
Tumor Suppressive Effects of GAS5 in Cancer Cells

 , and
, and
Abstract
:1. Introduction
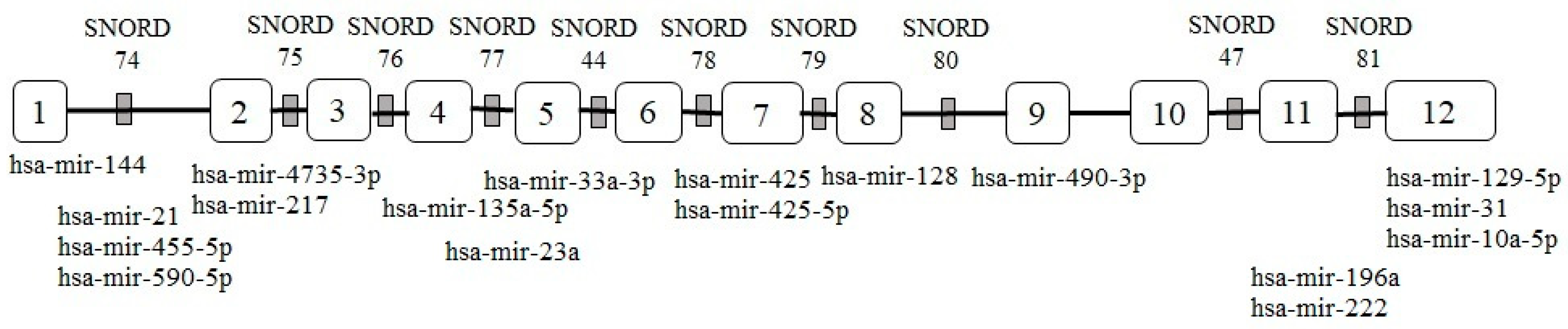
2. GAS5 Expression Is Regulated by the mTOR Signaling Pathway
3. Role of GAS5 in Oncogenic Signaling Pathways
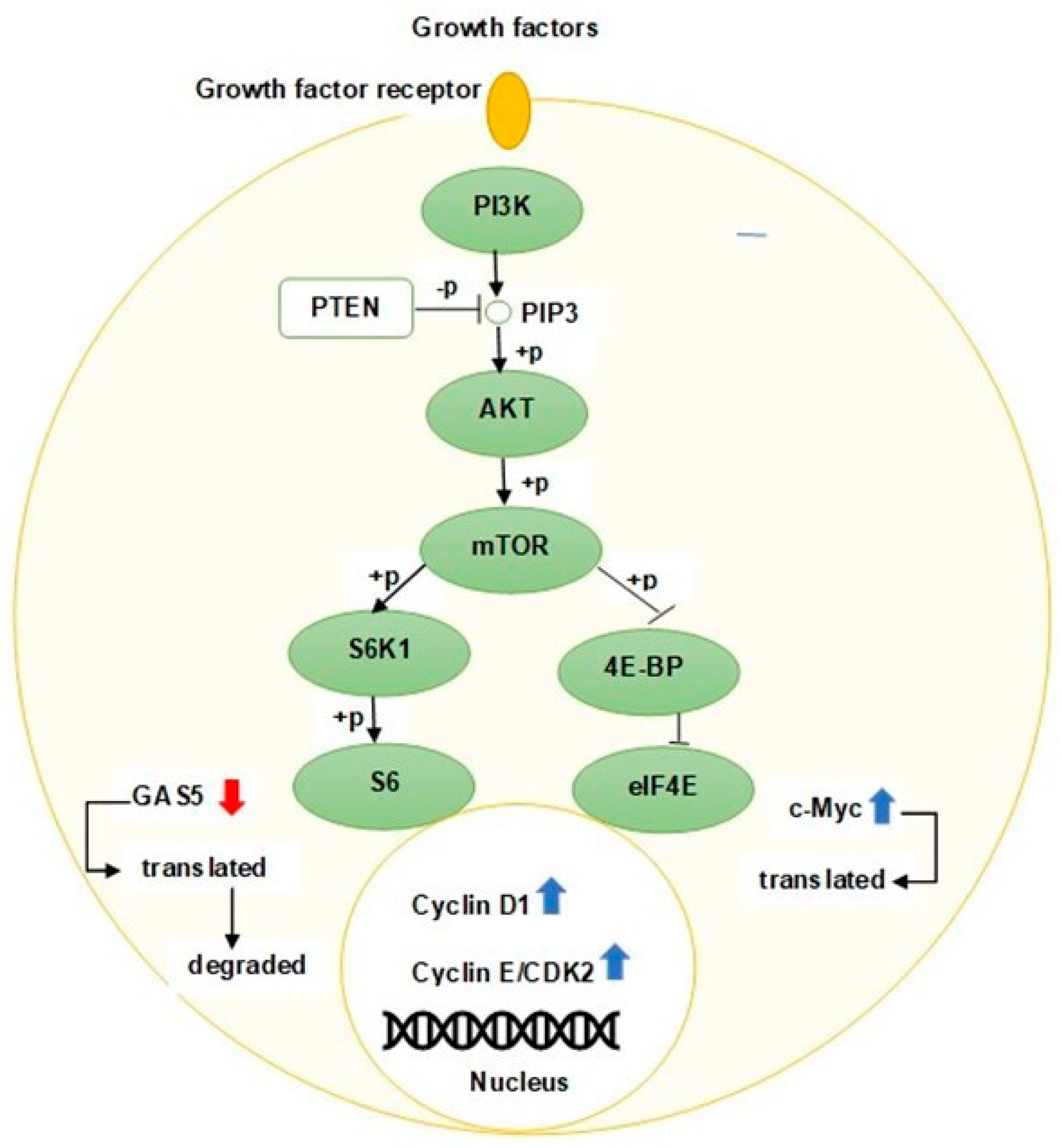
3.1. PI3K/AKT/mTOR Pathway
3.2. PTEN/AKT Pathway
4. GAS5 as Part of Cell Cycle Regulatory Mechanism
4.1. c-Myc Expression
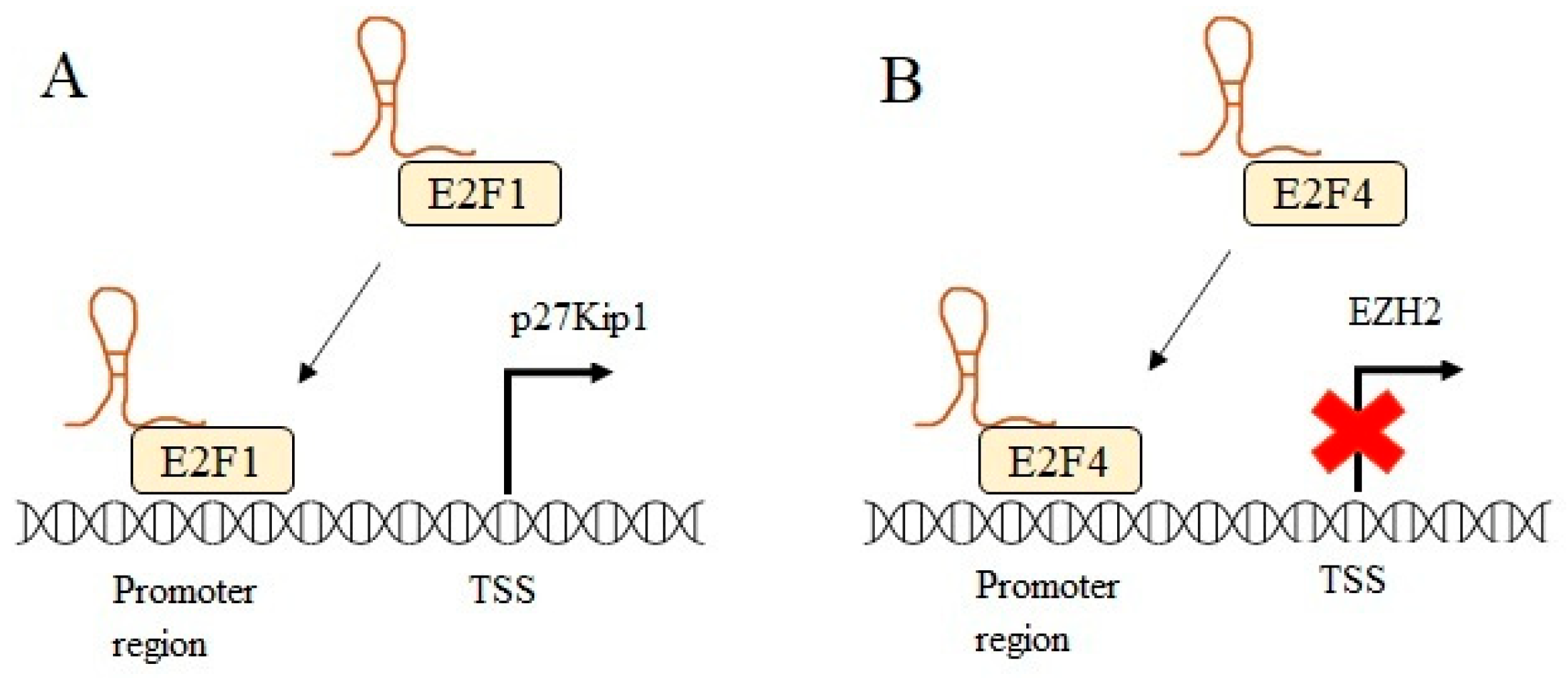
4.2. CDK Inhibitors
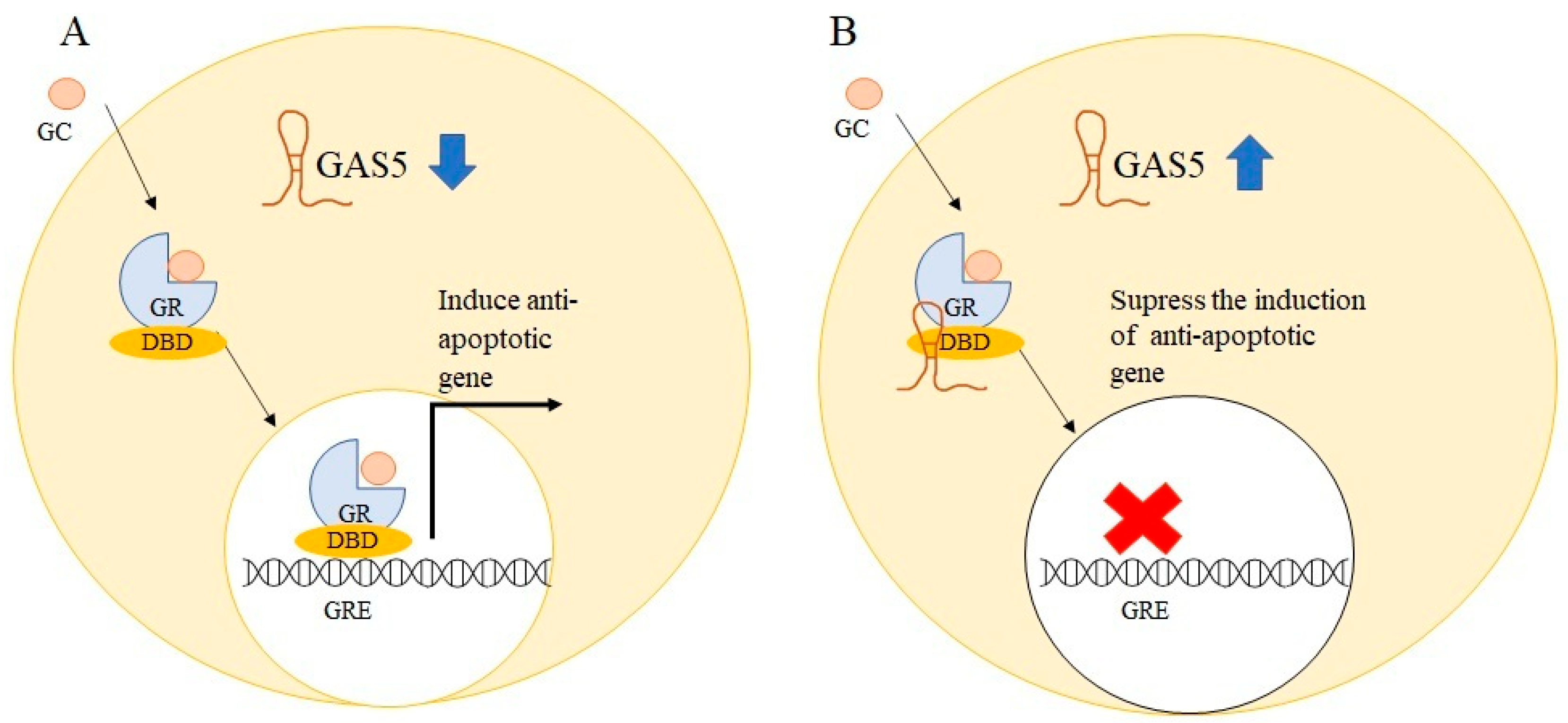
5. GAS5 Regulates Cellular Apoptosis
6. Genetic Variants Affect GAS5 Expression and Cancer Susceptibility
7. GAS5 Regulates Target Genes via Competing Endogenous RNA (ceRNA) Network
7.1. FOXO
7.2. hZIP1
8. GAS5 Modulates Chemosensitivity and Radiosensitivity in Cancer Cells
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Diamantopoulos, M.A.; Tsiakanikas, P.; Scorilas, A. Non-coding RNAs: The riddle of the transcriptome and their perspectives in cancer. Ann. Transl. Med. 2018, 6, 241. [Google Scholar] [CrossRef] [PubMed]
- Hangauer, M.J.; Vaughn, I.W.; McManus, M.T. Pervasive Transcription of the Human Genome Produces Thousands of Previously Unidentified Long Intergenic Noncoding RNAs. PLOS Genet. 2013, 9, e1003569. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Guzel, E.; Okyay, T.M.; Yalcinkaya, B.; Karacaoglu, S.; Gocmen, M.; Akcakuyu, M.H. Tumor suppressor and oncogenic role of long non-coding RNAs in cancer. North Clin. Istanb. 2020, 7, 81–86. [Google Scholar] [CrossRef]
- Goustin, A.S.; Thepsuwan, P.; Kosir, M.A.; Lipovich, L. The Growth-Arrest-Specific (GAS)-5 Long Non-Coding RNA: A Fascinating lncRNA Widely Expressed in Cancers. Noncoding RNA 2019, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Steitz, J.A. Classification of gas5 as a multi-small-nucleolar-RNA (snoRNA) host gene and a member of the 5′-terminal oligopyrimidine gene family reveals common features of snoRNA host genes. Mol. Cell Biol. 1998, 18, 6897–6909. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.; King, R.M.; Philipson, L. Genes specifically expressed at growth arrest of mammalian cells. Cell 1988, 54, 787–793. [Google Scholar] [CrossRef]
- Coccia, E.M.; Cicala, C.; Charlesworth, A.; Ciccarelli, C.; Rossi, G.B.; Philipson, L.; Sorrentino, V. Regulation and expression of a growth arrest-specific gene (gas5) during growth, differentiation, and development. Mol. Cell Biol. 1992, 12, 3514–3521. [Google Scholar] [CrossRef]
- Ciccarelli, C.; Philipson, L.; Sorrentino, V. Regulation of expression of growth arrest-specific genes in mouse fibroblasts. Mol. Cell Biol. 1990, 10, 1525–1529. [Google Scholar] [CrossRef]
- Mourtada-Maarabouni, M.; Hasan, A.M.; Farzaneh, F.; Williams, G.T. Inhibition of human T-cell proliferation by mammalian target of rapamycin (mTOR) antagonists requires noncoding RNA growth-arrest-specific transcript 5 (GAS5). Mol. Pharmacol. 2010, 78, 19–28. [Google Scholar] [CrossRef]
- Mourtada-Maarabouni, M.; Williams, G.T. Growth arrest on inhibition of nonsense-mediated decay is mediated by noncoding RNA GAS5. BioMed Res. Int. 2013, 2013, 358015. [Google Scholar] [CrossRef] [PubMed]
- Tani, H.; Torimura, M.; Akimitsu, N. The RNA degradation pathway regulates the function of GAS5 a non-coding RNA in mammalian cells. PLoS ONE 2013, 8, e55684. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourtada-Maarabouni, M.; Pickard, M.R.; Hedge, V.L.; Farzaneh, F.; Williams, G.T. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene 2009, 28, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Liu, W.; Li, F.; Zhao, W.; Qin, C. Decreased expression of lncRNA GAS5 predicts a poor prognosis in cervical cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 6776–6783. [Google Scholar] [PubMed]
- Li, J.; Wang, Y.; Zhang, C.-G.; Xiao, H.-J.; Xiao, H.-J.; Hu, J.-M.; Hou, J.-M.; He, J.-D. Effect of long non-coding RNA Gas5 on proliferation, migration, invasion and apoptosis of colorectal cancer HT-29 cell line. Cancer Cell Int. 2018, 18, 4. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Jin, F.-y.; Xia, R.; Kong, R.; Li, J.-h.; Xu, T.-p.; Liu, Y.-w.; Zhang, E.-b.; Liu, X.-h.; De, W. Decreased expression of long noncoding RNA GAS5 indicates a poor prognosis and promotes cell proliferation in gastric cancer. BMC Cancer 2014, 14, 319. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Li, C.; Lan, T.; Wu, L.; Yuan, Y.; Liu, Q.; Liu, Z. Decreased expression of long non-coding RNA GAS5 indicates a poor prognosis and promotes cell proliferation and invasion in hepatocellular carcinoma by regulating vimentin. Mol. Med. Rep. 2016, 13, 1541–1550. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.P.; Gao, W.S.; Huo, J.X.; Yang, Z.S. Long non-coding RNA GAS5 functions as a tumor suppressor in renal cell carcinoma. Asian Pac. J. Cancer Prev. 2013, 14, 1077–1082. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xin, S.; Zhang, K.; Shi, R.; Bao, X. Low GAS5 Levels as a Predictor of Poor Survival in Patients with Lower-Grade Gliomas. J. Oncol. 2019, 2019, 1785042. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Kong, C.; Liu, X.; Bi, J.; Li, Z.; Li, Z.; Zhu, Y.; Zhang, Z. GAS5 functions as a ceRNA to regulate hZIP1 expression by sponging miR-223 in clear cell renal cell carcinoma. Am. J. Cancer Res. 2018, 8, 1414–1426. [Google Scholar] [PubMed]
- Lyu, K.; Xu, Y.; Yue, H.; Li, Y.; Zhao, J.; Chen, L.; Wu, J.; Zhu, X.; Chai, L.; Li, C.; et al. Long Noncoding RNA GAS5 Acts As A Tumor Suppressor In Laryngeal Squamous Cell Carcinoma Via miR-21. Cancer Manag. Res. 2019, 11, 8487–8498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, D.; Zhou, C.; Lu, H.; Xu, R.; Xu, X.; He, X. LncRNA GAS5 inhibits proliferation and progression of prostate cancer by targeting miR-103 through AKT/mTOR signaling pathway. Tumor Biol. 2016, 37, 16187–16197. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, S.; Yang, X.; Li, X.; Chen, R. Association between polymorphism in the promoter region of lncRNA GAS5 and the risk of colorectal cancer. Biosci. Rep. 2019, 39, BSR20190091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, M.-H.; Lu, H.-J.; Lin, C.-W.; Lee, C.-Y.; Yang, S.-J.; Wu, P.-H.; Chen, M.-K.; Yang, S.-F. Genetic Variants of lncRNA GAS5 Are Associated with the Clinicopathologic Development of Oral Cancer. J. Pers. Med. 2021, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Feng, L.; Li, F.; Xue, Y.; Li, C.; Wang, H. A novel functional indel polymorphism within long non-coding RNAs growth arrest specific 5 conferred risk for cervical squamous cell carcinoma in Chinese Han populations. Transl. Cancer Res. 2017, 6, 424–431. [Google Scholar] [CrossRef]
- Xu, L.; Xia, C.; Xue, B.; Sheng, F.; Xiong, J.; Wang, S. A promoter variant of lncRNA GAS5 is functionally associated with the development of osteosarcoma. J. Bone Oncol. 2018, 12, 23–26. [Google Scholar] [CrossRef]
- Gendron, P.; Lemieux, S.; Major, F. Quantitative analysis of nucleic acid three-dimensional structures. J. Mol. Biol. 2001, 308, 919–936. [Google Scholar] [CrossRef] [Green Version]
- Fingar, D.C.; Salama, S.; Tsou, C.; Harlow, E.; Blenis, J. Mammalian cell size is controlled by mTOR and its downstream targets S6K1 and 4EBP1/eIF4E. Genes Dev. 2002, 16, 1472–1487. [Google Scholar] [CrossRef] [Green Version]
- Kawasome, H.; Papst, P.; Webb, S.; Keller, G.M.; Johnson, G.L.; Gelfand, E.W.; Terada, N. Targeted disruption of p70(s6k) defines its role in protein synthesis and rapamycin sensitivity. Proc. Natl. Acad. Sci. USA 1998, 95, 5033–5038. [Google Scholar] [CrossRef] [Green Version]
- Jefferies, H.B.; Fumagalli, S.; Dennis, P.B.; Reinhard, C.; Pearson, R.B.; Thomas, G. Rapamycin suppresses 5’TOP mRNA translation through inhibition of p70s6k. EMBO J. 1997, 16, 3693–3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raho, G.; Barone, V.; Rossi, D.; Philipson, L.; Sorrentino, V. The gas 5 gene shows four alternative splicing patterns without coding for a protein. Gene 2000, 256, 13–17. [Google Scholar] [CrossRef]
- Gingras, A.C.; Kennedy, S.G.; O’Leary, M.A.; Sonenberg, N.; Hay, N. 4E-BP1, a repressor of mRNA translation, is phosphorylated and inactivated by the Akt(PKB) signaling pathway. Genes Dev. 1998, 12, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Heesom, K.J.; Denton, R.M. Dissociation of the eukaryotic initiation factor-4E/4E-BP1 complex involves phosphorylation of 4E-BP1 by an mTOR-associated kinase. FEBS Lett. 1999, 457, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Rosenwald, I.B.; Lazaris-Karatzas, A.; Sonenberg, N.; Schmidt, E.V. Elevated levels of cyclin D1 protein in response to increased expression of eukaryotic initiation factor 4E. Mol. Cell Biol. 1993, 13, 7358–7363. [Google Scholar] [CrossRef]
- Quelle, D.E.; Ashmun, R.A.; Shurtleff, S.A.; Kato, J.Y.; Bar-Sagi, D.; Roussel, M.F.; Sherr, C.J. Overexpression of mouse D-type cyclins accelerates G1 phase in rodent fibroblasts. Genes Dev. 1993, 7, 1559–1571. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Zhang, X.; Liu, D. Overexpression of long noncoding RNA GAS5 suppresses tumorigenesis and development of gastric cancer by sponging miR-106a-5p through the Akt/mTOR pathway. Biol. Open 2019, 8, bio041343. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, M.; Ma, L.; Dang, X.; Du, G. LncRNA GAS5 Suppresses the Proliferation and Invasion of Osteosarcoma Cells via the miR-23a-3p/PTEN/PI3K/AKT Pathway. Cell Transplant. 2020, 29, 0963689720953093. [Google Scholar] [CrossRef]
- Gao, X.; Neufeld, T.P.; Pan, D. Drosophila PTEN regulates cell growth and proliferation through PI3K-dependent and -independent pathways. Dev. Biol. 2000, 221, 404–418. [Google Scholar] [CrossRef] [Green Version]
- Ramaswamy, S.; Nakamura, N.; Vazquez, F.; Batt, D.B.; Perera, S.; Roberts, T.M.; Sellers, W.R. Regulation of G1 progression by the PTEN tumor suppressor protein is linked to inhibition of the phosphatidylinositol 3-kinase/Akt pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 2110–2115. [Google Scholar] [CrossRef] [Green Version]
- Kanamori, Y.; Kigawa, J.; Itamochi, H.; Shimada, M.; Takahashi, M.; Kamazawa, S.; Sato, S.; Akeshima, R.; Terakawa, N. Correlation between Loss of PTEN Expression and Akt Phosphorylation in Endometrial Carcinoma. Clin. Cancer Res. 2001, 7, 892–895. [Google Scholar] [PubMed]
- Liu, L.; Huang, L.; He, J.; Cai, S.; Weng, Y.; Huang, S.; Ma, S. PTEN inhibits non-small cell lung cancer cell growth by promoting G(0)/G(1) arrest and cell apoptosis. Oncol. Lett. 2019, 17, 1333–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Chen, J.; Ou, B.; Liu, C.; Zou, Y.; Chen, Q. GAS5 knockdown reduces the chemo-sensitivity of non-small cell lung cancer (NSCLC) cell to cisplatin (DDP) through regulating miR-21/PTEN axis. Biomed. Pharmacother. 2017, 93, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ren, P.; Zhang, Y.; Gong, B.; Yu, D.; Sun, X. Long non-coding RNA GAS5 increases the radiosensitivity of A549 cells through interaction with the miR-21/PTEN/Akt axis. Oncol. Rep. 2020, 43, 897–907. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Ke, S.; Li, M.; Lin, C.; Liu, X.; Pan, Q. Downregulation of LncRNA GAS5 promotes liver cancer proliferation and drug resistance by decreasing PTEN expression. Mol. Genet. Genom. 2020, 295, 251–260. [Google Scholar] [CrossRef]
- Liu, L.; Wang, H.J.; Meng, T.; Lei, C.; Yang, X.H.; Wang, Q.S.; Jin, B.; Zhu, J.F. lncRNA GAS5 Inhibits Cell Migration and Invasion and Promotes Autophagy by Targeting miR-222-3p via the GAS5/PTEN-Signaling Pathway in CRC. Mol. Ther. Nucleic Acids 2019, 17, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-F.; Ye, Y.; Zhao, S.-J. LncRNA Gas5 acts as a ceRNA to regulate PTEN expression by sponging miR-222-3p in papillary thyroid carcinoma. Oncotarget 2017, 9, 3519–3530. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Song, W.Q.; Sun, P.; Jin, L.; Dai, H.Y. LncRNA-GAS5 induces PTEN expression through inhibiting miR-103 in endometrial cancer cells. J. Biomed. Sci. 2015, 22, 100. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.-Q.; Wang, J.-f.; Chen, D.-H.; Ma, X.-S.; Wu, Y.; Tang, Z.; Dang, X.-W. Long non-coding RNA GAS5 suppresses pancreatic cancer metastasis through modulating miR-32-5p/PTEN axis. Cell Biosci. 2017, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- West, M.J.; Stoneley, M.; Willis, A.E. Translational induction of the c-myc oncogene via activation of the FRAP/TOR signalling pathway. Oncogene 1998, 17, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Vlach, J.; Hennecke, S.; Alevizopoulos, K.; Conti, D.; Amati, B. Growth arrest by the cyclin-dependent kinase inhibitor p27Kip1 is abrogated by c-Myc. EMBO J. 1996, 15, 6595–6604. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Lou, Z.; Gupta, M. The long non-coding RNA GAS5 cooperates with the eukaryotic translation initiation factor 4E to regulate c-Myc translation. PLoS ONE 2014, 9, e107016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, S.; Edwards, M.C.; Bai, C.; Parker, S.; Zhang, P.; Baldini, A.; Harper, J.; Elledge, S. p57KIP2, a structurally distinct member of the p21CIP1 Cdk inhibitor family, is a candidate tumor suppressor gene. Genes Dev. 1995, 9 6, 650–662. [Google Scholar] [CrossRef] [Green Version]
- Sherr, C.J.; Roberts, J.M. Inhibitors of mammalian G1 cyclin-dependent kinases. Genes Dev. 1995, 9, 1149–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, G.; Liu, D.; Huang, C.; Wang, M.; Xiao, X.; Zeng, F.; Wang, L.; Jiang, G. LncRNA GAS5 Inhibits Cellular Proliferation by Targeting P27(Kip1). Mol. Cancer Res. 2017, 15, 789–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Yu, Z.; Meng, X.; Zhou, S.; Xiao, S.; Li, X.; Liu, S.; Yu, P. Long noncoding RNA GAS5 impairs the proliferation and invasion of endometrial carcinoma induced by high glucose via targeting miR-222-3p/p27. Am. J. Transl. Res. 2019, 11, 2413–2421. [Google Scholar]
- Liu, Y.; Zhao, J.; Zhang, W.; Gan, J.; Hu, C.; Huang, G.; Zhang, Y. lncRNA GAS5 enhances G1 cell cycle arrest via binding to YBX1 to regulate p21 expression in stomach cancer. Sci. Rep. 2015, 5, 10159. [Google Scholar] [CrossRef] [Green Version]
- Zingg, D.; Debbache, J.; Schaefer, S.M.; Tuncer, E.; Frommel, S.C.; Cheng, P.; Arenas-Ramirez, N.; Haeusel, J.; Zhang, Y.; Bonalli, M.; et al. The epigenetic modifier EZH2 controls melanoma growth and metastasis through silencing of distinct tumour suppressors. Nat. Commun. 2015, 6, 6051. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Yan, Z.; Hu, F.; Wei, W.; Yang, C.; Sun, Z. Long non-coding RNA GAS5 accelerates oxidative stress in melanoma cells by rescuing EZH2-mediated CDKN1C downregulation. Cancer Cell Int. 2020, 20, 116. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Guo, C.; Wang, L.; Luo, G.; Huang, C.; Li, Y.; Liu, D.; Zeng, F.; Jiang, G.; Xiao, X. Long noncoding RNA GAS5 promotes bladder cancer cells apoptosis through inhibiting EZH2 transcription. Cell Death Dis. 2018, 9, 238. [Google Scholar] [CrossRef]
- Volden, P.A.; Conzen, S.D. The influence of glucocorticoid signaling on tumor progression. Brain Behav. Immun. 2013, 30, S26–S31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pufall, M.A. Glucocorticoids and Cancer. Adv. Exp. Med. Biol. 2015, 872, 315–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakour, N.; Moriarty, F.; Moore, G.; Robson, T.; Annett, S.L. Prognostic Significance of Glucocorticoid Receptor Expression in Cancer: A Systematic Review and Meta-Analysis. Cancers 2021, 13, 1649. [Google Scholar] [CrossRef] [PubMed]
- Rutz, H.P.; Herr, I. Interference of glucocorticoids with apoptosis signaling and host-tumor interactions. Cancer Biol. Ther. 2004, 3, 715–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci. Signal. 2010, 3, ra8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickard, M.R.; Williams, G.T. The hormone response element mimic sequence of GAS5 lncRNA is sufficient to induce apoptosis in breast cancer cells. Oncotarget 2016, 7, 10104–10116. [Google Scholar] [CrossRef] [Green Version]
- Pickard, M.R.; Mourtada-Maarabouni, M.; Williams, G.T. Long non-coding RNA GAS5 regulates apoptosis in prostate cancer cell lines. Biochim. Biophys. Acta 2013, 1832, 1613–1623. [Google Scholar] [CrossRef] [Green Version]
- Runnebaum, I.B.; Brüning, A. Glucocorticoids Inhibit Cell Death in Ovarian Cancer and Up-regulate Caspase Inhibitor cIAP2. Clin. Cancer Res. 2005, 11, 6325–6332. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Zhang, D.-Y.; Li, X.; Yuan, X.-Q.; Yang, Y.-L.; Zhu, K.-W.; Zeng, H.; Li, X.-L.; Cao, S.; Zhou, H.-H.; et al. Long non-coding RNA GAS5 polymorphism predicts a poor prognosis of acute myeloid leukemia in Chinese patients via affecting hematopoietic reconstitution. Leuk. Lymphoma 2016, 58, 1948–1957. [Google Scholar] [CrossRef]
- Tao, R.; Hu, S.; Wang, S.; Zhou, X.; Zhang, Q.; Wang, C.; Zhao, X.; Zhou, W.; Zhang, S.; Li, C.; et al. Association between indel polymorphism in the promoter region of lncRNA GAS5 and the risk of hepatocellular carcinoma. Carcinogenesis 2015, 36, 1136–1143. [Google Scholar] [CrossRef] [Green Version]
- Weng, S.-L.; Ng, S.-C.; Lee, Y.-C.; Hsiao, Y.-H.; Hsu, C.-F.; Yang, S.-F.; Wang, P.-H. The relationships of genetic polymorphisms of the long noncoding RNA growth arrest-specific transcript 5 with uterine cervical cancer. Int. J. Med. Sci. 2020, 17, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhang, N.; Zheng, Y.; Chen, Y.-D.; Liu, J.; Yang, M. LncRNA GAS5 Indel Genetic Polymorphism Contributes to Glioma Risk Through Interfering Binding of Transcriptional Factor TFAP2A. DNA Cell Biol. 2018, 37, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, R.; Shahangian, S.S.; Salehi, Z.; Mashayekhi, F.; Talesh Sasani, S.; Mirzanezhad, L. Influence of a 5-bp Indel Polymorphism at Promoter of the GAS5 lncRNA and Risk of Breast Cancer. Asian Pac. J. Cancer Prev. 2020, 21, 3705–3710. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Wang, S.-S.; Yang, C.-K.; Li, J.-R.; Chen, C.-S.; Hung, S.-C.; Chiu, K.-Y.; Cheng, C.-L.; Ou, Y.-C.; Yang, S.-F. Impact of GAS5 genetic polymorphism on prostate cancer susceptibility and clinicopathologic characteristics. Int. J. Med. Sci. 2019, 16, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Rakhshan, A.; Esmaeili, M.H.; Kahaei, M.S.; Taheri, M.; Omrani, M.D.; Noroozi, R.; Ghafouri-Fard, S. A Single Nucleotide Polymorphism in GAS5 lncRNA is Associated with Risk of Bladder Cancer in Iranian Population. Pathol. Oncol. Res. 2020, 26, 1251–1254. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, Y.; Zhao, Y.; Jin, Y.; An, L.; Wu, B.; Liu, Z.; Chen, X.; Zhou, H.; Wang, H.; et al. Genetic polymorphisms of long non-coding RNA GAS5 predict platinum-based concurrent chemoradiotherapy response in nasopharyngeal carcinoma patients. Oncotarget 2017, 8, 62286–62297. [Google Scholar] [CrossRef] [Green Version]
- Weng, W.-C.; Chen, C.-J.; Chen, P.-N.; Wang, S.-S.; Hsieh, M.-J.; Yang, S.-F. Impact of Gene Polymorphisms in GAS5 on Urothelial Cell Carcinoma Development and Clinical Characteristics. Diagnostics 2020, 10, 260. [Google Scholar] [CrossRef]
- Ala, U. Competing Endogenous RNAs, Non-Coding RNAs and Diseases: An Intertwined Story. Cells 2020, 9, 1574. [Google Scholar] [CrossRef]
- Lekka, E.; Hall, J. Noncoding RNAs in disease. FEBS Lett. 2018, 592, 2884–2900. [Google Scholar] [CrossRef]
- Cheng, K.; Zhao, Z.; Wang, G.; Wang, J.; Zhu, W. lncRNA GAS5 inhibits colorectal cancer cell proliferation via the miR-182-5p/FOXO3a axis. Oncol. Rep. 2018, 40, 2371–2380. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Y.; Zheng, J.; Liu, X.; Chen, J.; Liu, L.; Wang, P.; Xue, Y. GAS5 suppresses malignancy of human glioma stem cells via a miR-196a-5p/FOXO1 feedback loop. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1605–1617. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, J.; Wang, Z.; Wang, P.; Gao, X.; Wang, Y. Long noncoding RNA GAS5 suppresses triple negative breast cancer progression through inhibition of proliferation and invasion by competitively binding miR-196a-5p. Biomed. Pharmacother. 2018, 104, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Bahceci, I.; Dogrusoz, U.; La, K.C.; Babur, Ö.; Gao, J.; Schultz, N. PathwayMapper: A collaborative visual web editor for cancer pathways and genomic data. Bioinformatics 2017, 33, 2238–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt-Ney, M. The FOXO’s Advantages of Being a Family: Considerations on Function and Evolution. Cells 2020, 9, 787. [Google Scholar] [CrossRef] [Green Version]
- Jiramongkol, Y.; Lam, E.W.F. FOXO transcription factor family in cancer and metastasis. Cancer Metastasis Rev. 2020, 39, 681–709. [Google Scholar] [CrossRef]
- Gaither, L.A.; Eide, D.J. The human ZIP1 transporter mediates zinc uptake in human K562 erythroleukemia cells. J. Biol. Chem. 2001, 276, 22258–22264. [Google Scholar] [CrossRef] [Green Version]
- Costello, L.C.; Franklin, R.B. Decreased zinc in the development and progression of malignancy: An important common relationship and potential for prevention and treatment of carcinomas. Expert Opin. Ther. Targets 2017, 21, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Franklin, R.B.; Feng, P.; Milon, B.; Desouki, M.M.; Singh, K.K.; Kajdacsy-Balla, A.; Bagasra, O.; Costello, L.C. hZIP1 zinc uptake transporter down regulation and zinc depletion in prostate cancer. Mol. Cancer 2005, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Kong, C.; Zhang, Z.; Liu, X.; Zhan, B.; Chen, Z.; Shi, D. hZIP1 that is down-regulated in clear cell renal cell carcinoma is negatively associated with the malignant potential of the tumor. Urol. Oncol. Semin. Orig. Investig. 2014, 32, 885–892. [Google Scholar] [CrossRef]
- Guo, J.; Li, L.; Guo, B.; Liu, D.; Shi, J.; Wu, C.; Chen, J.; Zhang, X.; Wu, J. Mechanisms of resistance to chemotherapy and radiotherapy in hepatocellular carcinoma. Transl. Cancer Res. 2018, 7, 765–781. [Google Scholar] [CrossRef]
- Wang, W.-T.; Han, C.; Sun, Y.-M.; Chen, T.-Q.; Chen, Y.-Q. Noncoding RNAs in cancer therapy resistance and targeted drug development. J. Hematol. Oncol. 2019, 12, 55. [Google Scholar] [CrossRef]
- Hahne, J.C.; Valeri, N. Non-Coding RNAs and Resistance to Anticancer Drugs in Gastrointestinal Tumors. Front. Oncol. 2018, 8, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Pan, T.; Jiang, D.; Jin, L.; Geng, Y.; Feng, X.; Shen, A.; Zhang, L. The lncRNA-GAS5/miR-221-3p/DKK2 Axis Modulates ABCB1-Mediated Adriamycin Resistance of Breast Cancer via the Wnt/ß-Catenin Signaling Pathway. Mol. Ther.-Nucleic Acids 2020, 19, 1434–1448. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, L.; Li, G.; Cai, M.; Tan, C.; Han, X.; Han, L. LncRNA GAS5 confers the radio sensitivity of cervical cancer cells via regulating miR-106b/IER3 axis. Int. J. Biol. Macromol. 2019, 126, 994–1001. [Google Scholar] [CrossRef]
- Lin, J.; Liu, Z.; Liao, S.; Li, E.; Wu, X.; Zeng, W. Elevation of long non-coding RNA GAS5 and knockdown of microRNA-21 up-regulate RECK expression to enhance esophageal squamous cell carcinoma cell radio-sensitivity after radiotherapy. Genomics 2020, 112, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Meng, L.; Zhong, Y.; Hu, F.; Wang, L.; Wang, M. The long intergenic noncoding RNA GAS5 reduces cisplatin-resistance in non-small cell lung cancer through the miR-217/LHPP axis. Aging 2021, 13, 2864–2884. [Google Scholar] [CrossRef]
- Liu, B.; Wu, S.; Ma, J.; Yan, S.; Xiao, Z.; Wan, L.; Zhang, F.; Shang, M.; Mao, A. lncRNA GAS5 Reverses EMT and Tumor Stem Cell-Mediated Gemcitabine Resistance and Metastasis by Targeting miR-221/SOCS3 in Pancreatic Cancer. Mol. Ther. Nucleic Acids 2018, 13, 472–482. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer | GAS5 Expression | miRNA Expression | Mechanism of Action | Reference |
|---|---|---|---|---|
| Liver cancer | Downregulated in HepG2 and Hep3B cell lines | Overexpression of miR-21 downregulates PTEN expression in HepG2 cells | GAS5 inhibited the miR-21 expression and increased PTEN expression | [45] |
| Colorectal cancer | Downregulated in HCT116 and SW480 cell lines | miR-222-3p was upregulated in HCT116 and SW480 cell lines compared to NCM460 | GAS5 upregulate PTEN via sponging miR-222-3p | [46] |
| Papillary thyroid carcinoma | Downregulated in BHP5-16, TPC, K1, and BHP2-7 cell lines | miR-222-3p is upregulated in BHP5-16 and K1 cell lines compared to Nthy-ori 3-1 | GAS5 upregulates PTEN via sponging miR-222-3p repressing cell proliferation | [47] |
| Endometrial cancer | Downregulated in HHUA and JEC cell lines | miR-103 is upregulated in endometrial tissues | GAS5 upregulate PTEN by inhibiting miR-103 in endometrial cancer cells | [48] |
| Pancreatic cancer | Downregulated in PANC-1 and BxPC-3 cell lines | miR-32-5p is upregulated in PANC-1 and BxPC-3 cell lines compared to HPDE6-C7 | GAS5 positively regulates PTEN through sponging miR-32-5p | [49] |
| Cancer | Polymorphism | GAS5 Expression | Result | Reference |
|---|---|---|---|---|
| Acute myeloid leukemia | rs55829688 CC genotype | Elevated | Poor prognosis | [69] |
| HCC | rs145204276 with Del/Del genotype | Elevated | Increased risk of HCC | [70] |
| Oral cancer | rs145204276 Ins/Del or Del/Del genotype | Elevated | Poor cell differentiation of oral cancer, advanced tumor stage, and larger tumor size | [25] |
| Uterine cervical cancer | rs145204276 Ins/Del and Del/Del genotypes | - | low-5 year survival hazard ratio | [71] |
| Breast cancer | rs145204276 Ins/Del or Del/Del genotype | - | Decreased risk of breast cancer | [73] |
| Prostate cancer | rs145204276 Ins/Del or Del/Del genotype | - | Reduced risk of lymph node metastasis and seminal vesicle invasion | [74] |
| Osteosarcoma | rs145204276 Del/Del genotype | Elevated | Decreased risk of osteosarcoma | [27] |
| Cervical squamous cell carcinoma (CSCC) | rs145204276 Del/Del genotype | Decreased | Increased risk of CSCC | [26] |
| Urothelial cell carcinoma | rs145204276 Ins/Del or Del/Del genotype | Decreased | Associated with larger tumor size | [77] |
| Glioma | rs145204276 Ins/Del or Del/Del genotype | Decreased | Increased glioma risk | [72] |
| Bladder cancer | T G haplotype (rs2067079 and rs6790) | - | Increased risk of bladder cancer | [75] |
| Nasopharyngeal carcinoma | rs2067079 and rs6790 | - | Severe myelosuppression and severe neutropenia | [76] |
| Cancer | GAS5 Expression in Tissues/Cells | miRNA Expression in Tissues/Cells | Mechanism of Action | Reference |
|---|---|---|---|---|
| Cervical cancer | Downregulated in radio-resistant tumor tissues and SiHa cell line compared to radiosensitive tissues and ME180 cell line, respectively | miR-106b is upregulated in radioresistant tumor tissues | Overexpression GAS5 upregulates IER3 via sponging miR-106b | [94] |
| Esophageal squamous cell carcinoma | Downregulated in ESCC radiation-resistant tissues and radiation-resistant TE-1-R cell line compared to radiosensitive tissues and parent TE-1 cell line, respectively | miR-21 is upregulated in ESCC radiation-resistant tissues and radiation-resistant TE-1-R cell line compared to radiosensitive tissues and arent TE-1 cell line | GAS5 upregulates RECK expression via sponging miR-21 | [95] |
| Breast cancer | Downregulated in chemo-resistant breast cancer tissues and chemo-resistant MCF-7 cells compared chemo-sensitive breast tissues and MCF-7 chemo-sensitive cell line, respectively | miR-221-3p is upregulated in chemo-resistant breast cancer tissues and chemo-resistant MCF-7 cells compared to chemo-sensitive breast tissues and MCF-7 chemo-sensitive cell line, respectively, | GAS5 reverses the ABCB1-mediated adriamycin resistance via sponging miR-221-3p and upregulating DKK2 expression | [93] |
| Non-small cell lung cancer | Downregulated in cisplatin-resistance A549 and H1299 cell lines compared to parental cell lines | miR-217 is upregulated in negative control cisplatin-resistance A549 and H1299 cell lines compared to GAS5-overexpressed cell lines | GAS5 upregulates LHPP expression via sponging miR-217 | [96] |
| Pancreatic cancer | Downregulated in PANC-1, AsPC-1, Capan-2, SW19990, and BxPC3 pancreatic cell lines compared to normal pancreatic epithelial cells, HPDE6-C7 | miR-221 is upregulated in PANC-1, AsPC-1, Capan-2, SW19990, and BxPC3 pancreatic cell lines compared to normal pancreatic epithelial cells, HPDE6-C7 | Overexpresssion if GAS5 upregulates SOCS3 expression via sponging miR-221 | [97] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, J.; Salehen, N.; Norazit, A.; Rahman, A.A.; Murad, N.A.A.; Rahman, N.M.A.N.A.; Ibrahim, K. Tumor Suppressive Effects of GAS5 in Cancer Cells. Non-Coding RNA 2022, 8, 39. https://doi.org/10.3390/ncrna8030039
Kaur J, Salehen N, Norazit A, Rahman AA, Murad NAA, Rahman NMANA, Ibrahim K. Tumor Suppressive Effects of GAS5 in Cancer Cells. Non-Coding RNA. 2022; 8(3):39. https://doi.org/10.3390/ncrna8030039
Chicago/Turabian StyleKaur, Jesminder, Nur’ain Salehen, Anwar Norazit, Amirah Abdul Rahman, Nor Azian Abdul Murad, Nik Mohd Afizan Nik Abd. Rahman, and Kamariah Ibrahim. 2022. "Tumor Suppressive Effects of GAS5 in Cancer Cells" Non-Coding RNA 8, no. 3: 39. https://doi.org/10.3390/ncrna8030039
APA StyleKaur, J., Salehen, N., Norazit, A., Rahman, A. A., Murad, N. A. A., Rahman, N. M. A. N. A., & Ibrahim, K. (2022). Tumor Suppressive Effects of GAS5 in Cancer Cells. Non-Coding RNA, 8(3), 39. https://doi.org/10.3390/ncrna8030039