The Arabidopsis Hypoxia Inducible AtR8 Long Non-Coding RNA also Contributes to Plant Defense and Root Elongation Coordinating with WRKY Genes under Low Levels of Salicylic Acid

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Correlation between Transcription of AtR8 lncRNA and WRKY TF Genes in Arabidopsis Seedlings under Hypoxic Stress
2.2. Impact of SA Exposure in atr8 and wrky Mutants on Root Elongation
2.3. Long-Term SA Treatment and Gene Responses
2.4. Effects of Short-Term SA Treatment on Gene Responses
2.5. Rapid Induction of WRKY and AtR8 Gene Expression by Pseudomonas syringae
2.6. Temporal Accumulation of RNAs after Germination
2.7. Regulation of NPR1-Mediated Genes
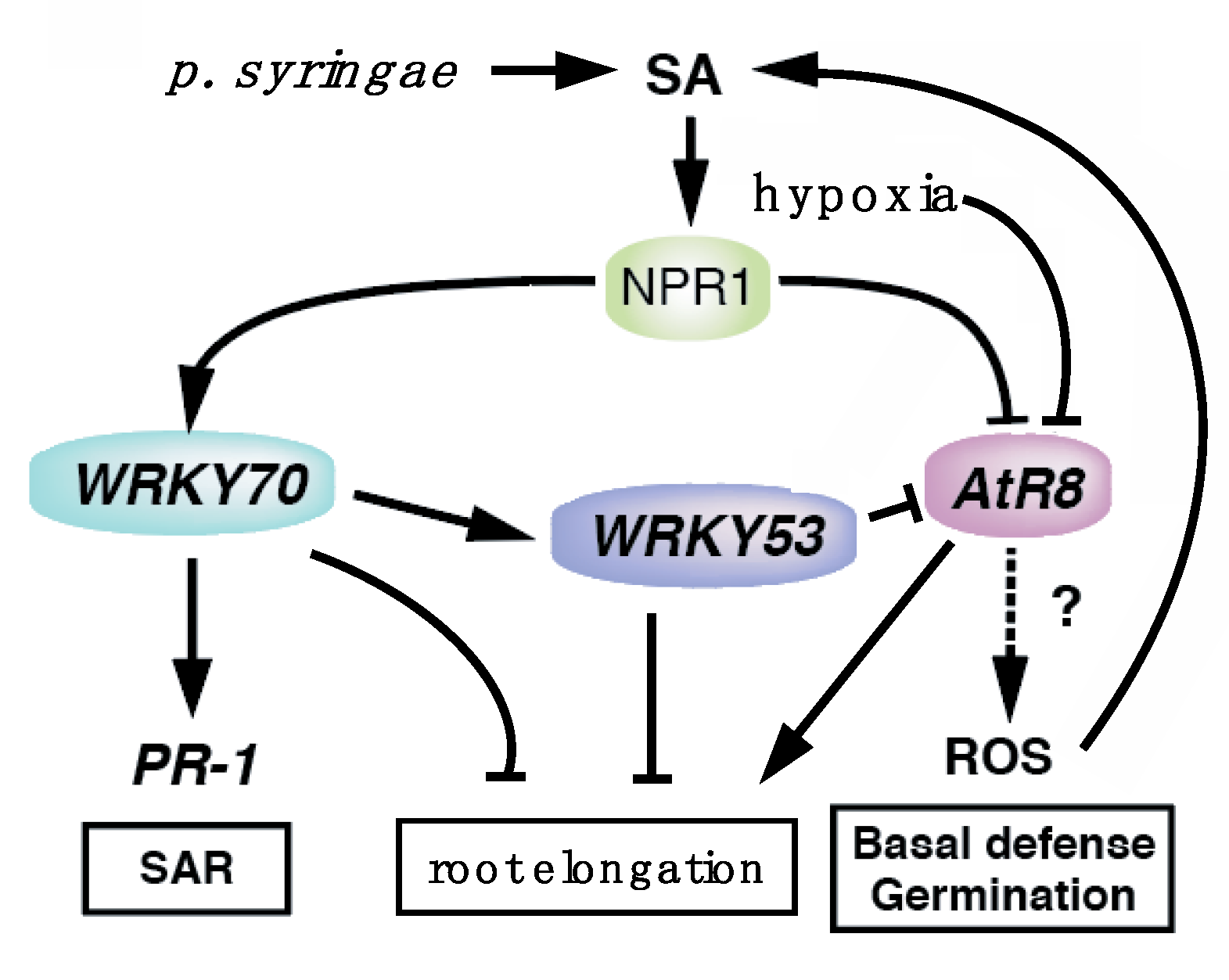
3. Discussion
3.1. Involvement of AtR8 lncRNA in Plant Defense
3.2. Delayed Root Elongation in atr8 Strongly Associates with SA
3.3. Concentration-Dependent SA Effects
3.4. How are AtR8 and WRKY70 Expression Regulated by SA?
3.5. Biological Implications of the Involvement of AtR8 lncRNA in the Defense Response
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Transcription Analysis of atr8 Mutant
4.3. Gene Expression Analysis
4.4. Salicylic Acid Treatment
4.5. Infection with Pseudomonas Syringae
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Hon, C.C.; Ramilowski, J.A.; Harshbarger, J.; Bertin, N.; Rackham, O.J.; Gough, J.; Denisenko, E.; Schmeier, S.; Poulsen, T.M.; Severin, J.; et al. An atlas of human long non-coding RNAs with accurate 5′ ends. Nature 2017, 543, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Liu, J.; Wang, H.; Wong, L.; Chua, N.H. PLncDB: Plant long non-coding RNA database. Bioinformatics 2013, 29, 1068–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amor, B.; Wirth, S.; Merchan, F.; Laporte, P.; d’Aubenton-Carafa, Y.; Hirsch, J.; Maizel, A.; Mallory, A.; Lucas, A.; Deragon, J.; et al. Novel long non-protein coding RNAs involved in Arabidopsis differentiation and stress responses. Genome Res. 2009, 19, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Stephen, S.; Taylor, J.; Helliwell, C.A.; Wang, M. Long noncoding RNAs responsive to Fusarium oxysporum infection in Arabidopsis thaliana. New Phytol. 2014, 201, 574–584. [Google Scholar] [CrossRef]
- Cui, J.; Luan, Y.; Jiang, N.; Bao, H.; Meng, J. Comparative transcriptome analysis between resistant and susceptible tomato allows the identification of lncRNA16397 conferring resistance to Phytophthora infestans by co-expressing glutaredoxin. Plant. J. 2017, 89, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Xin, M.; Wang, Y.; Yao, Y.; Song, N.; Hu, Z.; Qin, D.; Xie, C.; Peng, H.; Ni, Z.; Sun, Q. Identification and characterization of wheat long non-protein coding RNAs responsive to powdery mildew infection and heat stress by using microarray analysis and SBS sequencing. BMC Plant. Biol. 2011, 11, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Jung, C.; Xu, J.; Wang, H.; Deng, S.; Bernad, L.; Arenas-Huertero, C.; Chua, N.-H. Genome-Wide Analysis Uncovers Regulation of Long Intergenic Noncoding RNAs in Arabidopsis. Plant. Cell 2012, 24, 4333–4345. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Bai, X.; Luo, W.; Feng, Y.; Shao, X.; Bai, Q.; Sun, S.; Long, Q.; Wan, D. Genome-Wide Identification of Long Noncoding RNAs and Their Responses to Salt Stress in Two Closely Related Poplars. Front. Gen. 2019, 10, 777. [Google Scholar] [CrossRef]
- Wang, A.; Hu, J.; Gao, C.; Chen, G.; Wang, B.; Lin, C.; Song, L.; Ding, Y.; Zhou, G. Genome-wide analysis of long non-coding RNAs unveils the regulatory roles in the heat tolerance of Chinese cabbage (Brassica rapa ssp.chinensis). Sci. Rep. 2019, 9, 5002. [Google Scholar] [CrossRef] [Green Version]
- Chekanova, J.A. Long non-coding RNAs and their functions in plants. Curr. Opin. Plant. Biol. 2015, 27, 207–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, H.; Chua, N. Long noncoding RNA transcriptome of plants. Plant. Biotechnol. J. 2015, 13, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, S.; Li, J.; Sun, Q. Functions of plants long non-coding RNAs. Bioch. Biophys. Acta 2016, 1859, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Meng, X.; Dobrovolskaya, O.B.; Orlov, Y.L.; Chen, M. Non-coding RNAs and Their Roles in Stress Response in Plants. Genom. Proteom. Bioinform. 2017, 15, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zhu, H. Long Non-Coding RNAs: Rising Regulators of Plant Reproductive Development. Agronomy 2019, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Böhmdorfer, G.; Sethuraman, S.; Rowley, J.M.; Krzyszton, M.; Rothi, H.M.; Bouzit, L.; Wierzbicki, A.T. Long non-coding RNA produced by RNA polymerase V determines boundaries of heterochromatin. Elife 2016, 5, e19092. [Google Scholar] [CrossRef]
- Wu, J.; Okada, T.; Fukushima, T.; Tsudzuki, T.; Sugiura, M.; Yukawa, Y. A novel hypoxic stress-responsive long non-coding RNA transcribed by RNA polymerase III in Arabidopsis. RNA Biol. 2012, 9, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Liu, C.; Liu, Z.; Li, S.; Li, D.; Liu, S.; Huang, X.; Liu, S.; and Yukawa, Y. Pol III-dependent cabbage BoNR8 long ncRNA affects seed germination and growth in Arabidopsis. Plant. Cell Physiol. 2018, 10, 1093. [Google Scholar] [CrossRef] [Green Version]
- Röhrig, H.; Schmidt, J.; Miklashevichs, E.; Schell, J.; John, M. Soybean ENOD40 encodes two peptides that bind to sucrose synthase. Proc. National Acad. Sci. USA 2002, 99, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Steenackers, W.J.; Cesarino, I.; Klíma, P.; Quareshy, M.; Vanholme, R.; Corneillie, S.; Kumpf, R.P.; Van De Wouwer, D.; Ljung, K.; Goeminne, G.; et al. The Allelochemical MDCA Inhibits Lignification and Affects Auxin Homeostasis. Plant Physiol. 2016, 172, 874–888. [Google Scholar] [CrossRef] [Green Version]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [Green Version]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant. Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Viana, V.E.; Busanello, C.; da Maia, L.C.; Pegoraro, C.; Costa de Oliveira, A. Activation of rice WRKY transcription factors: An army of stress fighting soldiers? Curr. Opin. Plant. Biol. 2018, 45, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Bioch. Biophys. Acta 2012, 1819, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Brader, G.; Kariola, T.; Palva, T.E. WRKY70 modulates the selection of signaling pathways in plant defense. Plant. J. 2006, 46, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Amornsiripanitch, N.; Dong, X. A Genomic Approach to Identify Regulatory Nodes in the Transcriptional Network of Systemic Acquired Resistance in Plants. Plos. Pathog. 2006, 2, e123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Brader, G.; Palva, T.E. The WRKY70 Transcription Factor: A Node of Convergence for Jasmonate-Mediated and Salicylate-Mediated Signals in Plant Defense. Plant. Cell 2004, 16, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Hsu, F.C.; Chou, M.Y.; Chou, S.J.; Li, Y.R.; Peng, H.P.; Shih, M.C. Submergence confers immunity mediated by the WRKY22 transcription factor in Arabidopsis. Plant. Cell 2013, 25, 2699–2713. [Google Scholar] [CrossRef] [Green Version]
- Besseau, S.; Li, J.; Palva, T.E. WRKY54 and WRKY70 co-operate as negative regulators of leaf senescence in Arabidopsis thaliana. J. Exp. Bot. 2012, 63, 2667–2679. [Google Scholar] [CrossRef]
- Ulker, B.; Shahid Mukhtar, M.; Somssich, I.E. The WRKY70 transcription factor of Arabidopsis influences both the plant senescence and defense signaling pathways. Planta 2007, 226, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Brusslan, J.A.; Alvarez-Canterbury, A.M.; Nair, N.; Rice, J.C.; Hitchler, M.J.; Pellegrini, M. Genome-Wide Evaluation of Histone Methylation Changes Associated with Leaf Senescence in Arabidopsis. PLoS ONE 2012, 7, e33151. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wu, S. Molecular events in senescing Arabidopsis leaves. Plant. J. 2004, 39, 612–628. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Laun, T.; Zimmermann, P.; Zentgraf, U. Targets of the WRKY53 transcription factor and its role during leaf senescence in Arabidopsis. Plant. Mol. Biol. 2004, 55, 853–867. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Huhn, K.; Brandt, R.; Potschin, M.; Bieker, S.; Straub, D. REVOLUTA and WRKY53 connect early and late leaf development in Arabidopsis. Development 2014, 141, 4772–4783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Dong, Q.; Yu, D. Arabidopsis WRKY46 coordinates with WRKY70 and WRKY53 in basal resistance against pathogen Pseudomonas syringae. Plant. Sci 2012, 185–186, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Glazebrook, J.; Clarke, J.D.; Volko, S.; Dong, X. The Arabidopsis NPR1 Gene That Controls Systemic Acquired Resistance Encodes a Novel Protein Containing Ankyrin Repeats. Cell 1997, 88, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Berardini, T.; Mundodi, S.; Reiser, L.; Huala, E.; García-Hernández, M.; Zhang, P.; Mueller, L.A.; Yoon, J.; Doyle, A.; Lander, G.C.; et al. Functional Annotation of the Arabidopsis Genome Using Controlled Vocabularies1. Plant Physiol. 2004, 135, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Zavaliev, R.; Dong, X. Membrane Trafficking in Plant Immunity. Mol. Plant. 2017, 10, 1026–1034. [Google Scholar] [CrossRef]
- Bacete, L.; Mélida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. Cell Mol. Biol. 2017, 93, 614–636. [Google Scholar] [CrossRef]
- Benfey, P.N.; Linstead, P.J.; Roberts, K.; Schiefelbein, J.W.; Hauser, M.T.; Aeschbacher, R.A. Root development in Arabidopsis: Four mutants with dramatically altered root morphogenesis. Development 1993, 1, 57–70. [Google Scholar]
- Ji, W.C.; Jun, L. Control of Asymmetric Cell Divisions during root ground tissue maturation. Mol. Cells 2016, 7, 524–529. [Google Scholar]
- Levesque, M.P.; Vernoux, T.; Busch, W.; Cui, H.; Wang, J.Y.; Blilou, I.; Hassan, H.; Nakajima, K.; Matsumoto, N.; Lohmann, J.U.; et al. Whole-Genome Analysis of the SHORT-ROOT Developmental Pathway in Arabidopsis. Plos. Biol. 2006, 4, e143. [Google Scholar]
- Dhondt, S.; Coppens, F.; Winter, F.; Swarup, K.; Merks, R.M.; Inzé, D.; Bennett, M.J.; Beemster, G.T. SHORT-ROOT and SCARECROW Regulate Leaf Growth in Arabidopsis by Stimulating S-Phase Progression of the Cell Cycle. Plant. Physiol. 2010, 154, 1183–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Y.; Stahl, Y.; Weidtkamp-Peters, S.; Postma, M.; Zhou, W.; Goedhart, J.; Sánchez-Pérez, M.-I.; Gadella, T.W.; Simon, R.; Scheres, B.; et al. In vivo FRET–FLIM reveals cell-type-specific protein interactions in Arabidopsis roots. Nature 2017, 548, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Lakhotia, S.C. Long non-coding RNAs coordinate cellular responses to stress. Wiley Interdiscip. Rev. RNA 2012, 3, 779–796. [Google Scholar] [CrossRef]
- Seo, J.; Sun, H.-X.; Park, B.; Huang, C.-H.; Yeh, S.-D.; Jung, C.; Chua, N.-H. ELF18-Induced Long-Noncoding RNA Associates with Mediator to Enhance Expression of Innate Immune Response Genes in Arabidopsis. Plant. Cell 2017, 29, 1024–1038. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Chen, C.; Chen, Z. Expression profiles of the Arabidopsis WRKY gene superfamily during plant defense response. Plant. Mol. Biol. 2003, 51, 21–37. [Google Scholar] [CrossRef]
- Fukushima, S.; Mori, M.; Sugano, S.; Takatsuji, H. Transcription Factor WRKY62 Plays a Role in Pathogen Defense and Hypoxia-Responsive Gene Expression in Rice. Plant. Cell Physiol. 2016, 57, 2541–2551. [Google Scholar] [CrossRef] [Green Version]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant. Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Yang, Y.; Zhou, Y.; Zhou, J.; Fan, B.; Yu, J.-Q.; Chen, Z. Protein-protein interactions in the regulation of WRKY transcription factors. Mol. Plant. 2013, 6, 287–300. [Google Scholar] [CrossRef] [Green Version]
- Brodersen, P.; Malinovsky, F.; Hématy, K.; Newman, M.-A.; Mundy, J. The Role of Salicylic Acid in the Induction of Cell Death in Arabidopsis acd11. Plant. Physiol. 2005, 138, 1037–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrath, C.; Métraux, J.-P. Salicylic Acid Induction–Deficient Mutants of Arabidopsis Express PR-2 and PR-5 and Accumulate High Levels of Camalexin after Pathogen Inoculation. Plant. Cell 1999, 11, 1393–1404. [Google Scholar] [PubMed] [Green Version]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.S.; Jung, C.; Lee, S.; Min, K.; Lee, Y.-W.; Choi, Y.; Lee, J.S.; Song, J.T.; Kim, J.-K.; Choi, Y.D. AtMYB44 regulates WRKY70 expression and modulates antagonistic interaction between salicylic acid and jasmonic acid signaling. Plant J. 2012, 73, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Huang, X.; Liu, Z.; Li, S.; Okada, T.; Yukawa, Y.; Wu, J. Effect of AtR8 lncRNA partial deletion on Arabidopsis seed germination. Mol. Soil Biol. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Fu, Z.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Qi, G.; Chen, J.; Chang, M.; Chen, H.; Hall, K.; Korin, J.; Liu, F.; Wang, D.; Fu, Z. Pandemonium Breaks Out: Disruption of Salicylic Acid-Mediated Defense by Plant Pathogens. Mol. Plant. 2018, 11, 1427–1439. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Detchemendy, T.; Pajerowska-Mukhtar, K.; Mukhtar, M.S. NPR1 in JazzSet with Pathogen Effectors. Trends Plant. Sci. 2018, 23, 469–472. [Google Scholar] [CrossRef]
- Fan, W.; Dong, X. In Vivo Interaction between NPR1 and Transcription Factor TGA2 Leads to Salicylic Acid–Mediated Gene Activation in Arabidopsis. Plant Cell Online 2002, 14, 1377–1389. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.-M.; Zhu, Q.; Gao, B.-D.; Ke, S.-Y.; Yu, W.-C.; Xie, D.-X.; Peng, W. Transcription factor WRKY70 displays important but no indispensable roles in jasmonate and salicylic acid signaling. J. Integr. Plant. Biol. 2008, 50, 630–637. [Google Scholar] [CrossRef]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S. Growth–Defense Tradeoffs in Plants: A Balancing Act to Optimize Fitness. Mol. Plant. 2014, 7, 1267–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smakowska, E.; Kong, J.; Busch, W.; Belkhadi, Y. Organ-specific regulation of growth-defense tradeoffs by plants. Curr. Opin. Plant. Biol. 2016, 29, 129–137. [Google Scholar] [CrossRef] [PubMed]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant. Signal. Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leymarie, J.; Vitkauskaité, G.; Hoang, H.; Gendreau, E.; Chazoule, V.; Meimoun, P.; Corbineau, F.; El-Maarouf-Bouteau, H.; Bailly, C. Role of Reactive Oxygen Species in the Regulation of Arabidopsis Seed Dormancy. Plant. Cell Physiol. 2012, 53, 96–106. [Google Scholar] [CrossRef] [Green Version]
- De-la-Peña, C.; Badri, D.V.; Lei, Z.; Watson, B.S.; Brandão, M.M.; Silva-Filho, M.C.; Sumner, L.W.; Vivanco, J.M. Root Secretion of Defense-related Proteins Is Development-dependent and Correlated with Flowering Time. J. Biol. Chem. 2010, 285, 30654–30665. [Google Scholar] [CrossRef] [Green Version]
- Coninck, B.; Timmermans, P.; Vos, C.; Cammue, B.P.; Kazan, K. What lies beneath: Belowground defense strategies in plants. Trends Plant. Sci. 2015, 20, 91–101. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TAIR ID | Fold Change | Regulation | Expression Level | Gene Title |
|---|---|---|---|---|
| AT1G80840 | 9.87 | up | 1137.8 | WRKY40 |
| AT4G23810 | 7.95 | up | 1595.3 | WRKY53 |
| AT2G46400 | 5.52 | up | 786.6 | WRKY46 |
| AT5G22570 | 5.03 | up | 211.4 | WRKY38 |
| AT2G40750 | 4.31 | up | 1474.1 | WRKY54 |
| AT5G13080 | 4.14 | up | 464.9 | WRKY75 |
| AT3G56400 | 3.94 | up | 3225.8 | WRKY70 |
| AT2G38470 | 3.39 | up | 1878.9 | WRKY33 |
| AT5G49520 | 3.08 | up | 475.3 | WRKY48 |
| AT2G25000 | 2.21 | up | 784.7 | WRKY60 |
| AT1G66550 | 2.04 | up | 9.4 | WRKY67 |
| AT4G31550 | 2.04 | up | 1212.9 | WRKY11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Nayar, S.; Jia, H.; Kapoor, S.; Wu, J.; Yukawa, Y. The Arabidopsis Hypoxia Inducible AtR8 Long Non-Coding RNA also Contributes to Plant Defense and Root Elongation Coordinating with WRKY Genes under Low Levels of Salicylic Acid. Non-Coding RNA 2020, 6, 8. https://doi.org/10.3390/ncrna6010008
Li S, Nayar S, Jia H, Kapoor S, Wu J, Yukawa Y. The Arabidopsis Hypoxia Inducible AtR8 Long Non-Coding RNA also Contributes to Plant Defense and Root Elongation Coordinating with WRKY Genes under Low Levels of Salicylic Acid. Non-Coding RNA. 2020; 6(1):8. https://doi.org/10.3390/ncrna6010008
Chicago/Turabian StyleLi, Shuang, Saraswati Nayar, HuiYuan Jia, Sanjay Kapoor, Juan Wu, and Yasushi Yukawa. 2020. "The Arabidopsis Hypoxia Inducible AtR8 Long Non-Coding RNA also Contributes to Plant Defense and Root Elongation Coordinating with WRKY Genes under Low Levels of Salicylic Acid" Non-Coding RNA 6, no. 1: 8. https://doi.org/10.3390/ncrna6010008
APA StyleLi, S., Nayar, S., Jia, H., Kapoor, S., Wu, J., & Yukawa, Y. (2020). The Arabidopsis Hypoxia Inducible AtR8 Long Non-Coding RNA also Contributes to Plant Defense and Root Elongation Coordinating with WRKY Genes under Low Levels of Salicylic Acid. Non-Coding RNA, 6(1), 8. https://doi.org/10.3390/ncrna6010008