
Effects of Controlled Ozone Exposure on Circulating microRNAs and Vascular and Coagulation Biomarkers: A Mediation Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Study Participants
2.2. Study Design
2.3. Biomarker Measurement
2.3.1. miRNA Profiling
2.3.2. Protein Marker Measurement
2.4. Data Analysis
2.4.1. Identifying Differentially Expressed Circulating miRNA and Protein Biomarkers
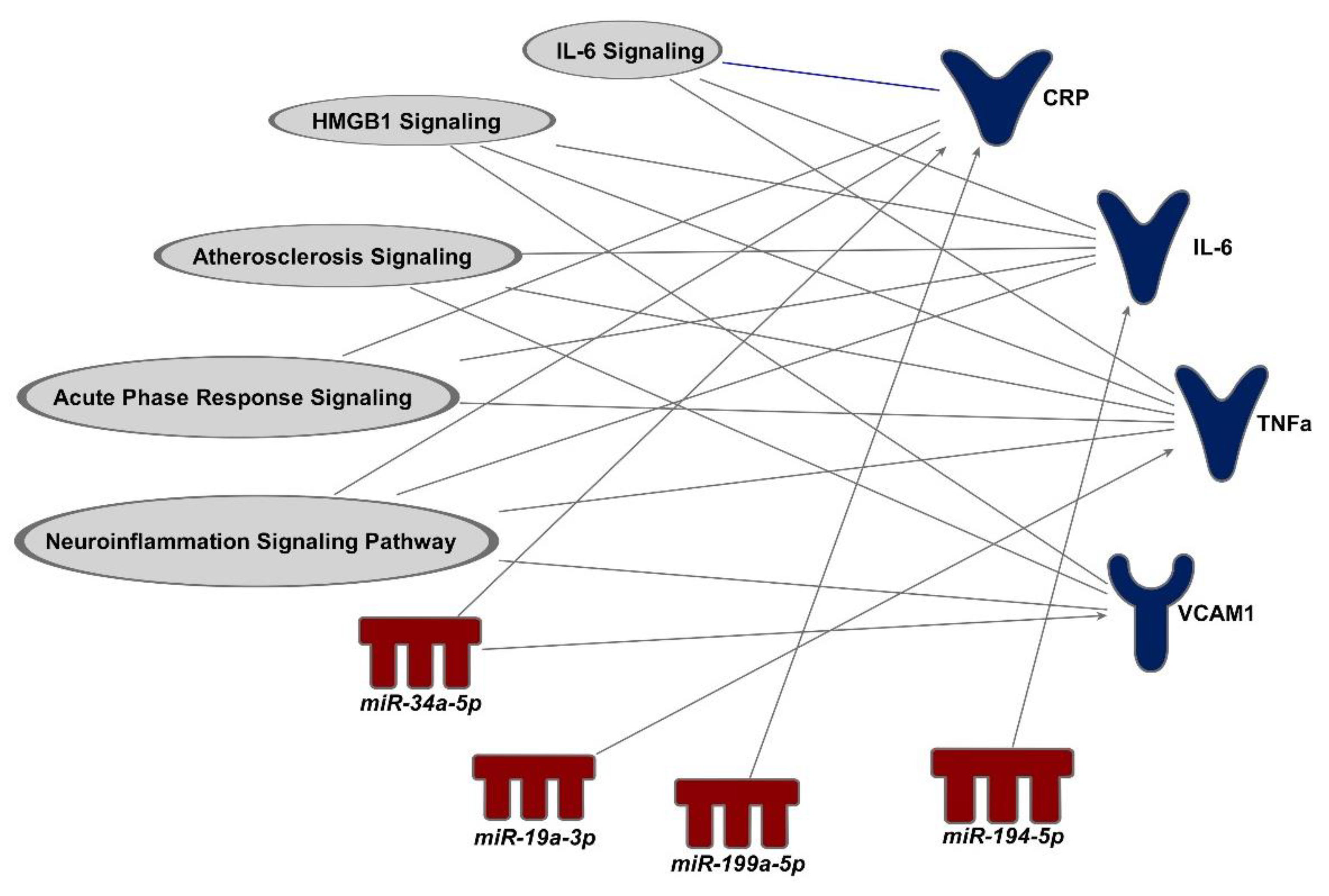
2.4.2. Functional Annotation between miRNA and Protein Biomarkers
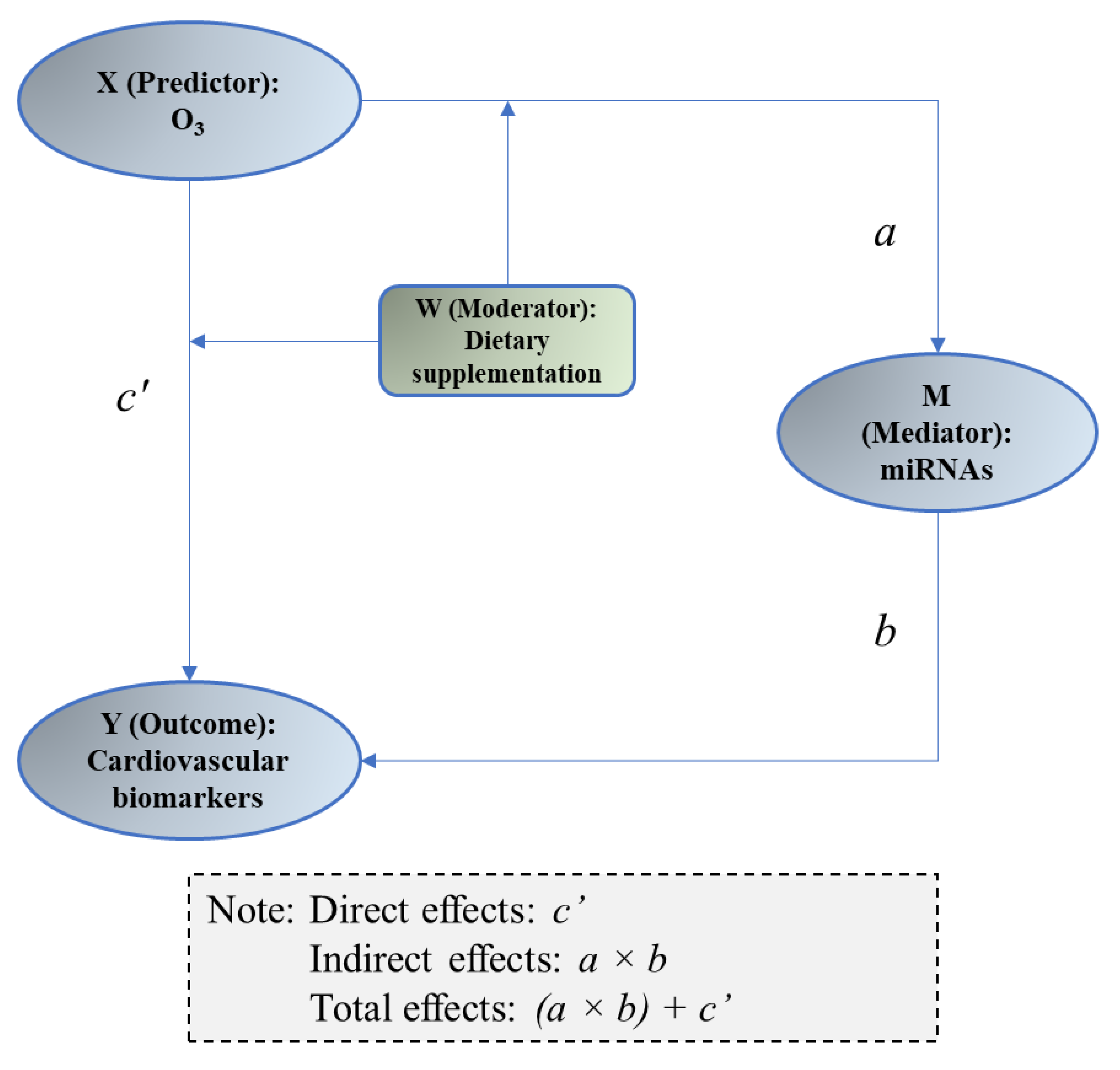
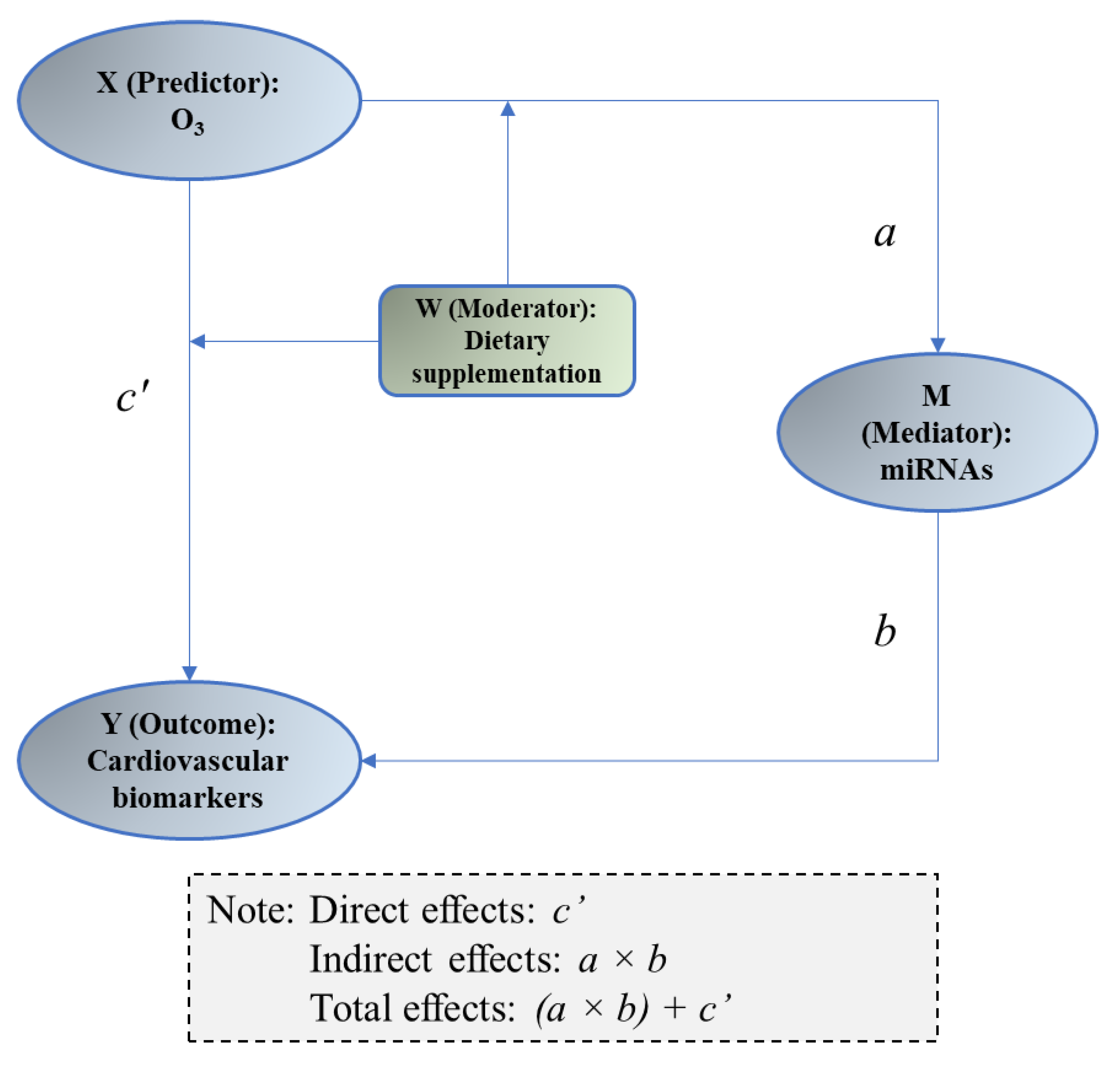
2.4.3. Mediation Analysis
3. Results
3.1. Participant Characteristics
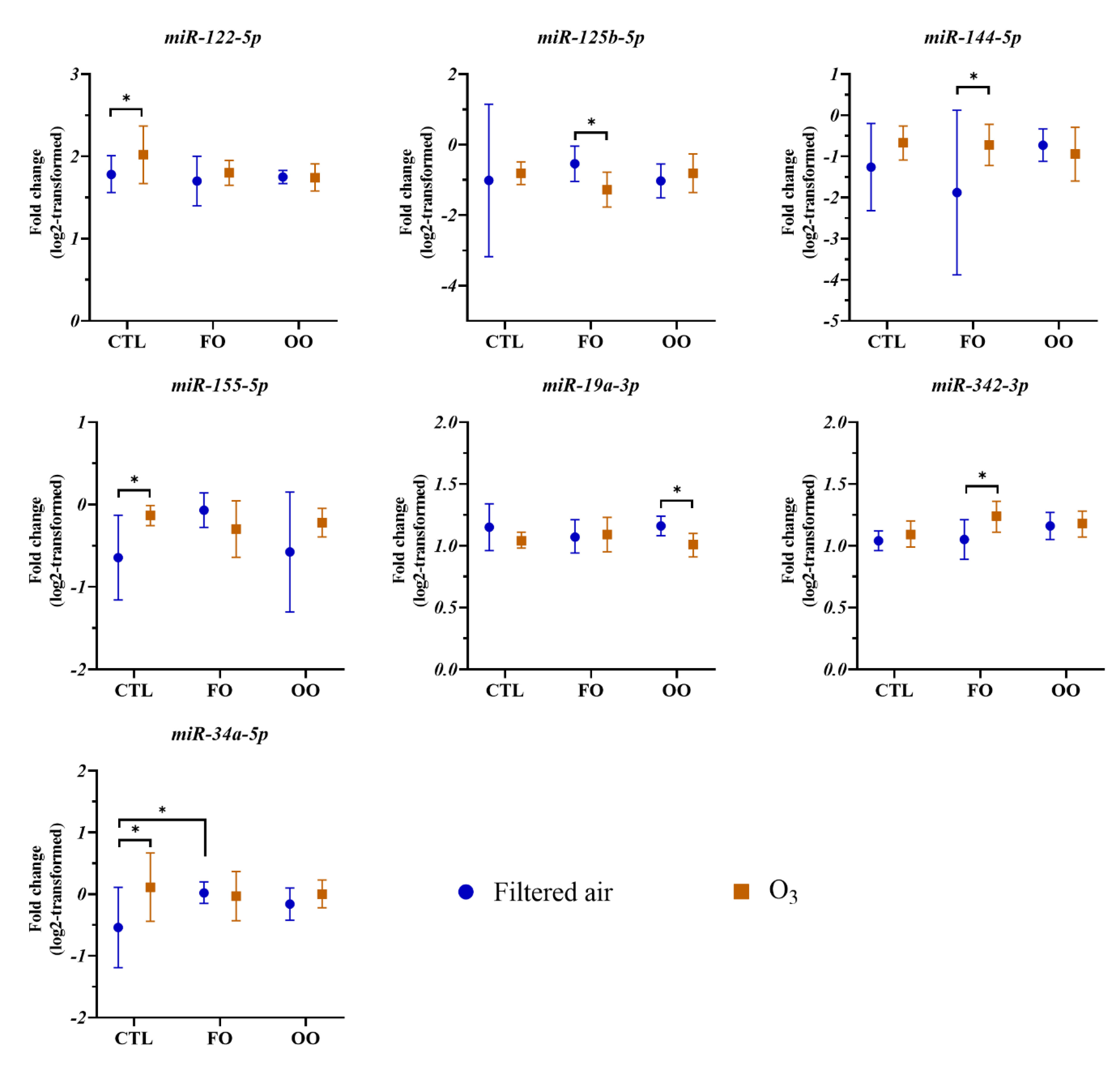
3.2. Effects of Dietary Supplementation and O3 Exposure on Circulating miRNAs
3.3. miRNAs Associated with O3 Exposure
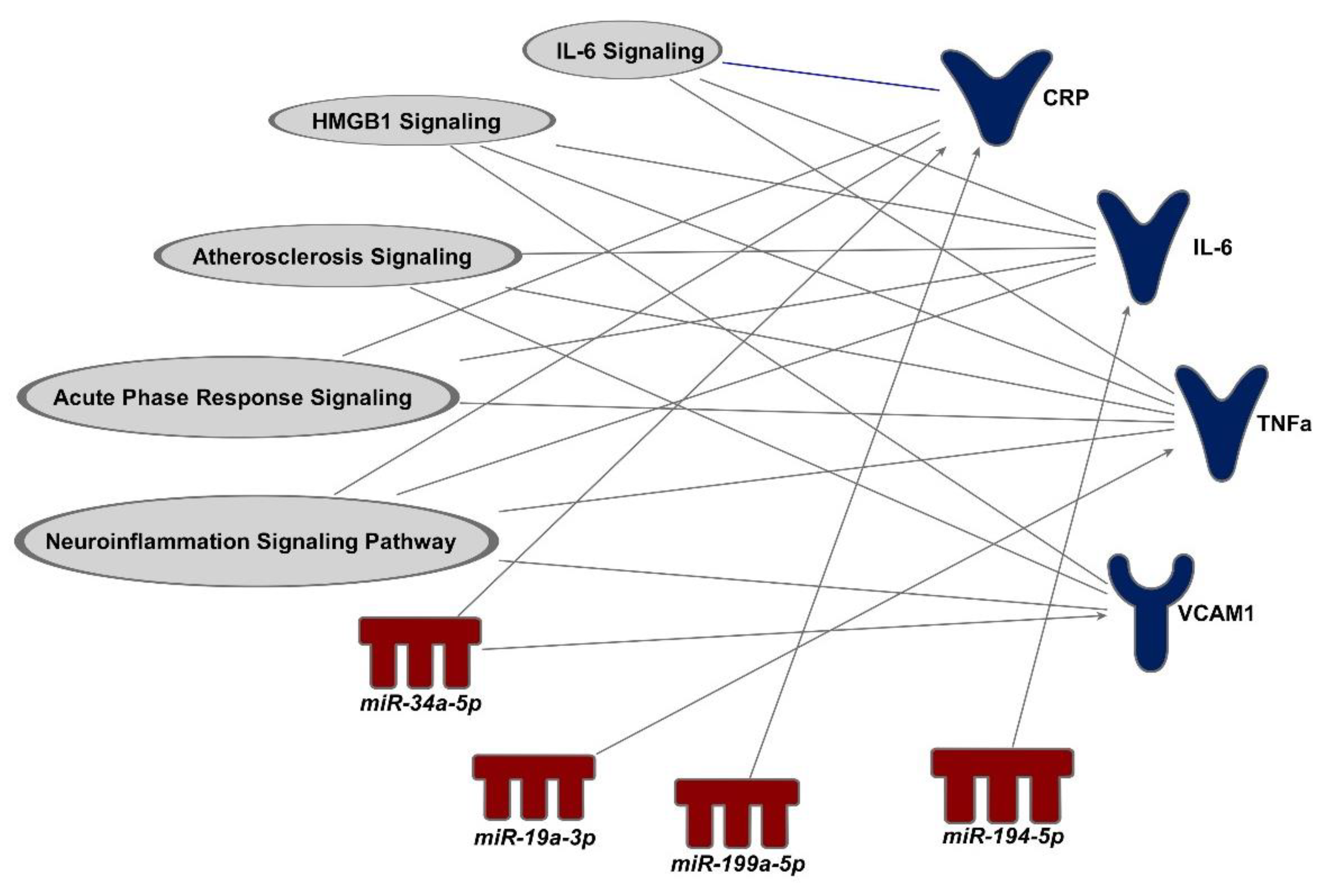
3.4. Proteins That Are Predicted to Be Downstream Biomarkers of miRNAs
3.5. Mediation of miRNAs between O3 Exposure and Changes in Vascular and Coagulation Biomarkers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EPA. Integrated Science Assessment (ISA) for Ozone and Related Photochemical Oxidants; Final Report, Feb 2013; U.S.E.P. Agency, Ed.; EPA: Washington, DC, USA, 2013.
- American Lung Association. The “State of the Air” 2021 Report. 2021. Available online: https://www.lung.org/getmedia/17c6cb6c-8a38-42a7-a3b0-6744011da370/sota-2021.pdf (accessed on 31 January 2023).
- Zhang, J.J.; Wei, Y.; Fang, Z. Ozone Pollution: A Major Health Hazard Worldwide. Front. Immunol. 2019, 10, 2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arjomandi, M.; Balmes, J.R.; Frampton, M.W.; Bromberg, P.; Rich, D.Q.; Stark, P.; Alexis, N.E.; Costantini, M.; Hollenbeck-Pringle, D.; Dagincourt, N.; et al. Respiratory Responses to Ozone Exposure. MOSES (The Multicenter Ozone Study in Older Subjects). Am. J. Respir. Crit. Care Med. 2018, 197, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Samet, J.M.; Hatch, G.E.; Horstman, D.; Steck-Scott, S.; Arab, L.; Bromberg, P.A.; Levine, M.; Mcdonnell, W.F.; Devlin, R.B. Effect of antioxidant supplementation on ozone-induced lung injury in human subjects. Am. J. Respir. Crit. Care Med. 2001, 164, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.B.; Duncan, K.E.; Jardim, M.; Schmitt, M.T.; Rappold, A.G.; Diaz-Sanchez, D. Controlled exposure of healthy young volunteers to ozone causes cardiovascular effects. Circulation 2012, 126, 104–111. [Google Scholar] [CrossRef]
- Yin, P.; Chen, R.; Wang, L.; Meng, X.; Liu, C.; Niu, Y.; Lin, Z.; Liu, Y.; Liu, J.; Qi, J.; et al. Ambient Ozone Pollution and Daily Mortality: A Nationwide Study in 272 Chinese Cities. Environ. Health Perspect. 2017, 125, 117006. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.C.; Hayes, R.B.; Ahn, J.; Shao, Y.; Silverman, D.T.; Jones, R.R.; Garcia, C.; Bell, M.L.; Thurston, G.D. Long-Term Exposure to Ozone and Cause-Specific Mortality Risk in the United States. Am. J. Respir. Crit. Care Med. 2019, 200, 1022–1031. [Google Scholar] [CrossRef]
- Bromberg, P.A. Mechanisms of the acute effects of inhaled ozone in humans. Biochim. Biophys. Acta 2016, 1860, 2771–2781. [Google Scholar] [CrossRef]
- Mudway, I.; Kelly, F. Ozone and the lung: A sensitive issue. Mol. Asp. Med. 2000, 21, 1–48. [Google Scholar] [CrossRef]
- Corteselli, E.M.; Gold, A.; Surratt, J.; Cui, T.; Bromberg, P.; Dailey, L.; Samet, J.M. Supplementation with omega-3 fatty acids potentiates oxidative stress in human airway epithelial cells exposed to ozone. Environ. Res. 2020, 187, 109627. [Google Scholar] [CrossRef]
- Devlin, R.B.; McDonnell, W.F.; Mann, R.; Becker, S.; House, D.E.; Schreinemachers, D.; Koren, H.S. Exposure of humans to ambient levels of ozone for 6.6 h causes cellular and biochemical changes in the lung. Am. J. Respir. Cell Mol. Biol. 1991, 4, 72–81. [Google Scholar] [CrossRef]
- Chen, H.; Smith, G.J.; Masood, S.; Tong, H. Extracellular MicroRNAs as Putative Biomarkers of Air Pollution Exposure, in Biomarkers in Toxicology; Patel, V.B., Preedy, V.R., Rajendram, R., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–24. [Google Scholar]
- Smith, G.J.; Tovar, A.; Kanke, M.; Wang, Y.; Deshane, J.S.; Sethupathy, P.; Kelada, S.N.P. Ozone-induced changes in the murine lung extracellular vesicle small RNA landscape. Physiol. Rep. 2021, 9, e15054. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Sun, X.; Li, L. Biogenesis and function of extracellular miRNAs. Exrna 2019, 1, 38. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xu, Y.; Rappold, A.; Diaz-Sanchez, D.; Tong, H. Effects of ambient ozone exposure on circulating extracellular vehicle microRNA levels in coronary artery disease patients. J. Toxicol. Environ. Health Part A 2020, 83, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Krauskopf, J.; Caiment, F.; van Veldhoven, K.; Chadeau-Hyam, M.; Sinharay, R.; Chung, K.F.; Cullinan, P.; Collins, P.; Barratt, B.; Kelly, F.J.; et al. The human circulating miRNome reflects multiple organ disease risks in association with short-term exposure to traffic-related air pollution. Environ. Int. 2018, 113, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Motta, V.; Favero, C.; Dioni, L.; Iodice, S.; Battaglia, C.; Angelici, L.; Vigna, L.; Pesatori, A.C.; Bollati, V. MicroRNAs are associated with blood-pressure effects of exposure to particulate matter: Results from a mediated moderation analysis. Environ. Res. 2016, 146, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Rodosthenous, R.S.; Coull, B.A.; Lu, Q.; Vokonas, P.S.; Schwartz, J.D.; Baccarelli, A.A. Ambient particulate matter and microRNAs in extracellular vesicles: A pilot study of older individuals. Part. Fibre Toxicol. 2016, 13, 13. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Li, H.; Cai, J.; Wang, C.; Lin, Z.; Liu, C.; Niu, Y.; Zhao, Z.; Li, W.; Kan, H. Fine Particulate Air Pollution and the Expression of microRNAs and Circulating Cytokines Relevant to Inflammation, Coagulation, and Vasoconstriction. Environ. Health Perspect. 2018, 126, 017007. [Google Scholar] [CrossRef] [Green Version]
- Pergoli, L.; Cantone, L.; Favero, C.; Angelici, L.; Iodice, S.; Pinatel, E.; Hoxha, M.; Dioni, L.; Letizia, M.; Albetti, B.; et al. Extracellular vesicle-packaged miRNA release after short-term exposure to particulate matter is associated with increased coagulation. Part. Fibre Toxicol. 2017, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Rodosthenous, R.S.; Kloog, I.; Colicino, E.; Zhong, J.; Herrera, L.A.; Vokonas, P.; Schwartz, J.; Baccarelli, A.A.; Prada, D. Extracellular vesicle-enriched microRNAs interact in the association between long-term particulate matter and blood pressure in elderly men. Environ. Res. 2018, 167, 640–649. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Yu, B.; Xu, Y.; Rappold, A.G.; Diaz-Sanchez, D.; Samet, J.M.; Tong, H. Circulating microRNAs as putative mediators in the association between short-term exposure to ambient air pollution and cardiovascular biomarkers. Ecotoxicol. Environ. Saf. 2022, 239, 113604. [Google Scholar] [CrossRef] [PubMed]
- VanderWeele, T.J. Mediation Analysis: A Practitioner’s Guide. Annu. Rev. Public Health 2016, 37, 17–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Tong, H.; Shen, W.; Montilla, T.S.; Case, M.W.; Almond, M.A.; Wells, H.B.; Alexis, N.E.; Peden, D.B.; Rappold, A.G.; et al. Fish oil blunts lung function decrements induced by acute exposure to ozone in young healthy adults: A randomized trial. Environ. Int. 2022, 167, 107407. [Google Scholar] [CrossRef] [PubMed]
- California Environmental Protection Agency. Environmental Protection Indicators for California, 2004 Update; California Environmental Protection Agency: Sacramento, CA, USA, 2005. [Google Scholar]
- Wang, T.; Ding, A.; Gao, J.; Wu, W.S. Strong ozone production in urban plumes from Beijing, China. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef] [Green Version]
- Kenward, M.G.; Roger, J.H. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [Green Version]
- MacKinnon, D.P.; Lockwood, C.M.; Hoffman, J.M.; West, S.G.; Sheets, V. A comparison of methods to test mediation and other intervening variable effects. Psychol. Methods 2002, 7, 83–104. [Google Scholar] [CrossRef]
- Fry, R.C.; Rager, J.E.; Bauer, R.; Sebastian, E.; Peden, D.B.; Jaspers, I.; Alexis, N.E. Air toxics and epigenetic effects: Ozone altered microRNAs in the sputum of human subjects. Am. J. Physiol. Cell. Mol. Physiol. 2014, 306, L1129–L1137. [Google Scholar] [CrossRef] [Green Version]
- Abdellatif, M. Differential expression of microRNAs in different disease states. Circ. Res. 2012, 110, 638–650. [Google Scholar] [CrossRef]
- Huang, S.; Lv, Z.; Guo, Y.; Li, L.; Zhang, Y.; Zhou, L.; Yang, B.; Wu, S.; Zhang, Y.; Xie, C.; et al. Identification of Blood Let-7e-5p as a Biomarker for Ischemic Stroke. PLoS ONE 2016, 11, e0163951. [Google Scholar] [CrossRef] [Green Version]
- Song, J.J.; Yang, M.; Liu, Y.; Song, J.W.; Wang, J.; Chi, H.J.; Liu, X.Y.; Zuo, K.; Yang, X.C.; Zhong, J.C. MicroRNA-122 aggravates angiotensin II-mediated apoptosis and autophagy imbalance in rat aortic adventitial fibroblasts via the modulation of SIRT6-elabela-ACE2 signaling. Eur. J. Pharmacol. 2020, 883, 173374. [Google Scholar] [CrossRef]
- Scrutinio, D.; Conserva, F.; Passantino, A.; Iacoviello, M.; Lagioia, R.; Gesualdo, L. Circulating microRNA-150-5p as a novel biomarker for advanced heart failure: A genome-wide prospective study. J. Heart Lung Transplant. 2017, 36, 616–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhang, Y. miR-342-3p affects hepatocellular carcinoma cell proliferation via regulating NF-kappaB pathway. Biochem. Biophys. Res. Commun. 2015, 457, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Li, Y.; Wu, Q.; Huang, L.; Xu, J.; Zeng, Q. Pathogenic role of microRNAs in atherosclerotic ischemic stroke: Implications for diagnosis and therapy. Genes Dis. 2022, 9, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Raucci, A.; Macrì, F.; Castiglione, S.; Badi, I.; Vinci, M.C.; Zuccolo, E. MicroRNA-34a: The bad guy in age-related vascular diseases. Cell. Mol. Life Sci. 2021, 78, 7355–7378. [Google Scholar] [CrossRef]
- Su, Y.; Sun, Y.; Tang, Y.; Li, H.; Wang, X.; Pan, X.; Liu, W.; Zhang, X.; Zhang, F.; Xu, Y.; et al. Circulating miR-19b-3p as a Novel Prognostic Biomarker for Acute Heart Failure. J. Am. Heart Assoc. 2021, 10, e022304. [Google Scholar] [CrossRef]
- Chen, H.; Li, X.; Liu, S.; Gu, L.; Zhou, X. MircroRNA-19a promotes vascular inflammation and foam cell formation by targeting HBP-1 in atherogenesis. Sci. Rep. 2017, 7, 12089. [Google Scholar] [CrossRef]
- Kim, T.; Valera, E.; Desplats, P. Alterations in Striatal microRNA-mRNA Networks Contribute to Neuroinflammation in Multiple System Atrophy. Mol. Neurobiol. 2019, 56, 7003–7021. [Google Scholar] [CrossRef]
- Tong, H.; Zhang, S.; Shen, W.; Chen, H.; Salazar, C.; Schneider, A.; Rappold, A.G.; Diaz-Sanchez, D.; Devlin, R.B.; Samet, J.M. Lung Function and Short-Term Ambient Air Pollution Exposure: Differential Impacts of Omega-3 and Omega-6 Fatty Acids. Ann. Am. Thorac. Soc. 2022, 19, 583–593. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Shen, W.; Salazar, C.; Schneider, A.; Wyatt, L.H.; Rappold, A.G.; Diaz-Sanchez, D.; Devlin, R.B.; Samet, J.M.; et al. Omega-3 fatty acids attenuate cardiovascular effects of short-term exposure to ambient air pollution. Part. Fibre Toxicol. 2022, 19, 12. [Google Scholar] [CrossRef]
- Albert, B.B.; Cameron-Smith, D.; Hofman, P.L.; Cutfield, W.S. Oxidation of Marine Omega-3 Supplements and Human Health. BioMed Res. Int. 2013, 2013, 464921. [Google Scholar] [CrossRef] [Green Version]
- Perona, J.; Cabellomoruno, R.; Ruizgutierrez, V. The role of virgin olive oil components in the modulation of endothelial function. J. Nutr. Biochem. 2006, 17, 429–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Li, M.; Geng, F. Omega-3 Polyunsaturated Fatty Acids Eicosapentaenoic Acid and Docosahexaenoic Acid Enhance Dexamethasone Sensitivity in Multiple Myeloma Cells by the p53/miR-34a/Bcl-2 Axis. Biochemistry 2017, 82, 826–833. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Characteristics | CTL (n = 6) | FO (n = 7) | OO (n = 10) | All (n = 23) |
| Age (years) | 23.5 (3.7) | 27.4 (4.7) | 26.5 (3.0) | 26.0 (3.9) |
| BMI (kg/m2) | 24.9 (3.6) | 25.8 (3.8) | 25.1 (2.7) | 25.3 (3.2) |
| Omega-3 index (%) | 4.0 (0.2) | 6.1 (1.2) | 4.0 (0.4) | 4.6 (1.2) |
| Race/ethnicity (no. of participants) | ||||
| African-American | 0 | 2 | 0 | 2 |
| Asian | 1 | 1 | 0 | 2 |
| Caucasian | 3 | 4 | 10 | 17 |
| Hispanic | 2 | 0 | 0 | 2 |
| miRNAs | F Value | p | Predicted Targets |
|---|---|---|---|
| miR-122-5p | 7.09 | 0.015 * | - |
| miR-144-5p | 6.39 | 0.0448 * | - |
| miR-192-5p | 4.37 | 0.0496 * | - |
| miR-194-5p | 7.83 | 0.0111 * | D-dimer |
| miR-199a-5p | 7.14 | 0.0146 * | CRP |
| miR-19a-3p | 5.47 | 0.0299 * | TNFα |
| miR-34a-5p | 4.77 | 0.0441 * | CRP, sVCAM1 |
| miRNAs (M) | Biomarker (Y) | X → M (a) | M → Y (b) | Direct Effects (c′) | Indirect Effects (a × b) | ||
|---|---|---|---|---|---|---|---|
| Coefficient (95%CI) | Proportion (%) | p | |||||
| miR-194-5p | D-dimer | 0.45 (−0.2, 1.1) | 0.16 (0.09, 0.24) | −0.17 (−0.29, −0.05) | 0.07 (−0.04, 0.19) | −76.78 | 0.20 |
| miR-199a-5p | CRP | −0.38 (−1.07, 0.31) | 0.02 (−0.05, 0.09) | 0.34 (0.22, 0.46) | −0.01 (−0.04, 0.02) | −2.17 | 0.64 |
| miR-19a-3p | TNFα | −1.03 (−1.81, −0.25) | −0.08 (−0.11, −0.05) | −0.21 (−0.28, −0.13) | 0.08 (0.01, 0.15) | −67.09 | 0.02 * |
| miR-34a-5p | CRP | 0.32 (−0.53, 1.18) | 0.22 (0.16, 0.27) | 0.27 (0.16, 0.37) | 0.07 (−0.11, 0.25) | 20.79 | 0.46 |
| miR-34a-5p | sVCAM1 | 0.32 (−0.53, 1.18) | 0.05 (0.02, 0.08) | 0.05 (−0.01, 0.11) | 0.02 (−0.03, 0.06) | 25.63 | 0.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Masood, S.; Rappold, A.G.; Diaz-Sanchez, D.; Samet, J.M.; Tong, H. Effects of Controlled Ozone Exposure on Circulating microRNAs and Vascular and Coagulation Biomarkers: A Mediation Analysis. Non-Coding RNA 2023, 9, 43. https://doi.org/10.3390/ncrna9040043
Chen H, Masood S, Rappold AG, Diaz-Sanchez D, Samet JM, Tong H. Effects of Controlled Ozone Exposure on Circulating microRNAs and Vascular and Coagulation Biomarkers: A Mediation Analysis. Non-Coding RNA. 2023; 9(4):43. https://doi.org/10.3390/ncrna9040043
Chicago/Turabian StyleChen, Hao, Syed Masood, Ana G. Rappold, David Diaz-Sanchez, James M. Samet, and Haiyan Tong. 2023. "Effects of Controlled Ozone Exposure on Circulating microRNAs and Vascular and Coagulation Biomarkers: A Mediation Analysis" Non-Coding RNA 9, no. 4: 43. https://doi.org/10.3390/ncrna9040043
APA StyleChen, H., Masood, S., Rappold, A. G., Diaz-Sanchez, D., Samet, J. M., & Tong, H. (2023). Effects of Controlled Ozone Exposure on Circulating microRNAs and Vascular and Coagulation Biomarkers: A Mediation Analysis. Non-Coding RNA, 9(4), 43. https://doi.org/10.3390/ncrna9040043