Breast Cancer and miR-SNPs: The Importance of miR Germ-Line Genetics


Abstract
1. Introduction
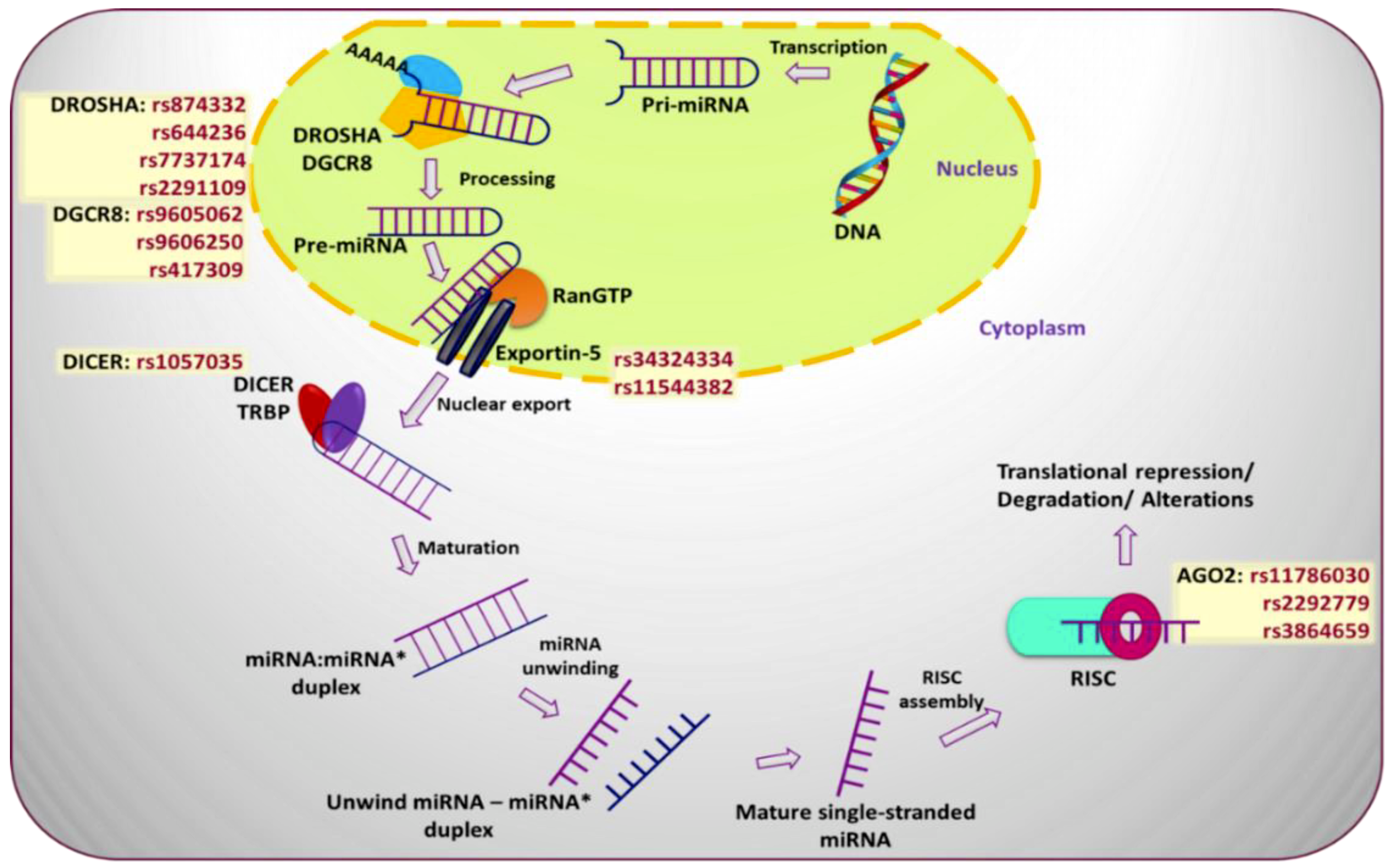
2. SNPs in miRNA Biogenesis Genes and Breast Cancer
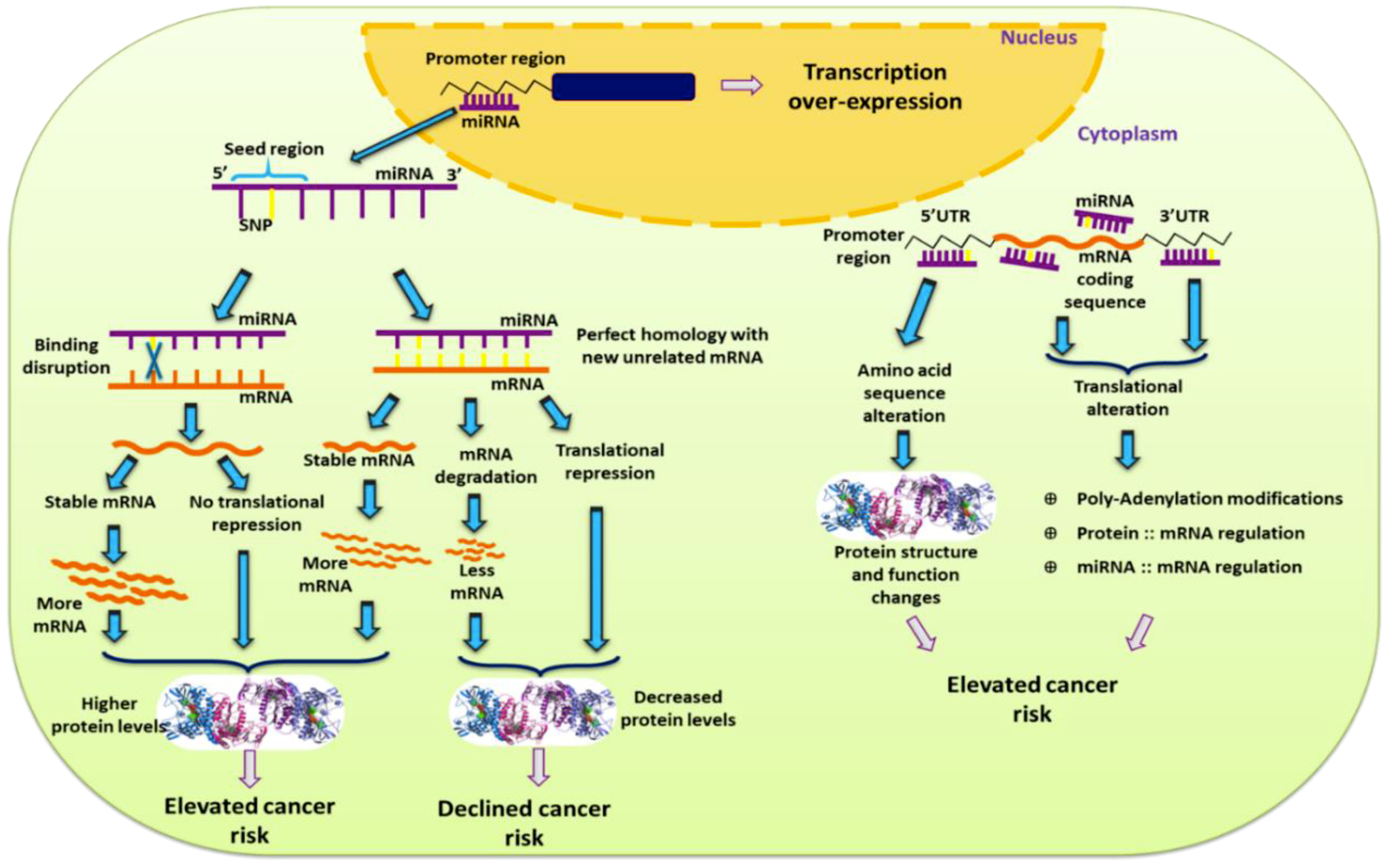
3. 3’UTR SNPs and Breast Cancer
4. The KRAS-Variant and Breast Cancer Risk
5. miRNA-Coding SNPs
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- US Breast Cancer Statistics. Available online: https://www.breastcancer.org/symptoms/understand_bc/statistics (accessed on 14 January 2019).
- How Common is Breast Cancer? Available online: https://www.cancer.org/cancer/breast-cancer/about/how-common-is-breast-cancer.html (accessed on 14 January 2019).
- Vogel, V. Approaches to Understanding Breast Cancer. In The Breast; Bland, K.K.VS., Copeland, E.M., Gradishar, W.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 207–218. [Google Scholar]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Frixa, T.; Donzelli, S.; Blandino, G. Oncogenic MicroRNAs: Key Players in Malignant Transformation. Cancers 2015, 7, 2466–2485. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Dien, G.T.; Smith, R.A.; Haupt, L.M.; Griffiths, L.R.; Nguyen, H.T. Genetic polymorphisms in miRNAs targeting the estrogen receptor and their effect on breast cancer risk. Meta Gene 2014, 2, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Gordon, O. Emery and Rimoin’s Principles and Practice of Medical Genetics; Rimoin, D.P.R., Korf, B., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 1–31. [Google Scholar]
- Murata, M.; Thompson, P.E. Two-nucleotide codon change in a hemoglobin polymorphism of the Celebes black ape (Macaca nigra). Biochem. Genet. 1976, 14, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [PubMed]
- Salzman, D.W.; Weidhaas, J.B. SNPing cancer in the bud: microRNA and microRNA-target site polymorphisms as diagnostic and prognostic biomarkers in cancer. Pharmacol. Ther. 2013, 137, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Weidhaas, J.B. Functional microRNA binding site variants. Mol. Oncol. 2019, 13, 4–8. [Google Scholar]
- Zheng, H.; Song, F.; Zhang, L.; Yang, D.; Ji, P.; Wang, Y.; Almeida, M.; Calin, G.A.; Hao, X.; Wei, Q.; et al. Genetic variants at the miR-124 binding site on the cytoskeleton-organizing IQGAP1 gene confer differential predisposition to breast cancer. Int. J. Oncol. 2011, 38, 1153–1161. [Google Scholar] [CrossRef]
- Chen, K.; Song, F.; Calin, G.A.; Wei, Q.; Hao, X.; Zhang, W. Polymorphisms in microRNA targets: A gold mine for molecular epidemiology. Carcinogenesis 2008, 29, 1306–1311. [Google Scholar] [CrossRef]
- Nicoloso, M.S.; Sun, H.; Spizzo, R.; Kim, H.; Wickramasinghe, P.; Shimizu, M.; Wojcik, S.E.; Ferdin, J.; Kunej, T.; Xiao, L.; et al. Single-nucleotide polymorphisms inside microRNA target sites influence tumor susceptibility. Cancer Res. 2010, 70, 2789–2798. [Google Scholar] [CrossRef]
- Ryan, B.M.; Robles, A.I.; Harris, C.C. Genetic variation in microRNA networks: The implications for cancer research. Nat. Rev. Cancer 2010, 10, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhong, H.; Wu, Q.; Wang, M.; Zhou, J.; Zhou, Y.; Lu, X.; Ying, B. Association between SNPs in microRNA machinery genes and gastric cancer susceptibility, invasion, and metastasis in Chinese Han population. Oncotarget 2017, 8, 86435–86446. [Google Scholar] [CrossRef]
- Guo, Z.; Shu, Y.; Zhou, H.; Zhang, W. Identification of diagnostic and prognostic biomarkers for cancer: Focusing on genetic variations in microRNA regulatory pathways (Review). Mol. Med. Rep. 2016, 13, 1943–1952. [Google Scholar] [CrossRef] [PubMed]
- Ryan, B. Advances in Cancer Research; Croce, C.F.P.B., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 151–171. [Google Scholar]
- Hoffman, A.E.; Zheng, T.; Yi, C.; Leaderer, D.; Weidhaas, J.; Slack, F.; Zhang, Y.; Paranjape, T.; Zhu, Y. microRNA miR-196a-2 and breast cancer: A genetic and epigenetic association study and functional analysis. Cancer Res. 2009, 69, 5970–5977. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Lee, K.M.; Choi, J.Y.; Han, S.; Lee, J.Y.; Li, L.; Park, S.K.; Yoo, K.Y.; Noh, D.Y.; Ahn, S.H.; et al. Common genetic polymorphisms of microRNA biogenesis pathway genes and risk of breast cancer: A case-control study in Korea. Breast Cancer Res. Treat. 2011, 130, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Jeon, S.; Lee, K.M.; Han, S.; Song, M.; Choi, J.Y.; Park, S.K.; Yoo, K.Y.; Noh, D.Y.; Ahn, S.H.; et al. Common genetic polymorphisms of microRNA biogenesis pathway genes and breast cancer survival. BMC Cancer 2012, 12, 195. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Kroliczewski, J.; Sobolewska, A.; Lejnowski, D.; Collawn, J.F.; Bartoszewski, R. microRNA single polynucleotide polymorphism influences on microRNA biogenesis and mRNA target specificity. Gene 2018, 640, 66–72. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, J.; Wu, J.; Hu, Z.; Qin, Z.; Liu, X.; Guan, X.; Wang, Y.; Han, J.; Jiang, T.; et al. Evaluation of genetic variants in microRNA biosynthesis genes and risk of breast cancer in Chinese women. Int. J. Cancer 2013, 133, 2216–2224. [Google Scholar] [CrossRef]
- Leaderer, D.; Hoffman, A.E.; Zheng, T.; Fu, A.; Weidhaas, J.; Paranjape, T.; Zhu, Y. Genetic and epigenetic association studies suggest a role of microRNA biogenesis gene exportin-5 (XPO5) in breast tumorigenesis. Int. J. Mol. Epidemiol. Genet. 2011, 2, 9–18. [Google Scholar]
- Ma, H.; Yuan, H.; Yuan, Z.; Yu, C.; Wang, R.; Jiang, Y.; Hu, Z.; Shen, H.; Chen, N. Genetic Variations in Key MicroRNA Processing Genes and Risk of Head and Neck Cancer: A Case-Control Study in Chinese Population. PLoS ONE 2012, 7, e47544. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.Y.; Kuang, D.; Yin, X.X. Association between the DICER rs1057035 polymorphism and cancer risk: Evidence from a meta-analysis of 12,675 individuals. Asian Pac. J. Cancer Prev. APJCP 2015, 16, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Luo, C.; Peng, R.; Guo, Q.; Wang, K.; Wang, P.; Ye, H.; Song, C. MiRNA-binding site functional polymorphisms in DNA repair genes RAD51, RAD52, and XRCC2 and breast cancer risk in Chinese population. Tumour Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qin, Z.; Hu, Z.; Guan, X.; Wang, Y.; He, Y.; Xue, J.; Liu, X.; Chen, J.; Dai, J.; et al. Genetic variation in a hsa-let-7 binding site in RAD52 is associated with breast cancer susceptibility. Carcinogenesis 2013, 34, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Dorairaj, J.J.; Salzman, D.W.; Wall, D.; Rounds, T.; Preskill, C.; Sullivan, C.A.; Lindner, R.; Curran, C.; Lezon-Geyda, K.; McVeigh, T.; et al. A germline mutation in the BRCA1 3′UTR predicts Stage IV breast cancer. BMC Cancer 2014, 14, 421. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, C.; Speed, W.C.; Paranjape, T.; Keane, K.; Blitzblau, R.; Hollestelle, A.; Safavi, K.; van den Ouweland, A.; Zelterman, D.; Slack, F.J.; et al. Rare BRCA1 haplotypes including 3′UTR SNPs associated with breast cancer risk. Cell Cycle 2011, 10, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Pongsavee, M.; Yamkamon, V.; Dakeng, S.; P, O.c.; Smith, D.R.; Saunders, G.F.; Patmasiriwat, P. The BRCA1 3′-UTR: 5711+421T/T_5711+1286T/T genotype is a possible breast and ovarian cancer risk factor. Genet. Test. Mol. Biomark. 2009, 13, 307–317. [Google Scholar] [CrossRef]
- Newman, B.; Mu, H.; Butler, L.M.; Millikan, R.C.; Moorman, P.G.; King, M.C. Frequency of breast cancer attributable to BRCA1 in a population-based series of American women. Jama 1998, 279, 915–921. [Google Scholar] [CrossRef]
- Yang, F.; Chen, F.; Xu, J.; Guan, X. Identification and frequency of the rs12516 and rs8176318 BRCA1 gene polymorphisms among different populations. Oncol. Lett. 2016, 11, 2481–2486. [Google Scholar] [CrossRef]
- Erturk, E.; Cecener, G.; Polatkan, V.; Gokgoz, S.; Egeli, U.; Tunca, B.; Tezcan, G.; Demirdogen, E.; Ak, S.; Tasdelen, I. Evaluation of genetic variations in miRNA-binding sites of BRCA1 and BRCA2 genes as risk factors for the development of early-onset and/or familial breast cancer. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 8319–8324. [Google Scholar] [CrossRef]
- Brendle, A.; Lei, H.; Brandt, A.; Johansson, R.; Enquist, K.; Henriksson, R.; Hemminki, K.; Lenner, P.; Försti, A. Polymorphisms in predicted microRNA-binding sites in integrin genes and breast cancer: ITGB4 as prognostic marker. Carcinogenesis 2008, 29, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Tchatchou, S.; Jung, A.; Hemminki, K.; Sutter, C.; Wappenschmidt, B.; Bugert, P.; Weber, B.H.; Niederacher, D.; Arnold, N.; Varon-Mateeva, R.; et al. A variant affecting a putative miRNA target site in estrogen receptor (ESR) 1 is associated with breast cancer risk in premenopausal women. Carcinogenesis 2009, 30, 59–64. [Google Scholar] [CrossRef]
- Fishman, J.; Osborne, M.P.; Telang, N.T. The role of estrogen in mammary carcinogenesis. Ann. N. Y. Acad. Sci. 1995, 768, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.; Davio, C.; Rivera, E.; Melito, G.; Cricco, G.; Andrade, N.; Caro, R.; Bergoc, R. Hormone dependence of mammary tumors induced in rats by intraperitoneal NMU injection. Cancer Investig. 1997, 15, 8–17. [Google Scholar] [CrossRef]
- Forma, E.; Brzeziańska, E.; Krześlak, A.; Chwatko, G.; Jóźwiak, P.; Szymczyk, A.; Smolarz, B.; Romanowicz-Makowska, H.; Różański, W.; Bryś, M. Association between the c.*229C>T polymorphism of the topoisomerase IIβ binding protein 1 (TopBP1) gene and breast cancer. Mol. Biol. Rep. 2013, 40, 3493–3502. [Google Scholar] [CrossRef] [PubMed]
- Forma, E. TopBP1 in DNA Damage Response. In DNA Repair; Kruman, I., Ed.; INTECH Open Access Publisher: Rijeka, Croatia, 2011; Volume 4, pp. 281–304. [Google Scholar]
- Rolli, M.; Fransvea, E.; Pilch, J.; Saven, A.; Felding-Habermann, B. Activated integrin alphavbeta3 cooperates with metalloproteinase MMP-9 in regulating migration of metastatic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 9482–9487. [Google Scholar] [CrossRef]
- Ren, F.; Tang, R.; Zhang, X.; Madushi, W.M.; Luo, D.; Dang, Y.; Li, Z.; Wei, K.; Chen, G. Overexpression of MMP Family Members Functions as Prognostic Biomarker for Breast Cancer Patients: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0135544. [Google Scholar] [CrossRef]
- Pirooz, H.J.; Jafari, N.; Rastegari, M.; Fathi-Roudsari, M.; Tasharrofi, N.; Shokri, G.; Tamadon, M.; Sazegar, H.; Kouhkan, F. Functional SNP in microRNA-491-5p binding site of MMP9 3′-UTR affects cancer susceptibility. J. Cell. Biochem. 2018, 119, 5126–5134. [Google Scholar] [CrossRef] [PubMed]
- Paranjape, T.; Heneghan, H.; Lindner, R.; Keane, F.K.; Hoffman, A.; Hollestelle, A.; Dorairaj, J.; Geyda, K.; Pelletier, C.; Nallur, S.; et al. A 3′-untranslated region KRAS variant and triple-negative breast cancer: A case-control and genetic analysis. Lancet Oncol. 2011, 12, 377–386. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. An epigenetic switch involving NF-kappaB, Lin28, Let-7 MicroRNA, and IL6 links inflammation to cell transformation. Cell 2009, 139, 693–706. [Google Scholar] [CrossRef]
- Sanaei, S.; Hashemi, M.; Eskandari, E.; Hashemi, S.M.; Bahari, G. KRAS Gene Polymorphisms and their Impact on Breast Cancer Risk in an Iranian Population. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 1301–1305. [Google Scholar]
- McVeigh, T.P.; Jung, S.Y.; Kerin, M.J.; Salzman, D.W.; Nallur, S.; Nemec, A.A.; Dookwah, M.; Sadofsky, J.; Paranjape, T.; Kelly, O.; et al. Estrogen withdrawal, increased breast cancer risk and the KRAS-variant. Cell Cycle 2015, 14, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Cerne, J.Z.; Stegel, V.; Gersak, K.; Novakovic, S. KRAS rs61764370 is associated with HER2-overexpressed and poorly-differentiated breast cancer in hormone replacement therapy users: A case control study. BMC Cancer 2012, 12, 105. [Google Scholar] [CrossRef]
- Uvirova, M.; Simova, J.; Kubova, B.; Dvorackova, N.; Tomaskova, H.; Sedivcova, M.; Dite, P. Comparison of the prevalence of KRAS-LCS6 polymorphism (rs61764370) within different tumour types (colorectal, breast, non-small cell lung cancer and brain tumours). A study of the Czech population. Biomed. Pap. 2015, 159, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Hollestelle, A.; van der Baan, F.H.; Berchuck, A.; Johnatty, S.E.; Aben, K.K.; Agnarsson, B.A.; Aittomaki, K.; Alducci, E.; Andrulis, I.L.; Anton-Culver, H.; et al. No clinical utility of KRAS variant rs61764370 for ovarian or breast cancer. Gynecol. Oncol. 2016, 141, 386–401. [Google Scholar] [CrossRef]
- Ustinova, M.; Daneberga, Z.; Berzina, D.; Nakazawa-Miklasevica, M.; Maksimenko, J.; Gardovskis, J.; Miklasevics, E. Impact of KRAS variant rs61764370 on breast cancer morbidity. Exp. Oncol. 2015, 37, 292–294. [Google Scholar] [CrossRef]
- Duan, R.; Pak, C.; Jin, P. Single nucleotide polymorphism associated with mature miR-125a alters the processing of pri-miRNA. Hum. Mol. Genet. 2007, 16, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Schlehe, B.; Hemminki, K.; Sutter, C.; Bugert, P.; Wappenschmidt, B.; Volkmann, J.; Varon, R.; Weber, B.H.; Niederacher, D.; et al. A genetic variant in the pre-miR-27a oncogene is associated with a reduced familial breast cancer risk. Breast Cancer Res. Treat. 2010, 121, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Fang, W.; Wu, X.; Bian, S.; Chen, G.; Lu, L.; Weng, Y. Distinct effects of rs895819 on risk of different cancers: An update meta-analysis. Oncotarget 2017, 8, 75336–75349. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Duan, F.; Song, C.; Zhao, X.; Dai, L.; Cui, S. Systematic evaluation of cancer risk associated with rs2292832 in miR-149 and rs895819 in miR-27a: A comprehensive and updated meta-analysis. Oncotarget 2016, 7, 22368–22384. [Google Scholar] [CrossRef]
- Ferracin, M.; Querzoli, P.; Calin, G.A.; Negrini, M. MicroRNAs: Toward the clinic for breast cancer patients. Semin. Oncol. 2011, 38, 764–775. [Google Scholar] [CrossRef]
- Zhang, N.; Huo, Q.; Wang, X.; Chen, X.; Long, L.; Jiang, L.; Ma, T.; Yang, Q. A genetic variant in pre-miR-27a is associated with a reduced breast cancer risk in younger Chinese population. Gene 2013, 529, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Wang, L.; Zhou, B.; Yao, W.J.; Xu, S.; Zhou, Y.; Xie, Z.B. Associations of miRNA polymorphisms and expression levels with breast cancer risk in the Chinese population. Genet. Mol. Res. GMR 2015, 14, 6289–6296. [Google Scholar] [CrossRef] [PubMed]
- Yekta, S.; Shih, I.H.; Bartel, D.P. MicroRNA-directed cleavage of HOXB8 mRNA. Science 2004, 304, 594–596. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.M.; Kang, H.F.; Zhang, W.G.; Li, H.B.; Zhang, S.Q.; Ma, X.B.; Lin, S.; Wang, M.; Feng, Y.J.; Liu, K.; et al. The Associations of Single Nucleotide Polymorphisms in miR196a2, miR-499, and miR-608 With Breast Cancer Susceptibility: A STROBE-Compliant Observational Study. Medicine 2016, 95, e2826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Y.; Yan, W.; Wang, W.; Zhao, X.; Ma, X.; Gao, X.; Zhang, S. Association between three functional microRNA polymorphisms (miR-499 rs3746444, miR-196a rs11614913 and miR-146a rs2910164) and breast cancer risk: A meta-analysis. Oncotarget 2016, 8, 393–407. [Google Scholar] [CrossRef]
- Wang, P.Y.; Gao, Z.H.; Jiang, Z.H.; Li, X.X.; Jiang, B.F.; Xie, S.Y. The associations of single nucleotide polymorphisms in miR-146a, miR-196a and miR-499 with breast cancer susceptibility. PLoS ONE 2013, 8, e70656. [Google Scholar] [CrossRef]
- Hu, Z.; Liang, J.; Wang, Z.; Tian, T.; Zhou, X.; Chen, J.; Miao, R.; Wang, Y.; Wang, X.; Shen, H. Common genetic variants in pre-microRNAs were associated with increased risk of breast cancer in Chinese women. Hum. Mutat. 2009, 30, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, W.; Jiang, W.; Lin, J.; Jiang, Y.; Li, B.; Pang, D. A miRNA binding site single-nucleotide polymorphism in the 3′-UTR region of the IL23R gene is associated with breast cancer. PLoS ONE 2012, 7, e49823. [Google Scholar] [CrossRef]
- Chen, Q.H.; Wang, Q.B.; Zhang, B. Ethnicity modifies the association between functional microRNA polymorphisms and breast cancer risk: A HuGE meta-analysis. Tumour Biol. 2014, 35, 529–543. [Google Scholar] [CrossRef]
- Catucci, I.; Verderio, P.; Pizzamiglio, S.; Bernard, L.; Dall’olio, V.; Sardella, D.; Ravagnani, F.; Galastri, L.; Barile, M.; Peissel, B.; et al. The SNP rs895819 in miR-27a is not associated with familial breast cancer risk in Italians. Breast Cancer Res. Treat. 2012, 133, 805–807. [Google Scholar] [CrossRef]
- Crowley, E.H.; Arena, S.; Lamba, S.; Di Nicolantonio, F.; Bardelli, A. Targeted knock-in of the polymorphism rs61764370 does not affect KRAS expression but reduces let-7 levels. Hum. Mutat. 2014, 35, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Pilarski, R.; Patel, D.A.; Weitzel, J.; McVeigh, T.; Dorairaj, J.J.; Heneghan, H.M.; Miller, N.; Weidhaas, J.B.; Kerin, M.J.; McKenna, M.; et al. The KRAS-variant is associated with risk of developing double primary breast and ovarian cancer. PLoS ONE 2012, 7, e37891. [Google Scholar] [CrossRef] [PubMed]
- Hollestelle, A.; Pelletier, C.; Hooning, M.; Crepin, E.; Schutte, M.; Look, M.; Collee, J.M.; Nieuwlaat, A.; Dorssers, L.C.; Seynaeve, C.; et al. Prevalence of the variant allele rs61764370 T>G in the 3′UTR of KRAS among Dutch BRCA1, BRCA2 and non-BRCA1/BRCA2 breast cancer families. Breast Cancer Res. Treat. 2011, 128, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Y.; Chien, Y.C.; Wong, Y.K.; Lin, Y.L.; Lin, J.C. Effects of KRAS mutation and polymorphism on the risk and prognosis of oral squamous cell carcinoma. Head Neck 2012, 34, 663–666. [Google Scholar] [CrossRef]
- Paranjape, T.; Slack, F.J.; Weidhaas, J.B. MicroRNAs: Tools for cancer diagnostics. Gut 2009, 58, 1546–1554. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Su, X.; Xing, J.; Wang, Z.; Chen, L.; Cui, M.; Jiang, B. microRNAs and ceRNAs: RNA networks in pathogenesis of cancer. Chinese J. Cancer Res. 2013, 25, 235–239. [Google Scholar]
- Chen, J.; Xu, J.; Li, Y.; Zhang, J.; Chen, H.; Lu, J.; Wang, Z.; Zhao, X.; Xu, K.; Li, Y.; et al. Competing endogenous RNA network analysis identifies critical genes among the different breast cancer subtypes. Oncotarget 2016, 8, 10171–10184. [Google Scholar] [CrossRef] [PubMed]
- Abdollahzadeh, R.; Daraei, A.; Mansoori, Y.; Sepahvand, M.; Amoli, M.M.; Tavakkoly-Bazzaz, J. Competing endogenous RNA (ceRNA) cross talk and language in ceRNA regulatory networks: A new look at hallmarks of breast cancer. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| SNP | Affected miRNA | Related Gene | Breast Cancer Association | References |
|---|---|---|---|---|
| rs7963551 (C allele) | let-7 | RAD52 | Reduced risk | [28,29] |
| rs8176318 (G > T) | miR-639 | BRCA1 | Elevated TNBC risk | [30,34] |
| rs12516 | miR-1264 | BRCA1 | Elevated risk | [32,35] |
| rs3092995 | 3′UTR | BRCA1 | Elevated risk | [33] |
| SNP | Affected miRNA | Related Gene | Breast Cancer Association | References |
|---|---|---|---|---|
| rs743554 | miR-34a | ITGB4 | Elevated risk | [36] |
| rs1042538 (A > T) | miR-124 | IQGAP1 | Reduced risk | [12] |
| rs2747648 | miR-453 | ESR1 | Elevated risk | [37] |
| rs334348 | miR-628-5p | TGFBR1 | Elevated risk | [14] |
| rs1982073-TGFB1 | miR-187 | TGFB1 | Elevated risk | [14] |
| rs1056628 | miR-491-5p | MMP9 | Elevated risk | [44,45] |
| SNP | Affected miRNA | Related Gene | Breast Cancer Association | References |
|---|---|---|---|---|
| rs895819 (A > G) | pre-miRNA-27a | MIR27a | Reduced risk | [54,55,57,58,67] |
| rs61764370 | 3′UTR of let-7 | KRAS | Elevated risk | [45,47,48,68,69,70] |
| rs11614913 | pre-miR-196a2 | MIR196a2 | Elevated risk | [57,59,63] |
| rs3746444 | miR-499 | MIR499 | Elevated risk | [64,66,71] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malhotra, P.; Read, G.H.; Weidhaas, J.B. Breast Cancer and miR-SNPs: The Importance of miR Germ-Line Genetics. Non-Coding RNA 2019, 5, 27. https://doi.org/10.3390/ncrna5010027
Malhotra P, Read GH, Weidhaas JB. Breast Cancer and miR-SNPs: The Importance of miR Germ-Line Genetics. Non-Coding RNA. 2019; 5(1):27. https://doi.org/10.3390/ncrna5010027
Chicago/Turabian StyleMalhotra, Poonam, Graham H. Read, and Joanne B. Weidhaas. 2019. "Breast Cancer and miR-SNPs: The Importance of miR Germ-Line Genetics" Non-Coding RNA 5, no. 1: 27. https://doi.org/10.3390/ncrna5010027
APA StyleMalhotra, P., Read, G. H., & Weidhaas, J. B. (2019). Breast Cancer and miR-SNPs: The Importance of miR Germ-Line Genetics. Non-Coding RNA, 5(1), 27. https://doi.org/10.3390/ncrna5010027