–1063. [Google Scholar] [CrossRef]
- Olesen, M.T.J.; Kristensen, L.S. Circular RNAs as MicroRNA Sponges: Evidence and Controversies. Essays Biochem. 2021, 65, 685–696. [Google Scholar]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. CircRNA Biogenesis Competes with Pre-MRNA Splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Lukiw, W.J. Circular RNA (CircRNA) in Alzheimer’s Disease (AD). Front. Genet. 2013, 4, 307. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, S.; Liu, K. Structural Insights into PiRNA Biogenesis. Biochim. Biophys. Acta Gene Regul. Mech. 2022, 1865, 194799. [Google Scholar] [CrossRef]
- Huang, S.; Yoshitake, K.; Asakawa, S. A Review of Discovery Profiling of Piwi-Interacting Rnas and Their Diverse Functions in Metazoans. Int. J. Mol. Sci. 2021, 22, 11166. [Google Scholar] [CrossRef]
- Tian, H.; Jia, Y.; Niu, T.; Yu, Q.; Ding, Z. The Key Players of the Primary Root Growth and Development Also Function in Lateral Roots in Arabidopsis. Plant Cell Rep. 2014, 33, 745–753. [Google Scholar] [CrossRef]
- Smet, W.; Blilou, I. A Blast from the Past: Understanding Stem Cell Specification in Plant Roots Using Laser Ablation. Quant. Plant Biol. 2023, 4, e14. [Google Scholar] [CrossRef] [PubMed]
- Jansen, L.; Roberts, I.; de Rycke, R.; Beeckman, T. Phloem-Associated Auxin Response Maxima Determine Radial Positioning of Lateral Roots in Maize. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1525–1533. [Google Scholar] [CrossRef]
- Marin, E.; Jouannet, V.; Herz, A.; Lokerse, A.S.; Weijers, D.; Vaucheret, H.; Nussaume, L.; Crespi, M.D.; Maizel, A. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell 2010, 22, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Gautam, V.; Singh, A.; Yadav, S.; Singh, S.; Kumar, P.; Das, S.S.; Sarkar, A.K. Conserved LBL1-Ta-SiRNA and MiR165/166-RLD1/2 Modules Regulate Root Development in Maize. Development 2021, 148, dev190033. [Google Scholar] [CrossRef]
- Barrera-Rojas, C.H.; Rocha, G.H.B.; Polverari, L.; Di Pinheiro Brito, A.; Di Batista, S.; Notini, M.M.; Da Cruz, A.C.F.; Morea, E.G.O.; Sabatini, S.; Otoni, W.C.; et al. MiR156-Targeted SPL10 Controls Arabidopsis Root Meristem Activity and Root-Derived de Novo Shoot Regeneration via Cytokinin Responses. J. Exp. Bot. 2020, 71, 934–950. [Google Scholar] [CrossRef]
- Carlsbecker, A.; Lee, J.Y.; Roberts, C.J.; Dettmer, J.; Lehesranta, S.; Zhou, J.; Lindgren, O.; Moreno-Risueno, M.A.; Vatén, A.; Thitamadee, S.; et al. Cell Signalling by MicroRNA165/6 Directs Gene Dose-Dependent Root Cell Fate. Nature 2010, 465, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Xie, Y.; Guo, F.; Han, N.; Ma, S.; Zeng, Z.; Wang, J.; Yang, Y.; Zhu, M. Distinctive Expression Patterns and Roles of the MiRNA393/TIR1 Homolog Module in Regulating Flag Leaf Inclination and Primary and Crown Root Growth in Rice (Oryza Sativa). New Phytol. 2012, 196, 149–161. [Google Scholar] [CrossRef]
- Ariel, F.; Lucero, L.; Christ, A.; Mammarella, M.F.; Jegu, T.; Veluchamy, A.; Mariappan, K.; Latrasse, D.; Blein, T.; Liu, C.; et al. R-Loop Mediated Trans Action of the APOLO Long Noncoding RNA. Mol. Cell 2020, 77, 1055–1065.e4. [Google Scholar] [CrossRef]
- Rigo, R.; Bazin, J.; Romero-Barrios, N.; Moison, M.; Lucero, L.; Christ, A.; Benhamed, M.; Blein, T.; Huguet, S.; Charon, C.; et al. The Arabidopsis Lnc RNA ASCO Modulates the Transcriptome through Interaction with Splicing Factors. EMBO Rep. 2020, 21, e48977. [Google Scholar] [CrossRef]
- Moison, M.; Pacheco, J.M.; Lucero, L.; Fonouni-Farde, C.; Rodríguez-Melo, J.; Mansilla, N.; Christ, A.; Bazin, J.; Benhamed, M.; Ibañez, F.; et al. The LncRNA APOLO Interacts with the Transcription Factor WRKY42 to Trigger Root Hair Cell Expansion in Response to Cold. Mol. Plant 2021, 14, 937–948. [Google Scholar] [CrossRef]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.S.; Crespi, M. Long Noncoding RNA Modulates Alternative Splicing Regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef]
- Bazin, J.; Romero, N.; Rigo, R.; Charon, C.; Blein, T.; Ariel, F.; Crespi, M. Nuclear Speckle Rna Binding Proteins Remodel Alternative Splicing and the Non-Coding Arabidopsis Transcriptome to Regulate a Cross-Talk between Auxin and Immune Responses. Front. Plant Sci. 2018, 9, 1209. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, J.M.; Mansilla, N.; Moison, M.; Lucero, L.; Gabarain, V.B.; Ariel, F.; Estevez, J.M. The LncRNA APOLO and the Transcription Factor WRKY42 Target Common Cell Wall EXTENSIN Encoding Genes to Trigger Root Hair Cell Elongation. Plant Signal. Behav. 2021, 16, 1920191. [Google Scholar] [CrossRef]
- Xu, Y.; Ren, Y.; Lin, T.; Cui, D. Identification and Characterization of CircRNAs Involved in the Regulation of Wheat Root Length. Biol. Res. 2019, 52, 19. [Google Scholar] [CrossRef]
- Zhang, D.-J.; Yang, Y.-J.; Liu, C.-Y.; Zhang, F.; Wu, Q.-S. Root Hair Growth and Development in Response to Nutrients and Phytohormones. Root Biol. 2018, 52, 65–84. [Google Scholar]
- Ran, N.; Liu, S.; Qi, H.; Wang, J.; Shen, T.; Xu, W.; Xu, M. Long Non-Coding RNA LncWOX11a Suppresses Adventitious Root Formation of Poplar by Regulating the Expression of PeWOX11a. Int. J. Mol. Sci. 2023, 24, 5766. [Google Scholar] [CrossRef]
- Sundararajan, S.; Sivakumar, H.P.; Rajendran, V.; Kumariah, M.; Ramalingam, S. Adventitious Roots in Rice, the Model Cereal: Genetic Factors and the Influence of Environmental Cues—A Mini Review. Plant Cell Tissue Organ Cult. 2023, 154, 1–12. [Google Scholar] [CrossRef]
- Liu, R.; Wen, S.S.; Sun, T.T.; Wang, R.; Zuo, W.T.; Yang, T.; Wang, C.; Hu, J.J.; Lu, M.Z.; Wang, L.Q. PagWOX11/12a Positively Regulates the PagSAUR36 Gene That Enhances Adventitious Root Development in Poplar. J. Exp. Bot. 2022, 73, 7298–7311. [Google Scholar] [CrossRef]
- Qi, H.; Wu, L.; Shen, T.; Liu, S.; Cai, H.; Ran, N.; Wang, J.; Xu, M. Overexpression of the Long Non-Coding RNA LncWOX5 Negatively Regulates the Development of Adventitious Roots in Populus. Ind. Crops Prod. 2023, 192, 116054. [Google Scholar] [CrossRef]
- Paul Alvarez, J.; Furumizu, C.; Efroni, I.; Eshed, Y.; Bowman, J.L. Active Suppression of a Leaf Meristem Orchestrates Determinate Leaf Growth. eLife 2016, 5, e15023. [Google Scholar] [CrossRef]
- Yang, T.; Wang, Y.; Teotia, S.; Zhang, Z.; Tang, G. The Making of Leaves: How Small RNA Networks Modulate Leaf Development. Front. Plant Sci. 2018, 9, 824. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Nogueira, F.T.; Howell, M.D.; Montgomery, T.A.; Carrington, J.C.; Timmermans, M.C. Pattern formation via small RNA mobility. Genes Dev. 2009, 1, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Schwab, R.; Maizel, A.; Ruiz-Ferrer, V.; Garcia, D.; Bayer, M.; Crespi, M.; Voinnet, O.; Martienssen, R.A. Endogenous TasiRNAs Mediate Non-Cell Autonomous Effects on Gene Regulation in Arabidopsis thaliana. PLoS ONE 2009, 4, e5980. [Google Scholar] [CrossRef] [PubMed]
- Dotto, M.C.; Petsch, K.A.; Aukerman, M.J.; Beatty, M.; Hammell, M.; Timmermans, M.C.P. Genome-Wide Analysis of Leafbladeless1-Regulated and Phased Small RNAs Underscores the Importance of the TAS3 Ta-SiRNA Pathway to Maize Development. PLoS Genet. 2014, 10, e1004826. [Google Scholar] [CrossRef] [PubMed]
- Bresso, E.G.; Chorostecki, U.; Rodriguez, R.E.; Palatnik, J.F.; Schommer, C. Spatial Control of Gene Expression by MiR319-Regulated TCP Transcription Factors in Leaf Development. Plant Physiol. 2018, 176, 1694–1708. [Google Scholar] [CrossRef]
- Gupta, M.D.; Nathut, U. Divergence in Patterns of Leaf Growth Polarity Is Associated with the Expression Divergence of MiR396. Plant Cell 2015, 27, 2785–2799. [Google Scholar] [CrossRef]
- Husbands, A.Y.; Chitwood, D.H.; Plavskin, Y.; Timmermans, M.C.P. Signals and Prepatterns: New Insights into Organ Polarity in Plants. Genes Dev. 2009, 23, 1986–1997. [Google Scholar] [CrossRef]
- Schommer, C.; Debernardi, J.M.; Bresso, E.G.; Rodriguez, R.E.; Palatnik, J.F. Repression of Cell Proliferation by MiR319-Regulated TCP4. Mol. Plant 2014, 7, 1533–1544. [Google Scholar] [CrossRef]
- Liu, X.; Li, D.; Zhang, D.; Yin, D.; Zhao, Y.; Ji, C.; Zhao, X.; Li, X.; He, Q.; Chen, R.; et al. A Novel Antisense Long Noncoding RNA, TWISTED LEAF, Maintains Leaf Blade Flattening by Regulating Its Associated Sense R2R3-MYB Gene in Rice. New Phytol. 2018, 218, 774–788. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, J.; Kang, M.H.; Trang, T.T.M.; Lee, J.; Lee, H.; Jeong, H.; Lim, P.O. Dynamic Landscape of Long Noncoding RNAs during Leaf Aging in Arabidopsis. Front. Plant Sci. 2022, 13, 1068163. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Chen, G.; Shi, T. Identifying and Characterizing the Circular RNAs during the Lifespan of Arabidopsis Leaves. Front. Plant Sci. 2017, 8, 1278. [Google Scholar] [CrossRef]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial Patterning of Arabidopsis Shoots by Class III HD-ZIP and KANADI Genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef]
- Liu, D.; Song, Y.; Chen, Z.; Yu, D. Ectopic Expression of MiR396 Suppresses GRF Target Gene Expression and Alters Leaf Growth in Arabidopsis. Physiol. Plant. 2009, 136, 223–236. [Google Scholar] [CrossRef]
- Nikovics, K.; Blein, T.; Peaucelle, A.; Ishida, T.; Morin, H.; Aida, M.; Laufs, P. The Balance between the MIR164A and CUC2 Genes Controls Leaf Margin Serration in Arabidopsis. Plant Cell 2006, 18, 2929–2945. [Google Scholar] [CrossRef]
- Sieber, P.; Wellmer, F.; Gheyselinck, J.; Riechmann, J.L.; Meyerowitz, E.M. Redundancy and Specialization among Plant MicroRNAs: Role of the MIR164 Family in Developmental Robustness. Development 2007, 134, 1051–1060. [Google Scholar] [CrossRef]
- Zhou, G.K.; Kubo, M.; Zhong, R.; Demura, T.; Ye, Z.H. Overexpression of MiR165 Affects Apical Meristem Formation, Organ Polarity Establishment and Vascular Development in Arabidopsis. Plant Cell Physiol. 2007, 48, 391–404. [Google Scholar] [CrossRef]
- Merelo, P.; Ram, H.; Caggiano, M.P.; Ohno, C.; Ott, F.; Straub, D.; Graeff, M.; Cho, S.K.; Yang, S.W.; Wenkel, S.; et al. Regulation of MIR165/166 by Class II and Class III Homeodomain Leucine Zipper Proteins Establishes Leaf Polarity. Proc. Natl. Acad. Sci. USA 2016, 113, 11973–11978. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X. Argonautes Compete for MiR165/166 to Regulate Shoot Apical Meristem Development. Curr. Opin. Plant Biol. 2012, 15, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Wang, H. The Role of HD-ZIP III Transcription Factors and MiR165/166 in Vascular Development and Secondary Cell Wall Formation. Plant Signal. Behav. 2015, 10, e1078955. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Sato, F.; Ohme-Takagi, M. Roles of MiR319 and TCP Transcription Factors in Leaf Development. Plant Physiol. 2017, 175, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of Leaf Morphogenesis by MicroRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Efroni, I.; Blum, E.; Goldshmidt, A.; Eshed, Y. A Protracted and Dynamic Maturation Schedule Underlies Arabidopsis Leaf Development. Plant Cell 2008, 20, 2293–2306. [Google Scholar] [CrossRef]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-Regulating Factors (GRFs): A Small Transcription Factor Family with Important Functions in Plant Biology. Mol. Plant 2015, 8, 998–1010. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Y.; Shi, W.; Yu, P.; Hu, Y.; Lv, J.; Fu, C.; Fan, M.; Bai, M.Y. The MiR396-GRFs Module Mediates the Prevention of Photo-Oxidative Damage by Brassinosteroids during Seedling de-Etiolation in Arabidopsis. Plant Cell 2020, 32, 2525–2542. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Mecchia, M.A.; Vercruyssen, L.; Smaczniak, C.; Kaufmann, K.; Inze, D.; Rodriguez, R.E.; Palatnik, J.F. Post-Transcriptional Control of GRF Transcription Factors by MicroRNA MiR396 and GIF Co-Activator Affects Leaf Size and Longevity. Plant J. 2014, 79, 413–426. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Lin, X.; Hong, X.; Zhu, Y.; Li, W.; He, W.; An, F.; Guo, H. Adenine Phosphoribosyl Transferase 1 Is a Key Enzyme Catalyzing Cytokinin Conversion from Nucleobases to Nucleotides in Arabidopsis. Mol. Plant 2013, 6, 1661–1672. [Google Scholar] [CrossRef]
- Li, W.X.; Oono, Y.; Zhu, J.; He, X.J.; Wu, J.M.; Iida, K.; Lu, X.Y.; Cui, X.; Jin, H.; Zhu, J.K. The Arabidopsis NFYA5 Transcription Factor Is Regulated Transcriptionally and Posttranscriptionally to Promote Drought Resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef]
- Li, G.; Zhang, H.; Li, J.; Zhang, Z.; Li, Z. Genetic Control of Panicle Architecture in Rice. Crop. J. 2021, 9, 590–597. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, C.; Baulcombe, D.C.; Chen, Z.J. Maternal SiRNAs as Regulators of Parental Genome Imbalance and Gene Expression in Endosperm of Arabidopsis Seeds. Proc. Natl. Acad. Sci. USA 2012, 109, 5529–5534. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, S.; Jiao, W.; Wang, L.; Wang, L.; Ye, W.; Lu, J.; Hong, D.; You, S.; Cheng, Z.; et al. Both Maternally and Paternally Imprinted Genes Regulate Seed Development in Rice. New Phytol. 2017, 216, 373–387. [Google Scholar] [CrossRef]
- Khemka, N.; Singh Rajkumar, M.; Garg, R.; Jain, M. Genome-Wide Profiling of MiRNAs during Seed Development Reveals Their Functional Relevance in Seed Size/Weight Determination in Chickpea. Plant Direct 2021, 5, e00299. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Wang, Z.; Zhang, L.; Yao, J.; Hua, K.; Liu, X.; Shi, H.; Zhu, J.K. The Grain Yield Modulator MiR156 Regulates Seed Dormancy through the Gibberellin Pathway in Rice. Nat. Commun. 2019, 10, 3822. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Luo, X.; Sun, F.; Hu, J.; Zha, X.; Su, W.; Yang, J. Overexpressing LncRNA LAIR Increases Grain Yield and Regulates Neighbouring Gene Cluster Expression in Rice. Nat. Commun. 2018, 9, 3516. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Liu, Y.; Wang, W.; Wu, L.; Ma, J.; Zhang, S.; Wang, J.; Feng, F.; Yuan, H.; Huang, X. Identification and Functional Prediction of CircRNAs of Developing Seeds in High Oleic Acid Sunflower (Helianthus annuus L.). Acta Physiol. Plant 2023, 45, 13. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Liao, J.Y.; Li, Z.Y.; Yu, Y.; Zhang, J.P.; Li, Q.F.; Qu, L.H.; Shu, W.S.; Chen, Y.Q. Genome-Wide Screening and Functional Analysis Identify a Large Number of Long Noncoding RNAs Involved in the Sexual Reproduction of Rice. Genome Biol. 2014, 15, 512. [Google Scholar] [CrossRef]
- Cao, P.; Fan, W.; Li, P.; Hu, Y. Genome-Wide Profiling of Long Noncoding RNAs Involved in Wheat Spike Development. BMC Genom. 2021, 22, 493. [Google Scholar] [CrossRef]
- He, G.; Luo, X.; Tian, F.; Li, K.; Zhu, Z.; Su, W.; Qian, X.; Fu, Y.; Wang, X.; Sun, C.; et al. Haplotype Variation in Structure and Expression of a Gene Cluster Associated with a Quantitative Trait Locus for Improved Yield in Rice. Genome Res. 2006, 16, 618–626. [Google Scholar] [CrossRef]
- Yang, G.; Deng, P.; Guo, Q.; Shi, T.; Pan, W.; Cui, L.; Liu, X.; Nie, X. Population Transcriptomic Analysis Identifies the Comprehensive LncRNAs Landscape of Spike in Wheat (Triticum aestivum L.). BMC Plant Biol. 2022, 22, 450. [Google Scholar] [CrossRef]
- Sagun, J.V.; Yadav, U.P.; Alonso, A.P. Progress in Understanding and Improving Oil Content and Quality in Seeds. Front. Plant Sci. 2023, 14, 1116894. [Google Scholar] [CrossRef]
- Bhati, K.K.; Riyazuddin, R.; Pathak, A.; Singh, A. The survey of genetic engineering approaches for oil/fatty acid content improvement in oilseed crops. In Genome Engineering for Crop Improvement; Upadhyay, S.K., Ed.; Wiley: Hoboken, NJ, USA, 2021. [Google Scholar] [CrossRef]
- Shen, E.; Zhu, X.; Hua, S.; Chen, H.; Ye, C.; Zhou, L.; Liu, Q.; Zhu, Q.H.; Fan, L.; Chen, X. Genome-Wide Identification of Oil Biosynthesis-Related Long Non-Coding RNAs in Allopolyploid Brassica Napus 06 Biological Sciences 0604 Genetics 06 Biological Sciences 0607 Plant Biology. BMC Genom. 2018, 19, 745. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, A.; Zhao, Q.; Li, Z.; Lamboro, A.; He, H.; Li, Y.; Jiao, S.; Guan, S.; Liu, S.; et al. Genome-Wide Identification and Analysis of Long Non-Coding RNAs Involved in Fatty Acid Biosynthesis in Young Soybean Pods. Sci. Rep. 2021, 11, 7603. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, D.; Li, X.; Zhou, B.; Chang, T.; Hong, B.; Guan, C.; Guan, M. Integrated Analysis of LncRNA–MRNA Regulatory Networks Related to Lipid Metabolism in High-Oleic-Acid Rapeseed. Int. J. Mol. Sci. 2023, 24, 6277. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tan, Z.; Zeng, C.; Xiao, M.; Lin, S.; Yao, W.; Li, Q.; Guo, L.; Lu, S. Regulation of Seed Oil Accumulation by LncRNAs in Brassica Napus. Biotechnol. Biofuels Bioprod. 2023, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Mathan, J.; Yadav, A.; Goyal, A.K.; Chaudhury, A. Molecular and Transcriptional Regulation of Seed Development in Cereals: Present Status and Future Prospects; InTech Open: London, UK, 2021. [Google Scholar]
- Yoon, J.; Min, C.W.; Kim, J.; Baek, G.; Kim, D.; Jang, J.W.; Gupta, R.; Kim, S.T.; Cho, L.H. Quantitative Proteomic Analysis Deciphers the Molecular Mechanism for Endosperm Nuclear Division in Early Rice Seed Development. Plants 2023, 12, 3715. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Katoh, H.; Ogawa, D.; Kagaya, Y.; Sato, Y.; Kitano, H.; Nagato, Y.; Ishikawa, R.; Ono, A.; Kinoshita, T.; et al. Rice SNF2 Family Helicase ENL1 Is Essential for Syncytial Endosperm Development. Plant J. 2015, 81, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.F.; Zhang, Y.C.; Sun, Y.M.; Yu, Y.; Lei, M.Q.; Yang, Y.W.; Lian, J.P.; Feng, Y.Z.; Zhang, Z.; Yang, L.; et al. The Parent-of-Origin LncRNA MISSEN Regulates Rice Endosperm Development. Nat. Commun. 2021, 12, 6525. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Yadav, A.; Roy, J.K.; Bhati, K.K. Role of genome engineering for the development of resistant starch-rich, allergen-free and processing quality improved cereal crops. In Genome Engineering for Crop Improvement; Upadhyay, S.K., Ed.; Wiley: Hoboken, NJ, USA, 2021. [Google Scholar]
- Mishra, A.; Singh, A.; Sharma, M.; Kumar, P.; Roy, J. Development of EMS-Induced Mutation Population for Amylose and Resistant Starch Variation in Bread Wheat (Triticum Aestivum) and Identification of Candidate Genes Responsible for Amylose Variation. BMC Plant Biol. 2016, 16, 217. [Google Scholar] [CrossRef] [PubMed]
- Madhawan, A.; Sharma, A.; Bhandawat, A.; Rahim, M.S.; Kumar, P.; Mishra, A.; Parveen, A.; Sharma, H.; Verma, S.K.; Roy, J. Identification and Characterization of Long Non-Coding RNAs Regulating Resistant Starch Biosynthesis in Bread Wheat (Triticum aestivum L.). Genomics 2020, 112, 3065–3074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, R.; Meng, F.; Chen, Y.; Wang, W.; Yang, K.; Gao, Y.; Xin, M.; Du, J.; Hu, Z.; et al. A Comprehensive Atlas of Long Non-Coding RNAs Provides Insight into Grain Development in Wheat. Seed Biol. 2023, 2, 12. [Google Scholar] [CrossRef]
- Khalid, A.; Hameed, A.; Tahir, M.F. Wheat Quality: A Review on Chemical Composition, Nutritional Attributes, Grain Anatomy, Types, Classification, and Function of Seed Storage Proteins in Bread Making Quality. Front. Nutr. 2023, 10, 1053196. [Google Scholar] [CrossRef]
- Collin, A.; Daszkowska-Golec, A.; Szarejko, I. Updates on the Role of ABSCISIC ACID INSENSITIVE 5 (ABI5) and ABSCISIC ACID-RESPONSIVE ELEMENT BINDING FACTORS (ABFs) in ABA Signaling in Different Developmental Stages in Plants. Cells 2021, 10, 1996. [Google Scholar] [CrossRef]
- Gao, Q.; Liu, J.; Weng, H.; Yuan, X.; Xiao, W.; Wang, H. A Long Noncoding RNA Derived from LncRNA–MRNA Networks Modulates Seed Vigor. Int. J. Mol. Sci. 2022, 23, 9472. [Google Scholar] [CrossRef]
- Hazra, S.; Moulick, D.; Mukherjee, A.; Sahib, S.; Chowardhara, B.; Majumdar, A.; Upadhyay, M.K.; Yadav, P.; Roy, P.; Santra, S.C.; et al. Evaluation of Efficacy of Non-Coding RNA in Abiotic Stress Management of Field Crops: Current Status and Future Prospective. Plant Physiol. Biochem. 2023, 203, 107940. [Google Scholar] [CrossRef]
- Yang, H.; Cui, Y.; Feng, Y.; Hu, Y.; Liu, L.; Duan, L. Long Non-Coding RNAs of Plants in Response to Abiotic Stresses and Their Regulating Roles in Promoting Environmental Adaption. Cells 2023, 12, 729. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhao, F.; Zhou, B. The Characters of Non-Coding RNAs and Their Biological Roles in Plant Development and Abiotic Stress Response. Int. J. Mol. Sci. 2022, 23, 4124. [Google Scholar] [CrossRef] [PubMed]
- Gelaw, T.A.; Sanan-Mishra, N. Non-Coding RNAs in Response to Drought Stress. Int. J. Mol. Sci. 2021, 22, 12519. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.; Ahn, H.; Shin, S.J.; Kim, J.; Kwon, H.B.; Jung, W.; Kim, S. Clustering and evolutionary analysis of small RNAs identify regulatory siRNA clusters induced under drought stress in rice. BMC Syst. Biol. 2016, 23, 115. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Ni, Z.; Peng, H.; Sun, F.; Xin, M.; Sunkar, R.; Zhu, J.K.; Sun, Q. Non-coding small RNAs responsive to abiotic stress in wheat (Triticum aestivum L.). Funct. Integr. Genom. 2010, 10, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Ma, R.; Wang, C.; Chen, N.; Liu, S.; Qu, J.; Guan, S.; Ma, Y. Integration of MRNA and MicroRNA Analysis Reveals the Molecular Mechanisms Underlying Drought Stress Tolerance in Maize (Zea mays L.). Front. Plant Sci. 2022, 13, 932667. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Burd, S.; Lers, A. MiR408 Is Involved in Abiotic Stress Responses in Arabidopsis. Plant J. 2015, 84, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Balyan, S.; Kumar, M.; Mutum, R.D.; Raghuvanshi, U.; Agarwal, P.; Mathur, S.; Raghuvanshi, S. Identification of MiRNA-Mediated Drought Responsive Multi-Tiered Regulatory Network in Drought Tolerant Rice, Nagina. Sci. Rep. 2017, 7, 15446. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Liang, R.; Ge, L.; Li, W.; Xiao, H.; Lin, H.; Ruan, K.; Jin, Y. Identification of Drought-Induced MicroRNAs in Rice. Biochem. Biophys. Res. Commun. 2007, 354, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Hu, Z.; Jiang, Q.; Zhang, H. GmNFYA3, a Target Gene of MiR169, Is a Positive Regulator of Plant Tolerance to Drought Stress. Plant Mol. Biol. 2013, 82, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.J.; Jung, H.; Jeong, D.H.; Ha, S.H.; Choi, Y.D.; Kim, J.K. Transcriptome Profiling of Drought Responsive Noncoding RNAs and Their Target Genes in Rice. BMC Genom. 2016, 17, 563. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, S.R.; Danilevskaya, O.N.; Meng, X.; Beatty, M.; Zastrow-Hayes, G.; Harris, C.; Van Allen, B.; Habben, J.; Li, B. Genome-Wide Analysis of Alternative Splicing during Development and Drought Stress in Maize1[OPEN]. Plant Physiol. 2016, 170, 586–599. [Google Scholar] [CrossRef]
- Pang, J.; Zhang, X.; Ma, X.; Zhao, J. Spatio-Temporal Transcriptional Dynamics of Maize Long Non-Coding RNAs Responsive to Drought Stress. Genes 2019, 10, 138. [Google Scholar] [CrossRef]
- Qin, T.; Zhao, H.; Cui, P.; Albesher, N.; Xionga, L. A Nucleus-Localized Long Non-Coding Rna Enhances Drought and Salt Stress Tolerance. Plant Physiol. 2017, 175, 1321–1336. [Google Scholar] [CrossRef]
- Dong, S.-M.; Xiao, L.; Li, Z.-B.; Shen, J.; Yan, H.-B.; Li, S.-X.; Liao, W.-B.; Peng, M. A Novel Long Non-Coding RNA, DIR, Increases Drought Tolerance in Cassava by Modifying Stress-Related Gene Expression. J. Integr. Agric. 2022, 21, 2588–2602. [Google Scholar] [CrossRef]
- Zhang, P.; Fan, Y.; Sun, X.; Chen, L.; Terzaghi, W.; Bucher, E.; Li, L.; Dai, M. A Large-Scale Circular RNA Profiling Reveals Universal Molecular Mechanisms Responsive to Drought Stress in Maize and Arabidopsis. Plant J. 2019, 98, 697–713. [Google Scholar] [CrossRef]
- Ache, P.; Becker, D.; Ivashikina, N.; Dietrich, P.; Roelfsema, M.R.G.; Hedrich, R. GORK, a Delayed Outward Rectifier Expressed in Guard Cells of Arabidopsis Thaliana, Is a K+-Selective, K+-Sensing Ion Channel. FEBS Lett. 2000, 486, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Hoth, S.; Ache, P.; Wenkel, S.; Roelfsema, M.R.G.; Meyerhoff, O.; Hartung, W.; Hedrich, R. Regulation of the ABA-Sensitive Arabidopsis Potassium Channel Gene GORK in Response to Water Stress. FEBS Lett. 2003, 554, 119–126. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Plant Responses and Tolerance to Salt Stress: Physiological and Molecular Interventions. Int. J. Mol. Sci. 2022, 23, 15740. [Google Scholar] [CrossRef]
- Mirdar Mansuri, R.; Shobbar, Z.S.; Babaeian Jelodar, N.; Ghaffari, M.R.; Nematzadeh, G.A.; Asari, S. Dissecting Molecular Mechanisms Underlying Salt Tolerance in Rice: A Comparative Transcriptional Profiling of the Contrasting Genotypes. Rice 2019, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Wu, J.; Luo, Q.; Zhang, D.; Zhuang, W.; Xiao, G.; Deng, Q.; Bai, B. Effects of Salinity Stress at Reproductive Growth Stage on Rice (Oryza sativa L.) Composition, Starch Structure, and Physicochemical Properties. Front. Nutr. 2022, 9, 926217. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Agarwala, N.; Gill, S.S.; Varshney, R.K. Emerging Role of Plant Long Non Coding RNAs (LncRNAs) in Salinity Stress Response. Plant Stress 2023, 10, 100265. [Google Scholar] [CrossRef]
- Parmar, S.; Gharat, S.A.; Tagirasa, R.; Chandra, T.; Behera, L.; Dash, S.K.; Shaw, B.P. Identification and Expression Analysis of MiRNAs and Elucidation of Their Role in Salt Tolerance in Rice Varieties Susceptible and Tolerant to Salinity. PLoS ONE 2020, 15, e0230958. [Google Scholar] [CrossRef]
- Wang, Z.; Li, N.; Yu, Q.; Wang, H. Genome-Wide Characterization of Salt-Responsive Mirnas, Circrnas and Associated Cerna Networks in Tomatoes. Int. J. Mol. Sci. 2021, 22, 2238. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Guo, H.; Li, X.; Tang, M.; Guo, C.; Bao, H.; Huang, L.; Yi, Y.; Cui, Y.; Chen, L. OsSIDP301, a Member of the DUF1644 Family, Negatively Regulates Salt Stress and Grain Size in Rice. Front. Plant Sci. 2022, 13, 863233. [Google Scholar] [CrossRef]
- Ren, Z.; Zheng, Z.; Chinnusamy, V.; Zhu, J.; Cui, X.; Iida, K.; Zhu, J.K. RAS1, a Quantitative Trait Locus for Salt Tolerance and ABA Sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 5669–5674. [Google Scholar] [CrossRef]
- Jiang, Y.; Peng, X.; Zhang, Q.; Liu, Y.; Li, A.; Cheng, B.; Wu, J. Regulation of Drought and Salt Tolerance by OsSKL2 and OsASR1 in Rice. Rice 2022, 15, 46. [Google Scholar] [CrossRef]
- Borsani, O.; Zhu, J.; Verslues, P.E.; Sunkar, R.; Zhu, J.K. Endogenous SiRNAs Derived from a Pair of Natural Cis-Antisense Transcripts Regulate Salt Tolerance in Arabidopsis. Cell 2005, 123, 1279–1291. [Google Scholar] [CrossRef]
- Wan, J.; Meng, S.; Wang, Q.; Zhao, J.; Qiu, X.; Wang, L.; Li, J.; Lin, Y.; Mu, L.; Dang, K.; et al. Suppression of MicroRNA168 Enhances Salt Tolerance in Rice (Oryza sativa L.). BMC Plant Biol. 2022, 22, 563. [Google Scholar] [CrossRef]
- Mirdar Mansuri, R.; Azizi, A.H.; Sadri, A.H.; Shobbar, Z.S. Long Non-Coding RNAs as the Regulatory Hubs in Rice Response to Salt Stress. Sci. Rep. 2022, 12, 21696. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Z.; Liu, M.; Zhao, M.G.; Chen, R.; Zhang, W.H. Identification and Characterization of Long Non-Coding RNAs Involved in Osmotic and Salt Stress in Medicago Truncatula Using Genome-Wide High-Throughput Sequencing. BMC Plant Biol. 2015, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, Y.; Liu, H.; Liang, Z.; Zhang, M.; Zou, C.; Yuan, G.; Gao, S.; Pan, G.; Shen, Y.; et al. A Combination of a Genome-Wide Association Study and a Transcriptome Analysis Reveals CircRNAs as New Regulators Involved in the Response to Salt Stress in Maize. Int. J. Mol. Sci. 2022, 23, 9755. [Google Scholar] [CrossRef]
- Yin, J.; Liu, Y.; Lu, L.; Zhang, J.; Chen, S.; Wang, B. Comparison of Tolerant and Susceptible Cultivars Revealed the Roles of Circular RNAs in Rice Responding to Salt Stress. Plant Growth. Regul. 2022, 96, 243–254. [Google Scholar] [CrossRef]
- Jain, P.; Hussian, S.; Nishad, J.; Dubey, H.; Bisht, D.S.; Sharma, T.R.; Mondal, T.K. Identification and Functional Prediction of Long Non-Coding RNAs of Rice (Oryza sativa L.) at Reproductive Stage under Salinity Stress. Mol. Biol. Rep. 2021, 48, 2261–2271. [Google Scholar] [CrossRef]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and Waterlogging Stress in Plants: A Review Highlighting Research Opportunities and Understudied Aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef]
- Aslam, A.; Mahmood, A.; Ur-Rehman, H.; Li, C.; Liang, X.; Shao, J.; Negm, S.; Moustafa, M.; Aamer, M.; Hassan, M.U. Plant Adaptation to Flooding Stress under Changing Climate Conditions: Ongoing Breakthroughs and Future Challenges. Plants 2023, 12, 3824. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, D.; Spriggs, A.; Yang, J.; Pogson, B.J.; Dennis, E.S.; Wilson, I.W. Hypoxia-Responsive MicroRNAs and Trans-Acting Small Interfering RNAs in Arabidopsis. J. Exp. Bot. 2010, 61, 165–177. [Google Scholar] [CrossRef]
- Zhai, L.; Liu, Z.; Zou, X.; Jiang, Y.; Qiu, F.; Zheng, Y.; Zhang, Z. Genome-Wide Identification and Analysis of MicroRNA Responding to Long-Term Waterlogging in Crown Roots of Maize Seedlings. Physiol. Plant. 2013, 147, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Tan, Z.; Fang, T.; Tang, K.; Liang, K.; Qiu, F. A Comprehensive Transcriptomics Analysis Reveals Long Non-Coding RNA to Be Involved in the Key Metabolic Pathway in Response to Waterlogging Stress in Maize. Genes 2020, 11, 267. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.K.; Rawal, H.C.; Jain, P.; Mishra, V.; Nishad, J.; Chowrasia, S.; Sarkar, A.K.; Sen, P.; Naik, S.K.; Mondal, T.K. Identification and Analysis of MiRNAs-LncRNAs-MRNAs Modules Involved in Stem-Elongation of Deepwater Rice (Oryza sativa L.). Physiol. Plant. 2022, 174, e13736. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Chen, L.; Zhang, Y.; Kang, X.; Zhang, Z.; Wang, Y. Identification of Novel and Conserved Populus Tomentosa MicroRNA as Components of a Response to Water Stress. Funct. Integr. Genom. 2012, 12, 327–339. [Google Scholar] [CrossRef]
- Feyissa, B.A.; Amyot, L.; Nasrollahi, V.; Papadopoulos, Y.; Kohalmi, S.E.; Hannoufa, A. Involvement of the MiR156/SPL Module in Flooding Response in Medicago sativa. Sci. Rep. 2021, 11, 3243. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Jiang, F.; Zheng, X.; Wu, Z. Identification and Analysis of Oxygen Responsive MicroRNAs in the Root of Wild Tomato (S. habrochaites). BMC Plant Biol. 2019, 19, 100. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, K.; Pan, J.; Chen, X. Small RNA Sequencing Identifies Cucumber MiRNA Roles in Waterlogging-Triggered Adventitious Root Primordia Formation. Mol. Biol. Rep. 2019, 46, 6381–6389. [Google Scholar] [CrossRef] [PubMed]
- Kęska, K.; Szcześniak, M.W.; Adamus, A.; Czernicka, M. Waterlogging-Stress-Responsive LncRNAs, Their Regulatory Relationships with MiRNAs and Target Genes in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2021, 22, 8197. [Google Scholar] [CrossRef]
- Cho, Y.H.; Hong, J.W.; Kim, E.C.; Yoo, S.D. Regulatory Functions of SnRK1 in Stress-Responsive Gene Expression and in Plant Growth and Development. Plant Physiol. 2012, 158, 1955–1964. [Google Scholar] [CrossRef]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.R.; Bäurle, I. Arabidopsis MiR156 Regulates Tolerance to Recurring Environmental Stress through SPL Transcription Factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef]
- Jeyakumar, J.M.J.; Ali, A.; Wang, W.M.; Thiruvengadam, M. Characterizing the Role of the MiR156-SPL Network in Plant Development and Stress Response. Plants 2020, 9, 1206. [Google Scholar] [CrossRef]
- Wang, H.; Lu, S.; Guan, X.; Jiang, Y.; Wang, B.; Hua, J.; Zou, B. Dehydration-Responsive Element Binding Protein 1C, 1E, and 1G Promote Stress Tolerance to Chilling, Heat, Drought, and Salt in Rice. Front. Plant Sci. 2022, 13, 851731. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, J.; Liu, Z.; Li, X.; Wu, F.; He, Y. HEAT-INDUCED TAS1 TARGET1 Mediates Thermotolerance via Heat Stress Transcription Factor A1a-Directed Pathways in Arabidopsis. Plant Cell 2014, 26, 1764–1780. [Google Scholar] [CrossRef]
- Pokhrel, S.; Meyers, B.C. Heat-Responsive MicroRNAs and Phased Small Interfering RNAs in Reproductive Development of Flax. Plant Direct 2022, 6, e385. [Google Scholar] [CrossRef] [PubMed]
- Kume, K.; Tsutsumi, K.I.; Saitoh, Y. TAS1 Trans-Acting SiRNA Targets Are Differentially Regulated at Low Temperature, and TAS1 Trans-Acting SiRNA Mediates Temperature-Controlled At1g51670 Expression. Biosci. Biotechnol. Biochem. 2010, 74, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, Y.; Zhang, H.; Wang, J.; Zinta, G.; Xie, S.; Zhu, W.; Nie, W.F. Genome-Wide Identification of Circular RNAs in Response to Low-Temperature Stress in Tomato Leaves. Front. Genet. 2020, 11, 591806. [Google Scholar] [CrossRef]
- Zuo, J.; Wang, Q.; Zhu, B.; Luo, Y.; Gao, L. Deciphering the Roles of CircRNAs on Chilling Injury in Tomato. Biochem. Biophys. Res. Commun. 2016, 479, 132–138. [Google Scholar] [CrossRef]
- Pan, T.; Sun, X.; Liu, Y.; Li, H.; Deng, G.; Lin, H.; Wang, S. Heat Stress Alters Genome-Wide Profiles of Circular RNAs in Arabidopsis. Plant Mol. Biol. 2018, 96, 217–229. [Google Scholar] [CrossRef]
- Mao, H.D.; Yu, L.J.; Li, Z.J.; Yan, Y.; Han, R.; Liu, H.; Ma, M. Genome-Wide Analysis of the SPL Family Transcription Factors and Their Responses to Abiotic Stresses in Maize. Plant Gene 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Arshad, M.; Hannoufa, A. Alfalfa Transcriptome Profiling Provides Insight into MiR156-Mediated Molecular Mechanisms of Heat Stress Tolerance. Genome 2022, 65, 315–330. [Google Scholar] [CrossRef]
- Zhou, M.; Tang, W. MicroRNA156 Amplifies Transcription Factor-Associated Cold Stress Tolerance in Plant Cells. Mol. Genet. Genom. 2019, 294, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, F.; Cao, H.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. TamiR159 Directed Wheat TaGAMYB Cleavage and Its Involvement in Anther Development and Heat Response. PLoS ONE 2012, 7, e48445. [Google Scholar] [CrossRef]
- Lin, J.S.; Kuo, C.C.; Yang, I.C.; Tsai, W.A.; Shen, Y.H.; Lin, C.C.; Liang, Y.C.; Li, Y.C.; Kuo, Y.W.; King, Y.C.; et al. MicroRNA160 Modulates Plant Development and Heat Shock Protein Gene Expression to Mediate Heat Tolerance in Arabidopsis. Front. Plant Sci. 2018, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.A.; Sung, P.H.; Kuo, Y.W.; Chen, M.C.; Jeng, S.T.; Lin, J.S. Involvement of MicroRNA164 in Responses to Heat Stress in Arabidopsis. Plant Sci. 2023, 329, 111598. [Google Scholar] [CrossRef]
- Li, J.; Cao, Y.; Zhang, J.; Zhu, C.; Tang, G.; Yan, J. The MiR165/166–PHABULOSA Module Promotes Thermotolerance by Transcriptionally and Posttranslationally Regulating HSFA1. Plant Cell 2023, 35, 2952–2971. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, D.; Li, H.; Chen, Q.; Zhang, Z.; Liu, M.; Liu, J.; Song, Y.; He, J.; Xu, W.; et al. Characterization and Identification of Grapevine Heat Stress-Responsive MicroRNAs Revealed the Positive Regulated Function of Vvi-MiR167 in Thermostability. Plant Sci. 2023, 329, 111623. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, C.; Cheng, X.; Zhao, X.; Li, T.; Jiang, J. MiR168 Targets Argonaute1A Mediated MiRNAs Regulation Pathways in Response to Potassium Deficiency Stress in Tomato. BMC Plant Biol. 2020, 20, 477. [Google Scholar] [CrossRef]
- Mahale, B.M.; Fakrudin, B.; Ghosh, S.; Krishnaraj, P.U. LNA Mediated in Situ Hybridization of MiR171 and MiR397a in Leaf and Ambient Root Tissues Revealed Expressional Homogeneity in Response to Shoot Heat Shock in Arabidopsis thaliana. J. Plant Biochem. Biotechnol. 2014, 23, 93–103. [Google Scholar] [CrossRef]
- Jung, J.H.; Seo, P.J.; Ahn, J.H.; Park, C.M. Arabidopsis RNA-Binding Protein FCA Regulates MicroRNA172 Processing in Thermosensory Flowering. J. Biol. Chem. 2012, 287, 16007–16016. [Google Scholar] [CrossRef]
- Fang, Y.; Zheng, Y.; Lu, W.; Li, J.; Duan, Y.; Zhang, S.; Wang, Y. Roles of MiR319-Regulated TCPs in Plant Development and Response to Abiotic Stress. Crop. J. 2021, 9, 17–28. [Google Scholar] [CrossRef]
- Thiebaut, F.; Rojas, C.A.; Almeida, K.L.; Grativol, C.; Domiciano, G.C.; Lamb, C.R.C.; De Almeida Engler, J.; Hemerly, A.S.; Ferreira, P.C.G. Regulation of MiR319 during Cold Stress in Sugarcane. Plant Cell Environ. 2012, 35, 502–512. [Google Scholar] [CrossRef]
- Wang, S.T.; Sun, X.L.; Hoshino, Y.; Yu, Y.; Jia, B.; Sun, Z.W.; Sun, M.Z.; Duan, X.B.; Zhu, Y.M. MicroRNA319 Positively Regulates Cold Tolerance by Targeting OsPCF6 and OsTCP21 in Rice (Oryza sativa L.). PLoS ONE 2014, 9, e91357. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, S.; Zhou, M.; Yuan, N.; Li, Z.; Hu, Q.; Bethea, F.G.; Liu, H.; Li, S.; Luo, H. Transgenic Creeping Bentgrass Overexpressing Osa-MiR393a Exhibits Altered Plant Development and Improved Multiple Stress Tolerance. Plant Biotechnol. J. 2019, 17, 233–251. [Google Scholar] [CrossRef]
- Ding, B.; Yue, Y.; Chen, X.; Long, X.; Zhou, Z. Identification and Expression Analysis of MiR396 and Its Target Genes in Jerusalem Artichoke under Temperature Stress. Gene 2024, 893, 147908. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kwak, K.J.; Jung, H.J.; Lee, H.J.; Kang, H. MicroRNA402 Affects Seed Germination of Arabidopsis Thaliana under Stress Conditions via Targeting DEMETER-LIKE Protein3 MRNA. Plant Cell Physiol. 2010, 51, 1079–1083. [Google Scholar] [CrossRef]
- He, J.; Jiang, Z.; Gao, L.; You, C.; Ma, X.; Wang, X.; Xu, X.; Mo, B.; Chen, X.; Liu, L. Genome-Wide Transcript and Small RNA Profiling Reveals Transcriptomic Responses to Heat Stress. Plant Physiol. 2019, 181, 609–629. [Google Scholar] [CrossRef]
- Yu, J.; Su, D.; Yang, D.; Dong, T.; Tang, Z.; Li, H.; Han, Y.; Li, Z.; Zhang, B. Chilling and Heat Stress-Induced Physiological Changes and MicroRNA-Related Mechanism in Sweetpotato (Ipomoea batatas L.). Front. Plant Sci. 2020, 11, 687. [Google Scholar] [CrossRef]
- Xin, M.; Wang, Y.; Yao, Y.; Song, N.; Hu, Z.; Qin, D.; Xie, C.; Peng, H.; Ni, Z.; Sun, Q. Identification and Characterization of Wheat Long Non-Protein Coding RNAs Responsive to Powdery Mildew Infection and Heat Stress by Using Microarray Analysis and SBS Sequencing. BMC Plant Biol. 2011, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Dai, L.; Ai, J.; Wang, Y.; Ren, F. Identification and Functional Prediction of Cold-Related Long Non-Coding RNA (LncRNA) in Grapevine. Sci. Rep. 2019, 9, 6638. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Yang, X.; Aslam, M.M.; Li, M.; Zhu, L.; Chen, S.; Zhu, G.; Li, Y.; Liu, P. Genome-Wide Identification of Long Non-Coding RNAs in Two Contrasting Rapeseed (Brassica Napus L.) Genotypes Subjected to Cold Stress. Environ. Exp. Bot. 2022, 201, 104969. [Google Scholar] [CrossRef]
- Wang, A.; Hu, J.; Gao, C.; Chen, G.; Wang, B.; Lin, C.; Song, L.; Ding, Y.; Zhou, G. Genome-Wide Analysis of Long Non-Coding RNAs Unveils the Regulatory Roles in the Heat Tolerance of Chinese Cabbage (Brassica Rapa Ssp.Chinensis). Sci. Rep. 2019, 9, 5002. [Google Scholar] [CrossRef]
- Song, Y.; Chen, P.; Liu, P.; Bu, C.; Zhang, D. High-Temperature-Responsive Poplar LncRNAs Modulate Target Gene Expression via RNA Interference and Act as RNA Scaffolds to Enhance Heat Tolerance. Int. J. Mol. Sci. 2020, 21, 808. [Google Scholar] [CrossRef]
- Di, C.; Yuan, J.; Wu, Y.; Li, J.; Lin, H.; Hu, L.; Zhang, T.; Qi, Y.; Gerstein, M.B.; Guo, Y.; et al. Characterization of Stress-Responsive LncRNAs in Arabidopsis Thaliana by Integrating Expression, Epigenetic and Structural Features. Plant J. 2014, 80, 848–861. [Google Scholar] [CrossRef]
- Wunderlich, M.; Groß-Hardt, R.; Schöffl, F. Heat Shock Factor HSFB2a Involved in Gametophyte Development of Arabidopsis Thaliana and Its Expression Is Controlled by a Heat-Inducible Long Non-Coding Antisense RNA. Plant Mol. Biol. 2014, 85, 541–550. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, S.; Li, W.; Wang, S.; Hao, L.; Xu, C.; Yu, Y.; Xiang, L.; Li, T.; Jiang, F. A Long Noncoding RNA HILinc1 Enhances Pear Thermotolerance by Stabilizing PbHILT1 Transcripts through Complementary Base Pairing. Commun. Biol. 2022, 5, 1134. [Google Scholar] [CrossRef] [PubMed]
- Jampala, P.; Garhewal, A.; Lodha, M. Functions of Long Non-Coding RNA in Arabidopsis thaliana. Plant Signal. Behav. 2021, 16, 1925440. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Nayyar, H.; Roychowdhury, R.; Prasad, P.V.V.; Parida, S.K.; Siddique, K.H.M. Non-Coding RNAs (NcRNAs) in Plant: Master Regulators for Adapting to Extreme Temperature Conditions. Plant Physiol. Biochem. 2023, 205, 108164. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, J.; Lian, B.; Gu, H.; Li, Y.; Qi, Y. Global Identification of Arabidopsis LncRNAs Reveals the Regulation of MAF4 by a Natural Antisense RNA. Nat. Commun. 2018, 9, 5056. [Google Scholar] [CrossRef]
- Arshad, M.; Feyissa, B.A.; Amyot, L.; Aung, B.; Hannoufa, A. MicroRNA156 Improves Drought Stress Tolerance in Alfalfa (Medicago Sativa) by Silencing SPL13. Plant Sci. 2017, 258, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Gruber, M.Y.; Wall, K.; Hannoufa, A. An Insight into MicroRNA156 Role in Salinity Stress Responses of Alfalfa. Front. Plant Sci. 2017, 8, 356. [Google Scholar] [CrossRef]
- Iglesias, M.J.; Terrile, M.C.; Windels, D.; Lombardo, M.C.; Bartoli, C.G.; Vazquez, F.; Estelle, M.; Casalongué, C.A. MiR393 Regulation of Auxin Signaling and Redox-Related Components during Acclimation to Salinity in Arabidopsis. PLoS ONE 2014, 9, e107678. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Suo, J.; Shi, B.; Zhou, C.; Bai, B.; Bian, H.; Zhu, M.; Han, N. The Barley MiR393 Has Multiple Roles in Regulation of Seedling Growth, Stomatal Density, and Drought Stress Tolerance. Plant Physiol. Biochem. 2019, 142, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Bo Song, J.; Gao, S.; Sun, D.; Li, H.; Xia Shu, X.; Min Yang, Z. MiR394 and LCR Are Involved in Arabidopsis Salt and Drought Stress Responses in an Abscisic Acid-Dependent Manner. BMC Plant Biol. 2013, 13, 210. [Google Scholar] [CrossRef] [PubMed]
- Song, J.B.; Gao, S.; Wang, Y.; Li, B.W.; Zhang, Y.L.; Yang, Z.M. MiR394 and Its Target Gene LCR Are Involved in Cold Stress Response in Arabidopsis. Plant Gene 2016, 5, 56–64. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, X.; Niu, F.; Sun, X.; Hu, Z.; Gao, F.; Zhang, H.; Jiang, Q. Overexpression of LncRNA77580 Regulates Drought and Salinity Stress Responses in Soybean. Plants 2023, 12, 181. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Zhao, T. High Throughput Sequencing of CircRNAs in Tomato Leaves Responding to Multiple Stresses of Drought and Heat. Hortic. Plant J. 2020, 6, 34–38. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, A.; Mathan, J.; Dubey, A.K.; Singh, A. The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling. Non-Coding RNA 2024, 10, 13. https://doi.org/10.3390/ncrna10010013
Yadav A, Mathan J, Dubey AK, Singh A. The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling. Non-Coding RNA. 2024; 10(1):13. https://doi.org/10.3390/ncrna10010013
Chicago/Turabian StyleYadav, Amit, Jyotirmaya Mathan, Arvind Kumar Dubey, and Anuradha Singh. 2024. "The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling" Non-Coding RNA 10, no. 1: 13. https://doi.org/10.3390/ncrna10010013
APA StyleYadav, A., Mathan, J., Dubey, A. K., & Singh, A. (2024). The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling. Non-Coding RNA, 10(1), 13. https://doi.org/10.3390/ncrna10010013