
Novel Insights into Circular RNAs in Metastasis in Breast Cancer: An Update




Abstract
:1. Introduction
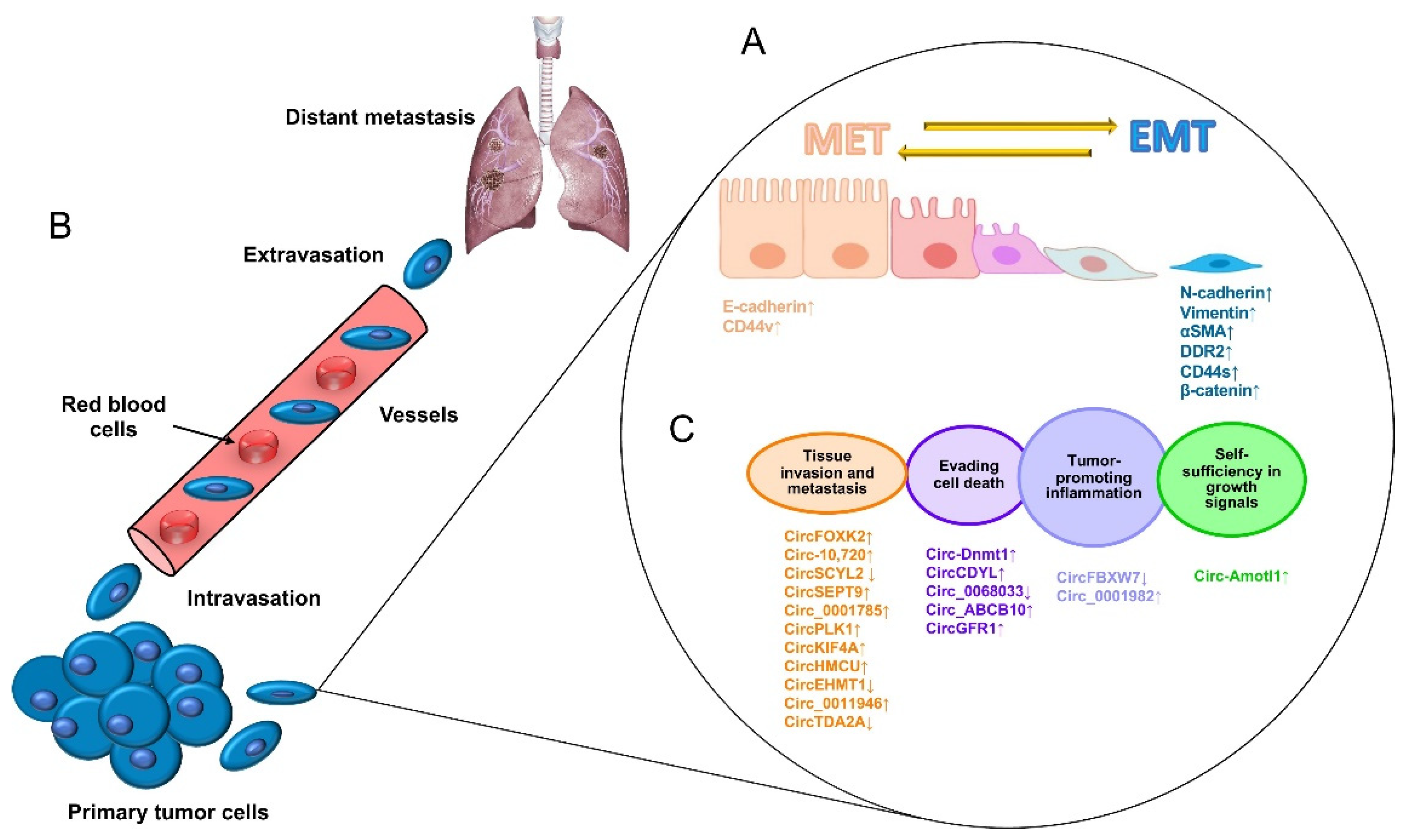
2. Metastasis in Breast Cancer: A Challenging Hallmark
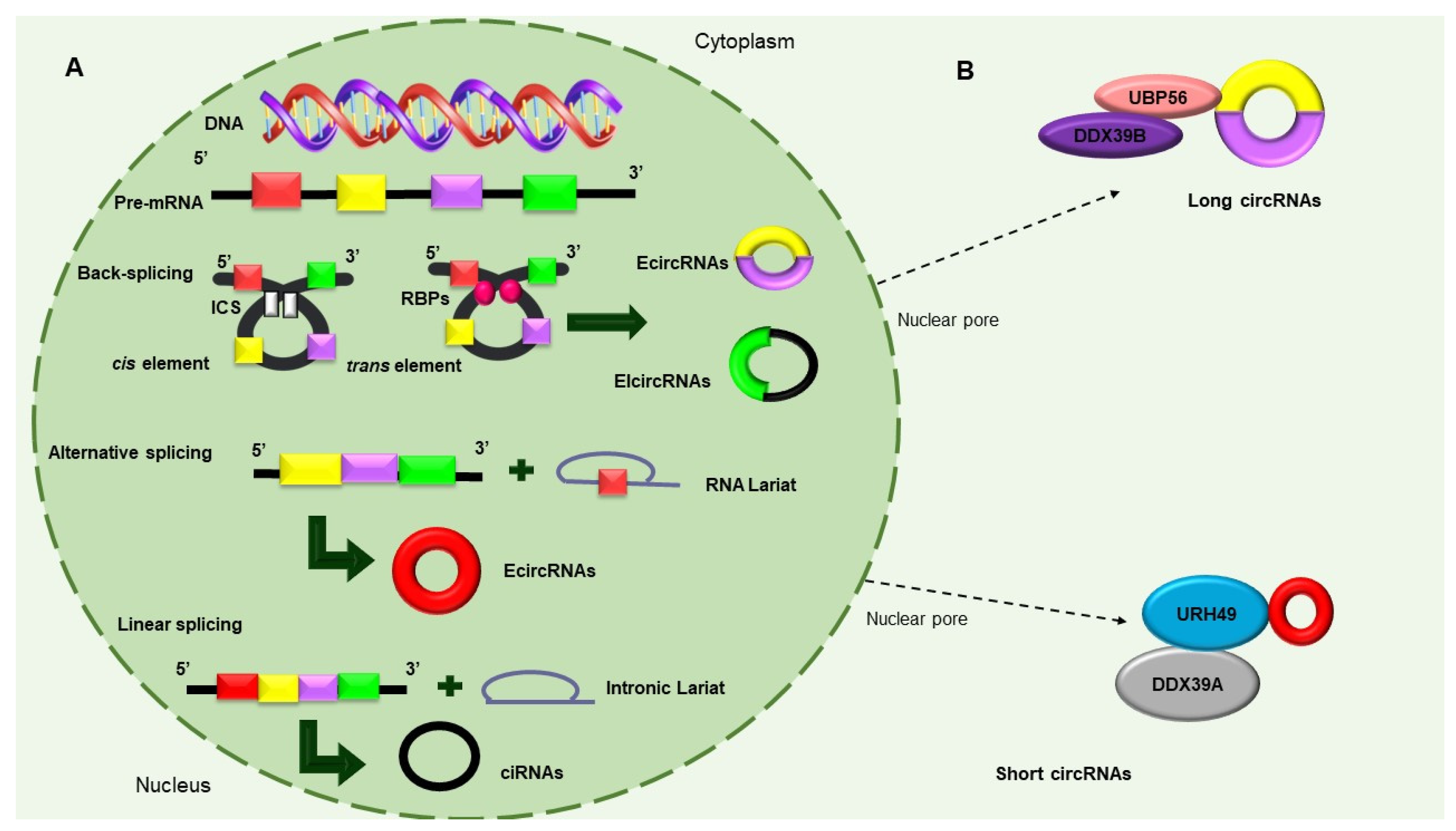
3. Circular RNAs: New Players in Gene Expression
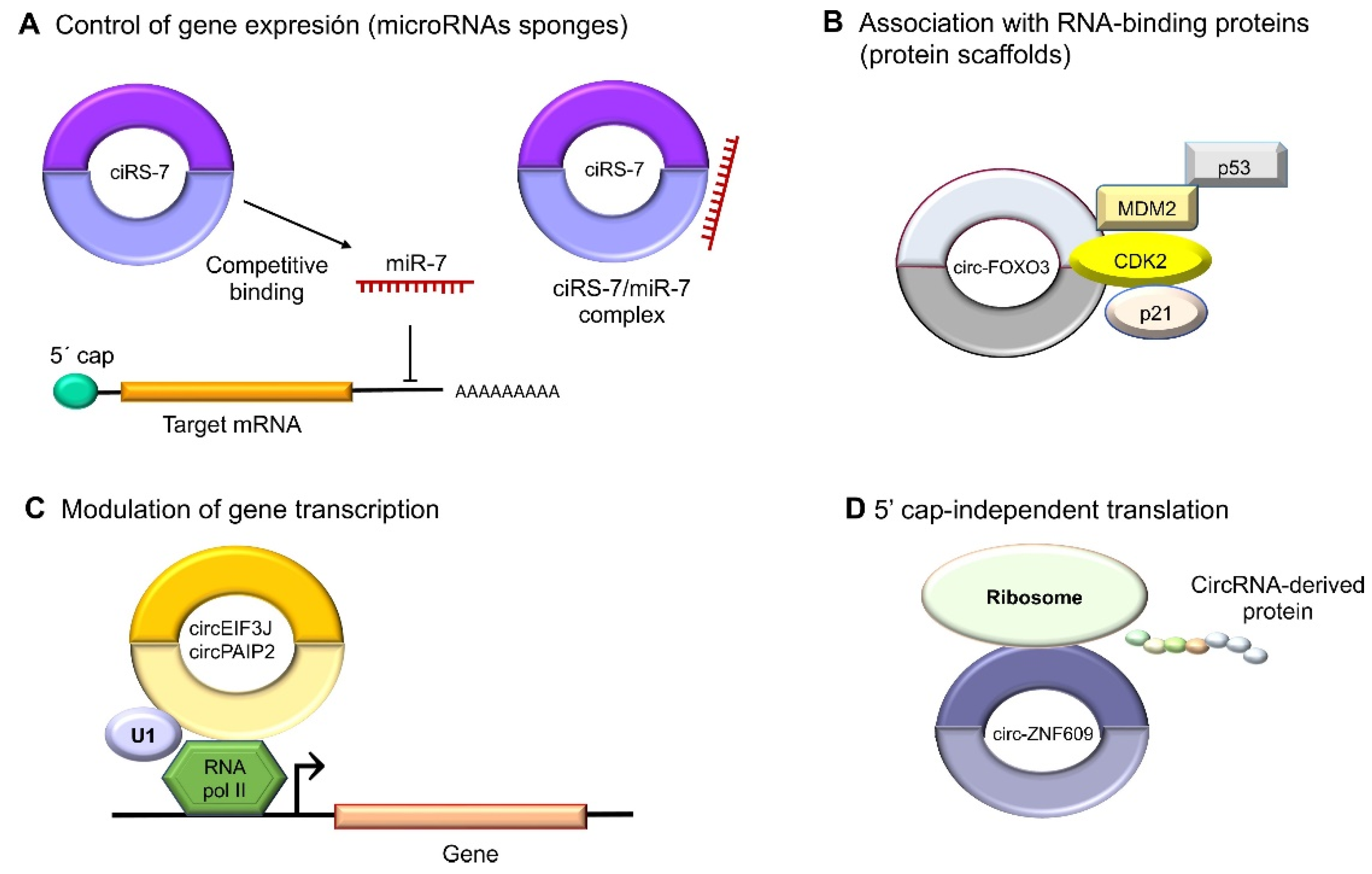
4. Roles of Circular RNAs in Cancer Cells
Functions in RNA Polymerase II Transcription and Splicing
5. Circular RNAs Functions in Triple-Negative Breast Cancer
CircRNAs Sponge Relevant MicroRNAs Involved in Tumor Progression and Metastasis
6. Circular RNAs Exported in Exosomes in TNBC
7. Circular RNAs Regulation of Resistance to Chemo/Radiotherapy
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Schottenfeld, D.; Fraumeni, J.F., Jr. Cancer Epidemiology and Prevention, 4th ed.; Oxford University Press: Oxford, UK, 2018; pp. 107–147. [Google Scholar]
- Harbeck, N.; Gnant, M. Breast cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef]
- Zubair, M.; Wang, S.; Ali, N. Advanced approaches to breast cancer classification and diagnosis. Front. Pharmacol. 2021, 11, 2487. [Google Scholar] [CrossRef] [PubMed]
- McDonald, E.S.; Clark, A.S.; Tchou, J.; Zhang, P.; Freedman, G.M. Clinical Diagnosis and Management of Breast Cancer. J. Am. Coll. Cardiol. 2016, 57, 9S–16S. [Google Scholar] [CrossRef] [PubMed]
- Maughan, K.L.; Lutterbie, M.A.; Ham, P.S. Treatment of breast cancer. Am. Fam. Physician. 2010, 81, 1339–1346. [Google Scholar] [PubMed]
- Howard, F.M.; Olopade, O.I. Epidemiology of Triple-Negative Breast Cancer: A Review. Cancer J. 2021, 27, 8–16. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef]
- Dass, S.A.; Tan, K.L.; Selva Rajan, R.; Mokhtar, N.F.; Mohd Adzmi, E.R.; Wan Abdul Rahman, W.F.; Tengku Din, T.; Bala-krishnan, V. Triple Negative Breast Cancer: A Review of Present and Future Diagnostic Modalities. Medicina 2021, 57, 62. [Google Scholar] [CrossRef]
- Herschkowitz, J.I.; Simin, K.; Weigman, V.J.; Mikaelian, I.; Usary, J.; Hu, Z.; Perou, C.M. Identification of conserved gene expression features between murine mammary carcinoma models and human breast tumors. Genome Biol. 2007, 8, 1–17. [Google Scholar] [CrossRef]
- Francis, I.M.; Altemaimi, R.A.; Al-Ayadhy, B.; Alath, P.; Jaragh, M.; Mothafar, F.J.; Kapila, K. Hormone receptors and human epidermal growth factor (HER2) expression in fine-needle aspirates from metastatic breast carcinoma–Role in patient management. J. Cytol. 2019, 36, 94. [Google Scholar] [PubMed]
- Marrugo-Ramírez, J.; Mir, M.; Samitier, J. Blood-based cancer biomarkers in liquid biopsy: A promising non-invasive alter-native to tissue biopsy. Int. J. Mol. Sci. 2018, 19, 2877. [Google Scholar] [CrossRef] [PubMed]
- Alimirzaie, S.; Bagherzadeh, M.; Akbari, M.R. Liquid biopsy in breast cancer: A comprehensive review. Clin. Genet. 2019, 95, 643–660. [Google Scholar] [CrossRef]
- Bergin, A.; Loi, S. Triple-negative breast cancer: Recent treatment advances. F1000Research 2019, 8, F1000. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Oncology meets immunology: The cancer-immunity cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Hu, L.; Zhang, X.; Jiang, S.; Li, J.; Zhang, Z.; Wang, X. The Diverse Function of PD-1/PD-L Pathway Beyond Cancer. Front. Immunol. 2019, 10, 2298. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Liu, N.; Yang, L.; Tang, J.; Wang, Y.; Mei, M. A Brief Review of circRNA Biogenesis, Detection, and Function. Curr. Genom. 2021, 22, 485–495. [Google Scholar] [CrossRef]
- Nguyen, D.X.; Bos, P.D.; Massagué, J. Metastasis: From dissemination to organ-specific colonization. Nat. Rev. Cancer. 2009, 9, 274–284. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, H.; Song, X.; Yang, Q. Metastatic heterogeneity of breast cancer: Molecular mechanism and potential ther-apeutic targets. Semin. Cancer Biol. 2020, 60, 14–27. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, Y.; Dai, H.; Han, B. Epithelial-Mesenchymal Transition-Mediated Tumor Therapeutic Resistance. Molecules 2022, 27, 4750. [Google Scholar] [CrossRef]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29 (Suppl. S16), 15–18. [Google Scholar] [CrossRef] [PubMed]
- Turai, P.I.; Gábor, N.; Henriett, B.; Attila, P.; Peter, I. MicroRNAs, long non-coding RNAs, and circular RNAs: Potential biomarkers and therapeutic targets in pheochromocytoma/paraganglioma. Cancers 2021, 13, 1522. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.K.; Mitchell, G. Regulatory non-coding RNAs: Everything is possible, but what is important? Nat. Methods 2022, 19, 1156–1159. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, R.; Singh, A.K.; Mishra, G.; Maurya, A.; Chellappan, D.K.; Gupta, G.; Hansbro, P.M.; Dua, K. An Overview of Circular RNAs. Adv. Exp. Med. Biol. 2018, 1087, 3–14. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Li, Z.; Kearse, M.G.; Huang, C. The nuclear export of circular RNAs is primarily defined by their length. RNA Biol. 2019, 16, 1–4. [Google Scholar] [CrossRef]
- Zhou, M.; Xiao, M.S.; Li, Z.; Huang, C. New progresses of circular RNA biology: From nuclear export to degradation. RNA Biol. 2021, 18, 1365–1373. [Google Scholar] [CrossRef]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE. 2012, 7, e30733. [Google Scholar] [CrossRef]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- Khan, F.A.; Nsengimana, B.; Khan, N.H.; Song, Z.; Ngowi, E.E.; Wang, Y.; Zhang, W.; Ji, S. Chimeric Peptides/Proteins Encoded by circRNA: An Update on Mechanisms and Functions in Human Cancers. Front. Oncol. 2022, 12, 781270. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, J.; Tian, Y.; Gao, Y.; Dong, X.; Chen, W.; Yuan, X.; Yin, W.; Xu, J.; Chen, K.; et al. CircRNA inhibits DNA damage repair by interacting with host gene. Mol. Cancer 2020, 19, 128. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in human diseases and potential use as biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Lu, J.Y.; Zhang, X.; Shao, W.; Xu, Y.; Li, P.; Hong, Y.; Cui, L.; Shan, G.; Tian, B.; et al. U1 snRNP regulates chromatin retention of noncoding RNAs. Nature 2020, 580, 147–150. [Google Scholar] [CrossRef]
- Qin, M.; Wei, G.; Sun, X. Circ-UBR5: An exonic circular RNA and novel small nuclear RNA involved in RNA splicing. Biochem. Biophys. Res. Commun. 2018, 503, 1027–1034. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell. 2017, 66, 22–37. [Google Scholar] [CrossRef]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, S.; Liao, X.; Li, M.; Chen, S.; Li, X.; Wu, X.; Yang, M.; Tang, M.; Hu, Y.; et al. Circular RNA circIKBKB promotes breast cancer bone metastasis through sustaining NF-κB/bone remodeling factors signaling. Mol. Cancer 2021, 20, 98. [Google Scholar] [CrossRef]
- Fu, B.; Liu, W.; Zhu, C.; Li, P.; Wang, L.; Pan, L.; Li, K.; Cai, P.; Meng, M.; Wang, Y.; et al. Circular RNA circBCBM1 promotes breast cancer brain metastasis by modulating miR-125a/BRD4 axis. Int. J. Biol. Sci. 2021, 17, 3104–3117. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Zhang, Y.; Wang, X.; Xue, B.; Sun, M.; Li, H. Circular RNA circ-ZEB1 acts as an oncogene in triple negative breast cancer via sponging miR-448. Int. J. Cell Biol. 2020, 126, 105798. [Google Scholar] [CrossRef]
- Du, W.W.; Li, X.; Ma, J.; Fang, L.; Wu, N.; Li, F.; Dhaliwal, P.; Yang, W.; Yee, A.J.; Yang, B.B. Promotion of tumor progression by exosome transmission of circular RNA circSKA3. Mol. Ther. Nucleic Acids. 2022, 27, 276–292. [Google Scholar] [CrossRef]
- Xiang, Z.; Xu, C.; Wu, G.; Liu, B.; Wu, D. CircRNA-UCK2 increased TET1 inhibits proliferation and invasion of prostate cancer cells via sponge MiRNA-767-5p. Open Med. 2019, 14, 833–842. [Google Scholar] [CrossRef]
- Xing, Z.; Wang, R.; Wang, X.; Liu, J.; Zhang, M.; Feng, K.; Wang, X. CircRNA circ-PDCD11 promotes triple-negative breast cancer progression via enhancing aerobic glycolysis. Cell Death Discov. 2021, 7, 218. [Google Scholar] [CrossRef] [PubMed]
- Lyu, L.; Zhang, S.; Deng, Y.; Wang, M.; Deng, X.; Yang, S.; Wu, Y.; Dai, Z. Regulatory mechanisms, functions, and clinical significance of CircRNAs in triple-negative breast cancer. J. Hematol. Oncol. 2021, 14, 41. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.R.; Huang, Q.D.; Zheng, Z.H.; Zhang, Z.T.; Guo, G.L. CircGFRA1 affects the sensitivity of triple-negative breast cancer cells to paclitaxel via the miR-361-5p/TLR4 pathway. J. Biochem. 2021, 169, 601–611. [Google Scholar] [CrossRef]
- Chen, B.; Wei, W.; Huang, X.; Xie, X.; Kong, Y.; Dai, D.; Yang, L.; Wang, J.; Tang, H.; Xie, X. CircEPSTI1 as a Prognostic Marker and Mediator of Triple-Negative Breast Cancer Progression. Theranostics 2018, 8, 4003–4015. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, X.; Li, C.; Zhang, H.; Liu, Y.; Han, D.; Li, Y.; Li, Z.; Luo, D.; Zhang, N.; et al. CircHIF1A regulated by FUS accelerates triple-negative breast cancer progression by modulating NFIB expression and translocation. Oncogene 2021, 40, 2756–2771. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhou, S.; Xu, H.; Wang, D.; Feng, J.; Zhao, J.; Zhong, S. Circular RNA expression in exosomes derived from breast cancer cells and patients. Epigenomics 2019, 11, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xue, B.; Zhang, Y.; Guo, G.; Duan, X.; Dou, D. Up-regulated circBACH2 contributes to cell proliferation, invasion, and migration of triple-negative breast cancer. Cell Death Dis. 2021, 12, 412. [Google Scholar] [CrossRef]
- Zheng, X.; Huang, M.; Xing, L.; Yang, R.; Wang, X.; Jiang, R.; Zhang, L.; Chen, J. The circRNA circSEPT9 mediated by E2F1 and EIF4A3 facilitates the carcinogenesis and development of triple-negative breast cancer. Mol. Cancer 2020, 19, 73. [Google Scholar] [CrossRef]
- Yang, R.; Xing, L.; Zheng, X.; Sun, Y.; Wang, X.; Chen, J. The circRNA circAGFG1 acts as a sponge of miR-195-5p to promote triple-negative breast cancer progression through regulating CCNE1 expression. Mol. Cancer 2019, 18, 4. [Google Scholar] [CrossRef]
- Li, J.; Ma, M.; Yang, X.; Zhang, M.; Luo, J.; Zhou, H.; Huang, N.; Xiao, F.; Lai, B.; Lv, W.; et al. Circular HER2 RNA positive triple negative breast cancer is sensitive to Pertuzumab. Mol. Cancer 2020, 19, 142. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, X.; Lan, J.; Wan, Y.; Zhu, X. Circular RNA circRPPH1 promotes triple-negative breast cancer progression via the miR-556-5p/YAP1 axis. Am. J. Transl. Res. 2020, 12, 6220–6234. [Google Scholar] [PubMed]
- Li, Y.; Shi, P.; Zheng, T.; Ying, Z.; Jiang, D. Circular RNA hsa_circ_0131242 Promotes Triple-Negative Breast Cancer Progression by Sponging hsa-miR-2682. Onco. Targets Ther. 2020, 13, 4791–4798. [Google Scholar] [CrossRef]
- Zhou, Y.; Ma, G.; Peng, S.; Tuo, M.; Li, Y.; Qin, X.; Yu, Q.; Kuang, S.; Cheng, H.; Li, J. Circ_0000520 contributes to triple-negative breast cancer progression through mediating the miR-1296/ZFX axis. Thorac. Cancer 2021, 12, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zou, Y.; Li, X.; Yang, A.; Ye, F.; Zhang, J.; Wei, W.; Kong, Y. CircGNB1 Facilitates Triple-Negative Breast Cancer Pro-gression by Regulating miR-141-5p-IGF1R Axis. Front. Genet. 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Shen, W.; Xu, J.; Gong, B.; Gao, B.; Zhu, J. CircUSP42 is downregulated in triple-negative breast cancer and associated with poor prognosis. Technol. Cancer Res. Treat. 2020, 19, 1533033820950827. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, F.; Ma, H.; Cui, X.; Yang, S.; Qin, R. CircCDYL Acts as a Tumor Suppressor in Triple Negative Breast Cancer by Sponging miR-190a-3p and Upregulating TP53INP1. Clin. Breast Cancer 2020, 20, 422–430. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, J.; Jin, W.; Sun, Y.; Xu, Y.; Wang, Y.; Liang, X.; Su, D. CircNR3C2 promotes HRD1-mediated tumor-suppressive effect via sponging miR-513a-3p in triple-negative breast cancer. Mol. Cancer 2021, 20, 25. [Google Scholar] [CrossRef]
- Xiao, W.; Zheng, S.; Zou, Y.; Yang, A.; Xie, X.; Tang, H.; Xie, X. CircAHNAK1 inhibits proliferation and metastasis of tri-ple-negative breast cancer by modulating miR-421 and RASA1. Aging 2019, 11, 12043–12056. [Google Scholar] [CrossRef]
- Hu, J.; Ji, C.; Hua, K.; Wang, X.; Deng, X.; Li, J.; Graham, D.; Fang, L. Hsa_circ_0091074 regulates TAZ expression via mi-croRNA-1297 in triple negative breast cancer cells. Int. J. Oncol. 2020, 56, 1314–1326. [Google Scholar] [CrossRef] [PubMed]
- Dou, D.; Ren, X.; Han, M.; Xu, X.; Ge, X.; Gu, Y.; Wang, X.; Zhao, S. CircUBE2D2 (has_circ_0005728) promotes cell proliferation, metastasis and chemoresistance in triple-negative breast cancer by regulating miR-512-3p/CDCA3 axis. Cancer Cell Int. 2020, 20, 454. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liang, D.; Shen, P.; Yu, Y.; Yan, Y.; You, W. Hsa_circ_0092276 promotes doxorubicin resistance in breast cancer cells by regulating autophagy via miR-348/ATG7 axis. Transl. Oncol. 2021, 14, 101045. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Song, X.; Li, Y.; Su, P.; Han, D.; Ma, T.; Guo, R.; Chen, B.; Zhao, W.; Sang, Y.; et al. Correction: CircKDM4C suppresses tumor progression and attenuates doxorubicin resistance by regulating miR-548p/PBLD axis in breast cancer. Oncogene 2021, 40, 2816. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wan, R. The Emerging Roles of circSMARCA5 in Cancer. J. Oncol. 2022, 7, 3015818. [Google Scholar] [CrossRef]
- Zeng, K.; He, B.; Yang, B.B.; Xu, T.; Chen, X.; Xu, M.; Liu, X.; Sun, H.; Pan, Y.; Wang, S. The pro-metastasis effect of circANKS1B in breast cancer. Mol. Cancer 2018, 17, 160. [Google Scholar] [CrossRef] [PubMed]
- Jadlii, A.S.; Ballasy, N.; Edalat, P.; Patel, V.B. Inside(sight) of tiny communicator: Exosome biogenesis, secretion, and uptake. Mol. Cellular Biochem. 2020, 467, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Vietri, M.; Radulovic, M.; Stenmark, H. The many functions of ESCRTs. Nat. Rev. Mol. Cell Biol. 2020, 21, 25–42. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef]
- Yang, S.J.; Wang, D.D.; Zhong, S.L.; Chen, W.Q.; Wang, F.L.; Zhang, J.; Xu, W.X.; Xu, D.; Zhang, Q.; Li, J.; et al. Tumor-derived exosomal circPSMA1 facilitates the tumorigenesis, metastasis, and migration in triple-negative breast cancer (TNBC) through miR-637/Akt1/β-catenin (cyclin D1) axis. Cell Dis. 2021, 12, 420. [Google Scholar] [CrossRef]
- Zhan, Y.; Du, J.; Min, Z.; Ma, L.; Zhang, W.; Zhu, W.; Liu, Y. Carcinoma-associated fibroblasts derived exosomes modulate breast cancer cell stemness through exonic circHIF1A by miR-580-5p in hypoxic stress. Cell Death Discov. 2021, 7, 141. [Google Scholar] [CrossRef]
- Chen, D.R.; Lu, D.Y.; Lin, H.Y.; Yeh, W.L. Mesenchymal stem cell-induced doxorubicin resistance in triple negative breast cancer. Biomed. Res. Int. 2014, 2014, 532161. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Fang, L.; Yang, Q.; Hibberd, S.; Du, W.W.; Wu, N.; Yang, B.B. Posttranscriptional regulation of AKT by circular RNA angiomotin- like 1 mediates chemoresistance against paclitaxel in breast cancer cells. Aging 2019, 11, 11369–11381. [Google Scholar] [CrossRef]
- Zhong, Y.; Du, Y.; Yang, X.; Mo, Y.; Fan, C.; Xiong, F.; Ren, D.; Ye, X.; Li, C.; Wang, Y.; et al. Circular RNAs function as ceRNAs to regulate and control human cancer progression. Mol. Cancer 2018, 17, 79. [Google Scholar] [CrossRef]
- Li, H.; Xu, W.; Xia, Z.; Liu, W.; Pan, G.; Ding, J.; Li, J.; Wang, J.; Xie, X.; Jiang, D. Hsa_circ_0000199 facilitates chemo-tolerance of triple-negative breast cancer by interfering with miR-206/613-led PI3K/Akt/mTOR signaling. Aging 2021, 13, 4522–4551. [Google Scholar] [CrossRef]
- Zang, H.; Li, Y.; Zhang, X.; Huang, G. Circ-RNF111 contributes to paclitaxel resistance in breast cancer by elevating E2F3 expression via miR-140-5p. Thorac. Cancer 2020, 11, 1891–1903. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, Y.; Jiang, L.; Lu, L.; Dai, T.; Li, A.; Chen, Y.; Zhang, L. CircWAC induces chemotherapeutic resistance in triple-negative breast cancer by targeting miR-142, upregulating WWP1 and activating the PI3K/AKT pathway. Mol. Cancer 2021, 20, 43. [Google Scholar] [CrossRef]
- Hao, J.; Du, X.; Lv, F.; Shi, Q. Knockdown of circ_0006528 Suppresses Cell Proliferation, Migration, Invasion, and Adriamycin Chemoresistance via Regulating the miR-1236-3p/CHD4 Axis in Breast Cancer. J. Surg. Res. 2021, 260, 104–115. [Google Scholar] [CrossRef]
- Xie, H.; Zheng, R. Circ_0085495 knockdown reduces adriamycin resistance in breast cancer through miR-873-5p/integrin β1 axis. Anti-Cancer Drugs 2022, 33, e166–e177. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Fan, J.; Shi, W.; Zhou, Z. Circ_0001667 knockdown blocks cancer progression and attenuates adriamycin resistance by depleting NCOA3 via releasing miR-4458 in breast cancer. Drug Dev. Res. 2022, 83, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ren, Y.; Yao, R.; Zhou, L.; Fan, R. Circular RNA circ-MMP11 Contributes to Lapatinib Resistance of Breast Cancer Cells by Regulating the miR-153-3p/ANLN Axis. Front. Oncol. 2021, 11, 639961. [Google Scholar] [CrossRef]
- Yao, Y.; Li, X.; Cheng, L.; Wu, X.; Wu, B. Circular RNA FAT atypical cadherin 1 (circFAT1)/microRNA-525-5p/spindle and kinetochore-associated complex subunit 1 (SKA1) axis regulates oxaliplatin resistance in breast cancer by activating the notch and Wnt signaling pathway. Bioengineered 2021, 12, 4032–4043. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, Y.; Wang, F.; Li, L.; Qiu, X. CircFBXL5 promotes the 5-FU resistance of breast cancer via modulating miR-216b/HMGA2 axis. Cancer Cell Int. 2021, 21, 384. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Q.; He, S. Has_circ_0025202 suppresses cell tumorigenesis and tamoxifen resistance via miR-197-3p/HIPK3 axis in breast cancer. World J. Surg. Oncol. 2021, 19, 39. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, J.; Ao, N.; Yu, H.; Peng, Y.; Ou, L.; Zhang, S. Secondary cancer risk after radiation therapy for breast cancer with different radiotherapy techniques. Sci. Rep. 2020, 10, 1220. [Google Scholar] [CrossRef]
- Song, P.; Wu, J.; Chen, J.; Wang, F.; Chen, J.; Wang, G. Knockdown of circ-ADAM9 inhibits malignant phenotype and enhances radiosensitivity in breast cancer cells via acting as a sponge for miR-383-5p. Strahlenther. Onkol. 2023, 199, 78–89. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, J.; Wang, H.; Li, J. Circ-ABCC1 enhances radioresistance of breast cancer cells via miR-627-5p/ABCC1 axis. Cell. Mol. Biol. 2022, 68, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Urbano, M.A.; Griñán-Lisón, C.; Marchal, J.A.; Núñez, M.I. CSC radioresistance: A therapeutic challenge to improve radiotherapy effectiveness in cancer. Cell 2020, 9, 1651. [Google Scholar] [CrossRef]
- Li, F.; Zhou, K.; Gao, L.; Zhang, B.; Li, W.; Yan, W.; Song, X.; Yu, H.; Wang, S.; Yu, N.; et al. Radiation induces the generation of cancer stem cells: A novel mechanism for cancer radioresistance. Oncol. Lett. 2016, 12, 3059–3065. [Google Scholar] [CrossRef]
- Inalegwu, A.; Cuypers, B.; Claesen, J.; Janssen, A.; Coolkens, A.; Baatout, S.; Laukens, K.; De Vos, W.H.; Quintens, R. Fractionated irradiation of MCF7 breast cancer cells rewires a gene regulatory circuit towards a treatment-resistant stemness phenotype. Mol. Oncol. 2022, 16, 3410–3435. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Shen, D.; Liu, Z.; Zhang, J.; Zhang, J.; Geng, C. Circ_0008500 Knockdown Improves Radiosensitivity and Inhibits Tumorigenesis in Breast Cancer Through the miR-758-3p/PFN2 Axis. J. Mammary Gland Biol. 2022, 27, 37–52. [Google Scholar] [CrossRef] [PubMed]
- He, Z.Y.; Zhuo, R.G.; Yang, S.P.; Zhou, P.; Xu, J.Y.; Zhou, J.; Wu, S.G. CircNCOR1 regulates breast cancer radiotherapy efficacy by regulating CDK2 via hsa-miR-638 binding. Cell. Signal. 2023, 109, 110787. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| CircRNA | Expression in TNBC | Regulated Axis | Functions and Prognosis | Reference |
|---|---|---|---|---|
| circZEB1 | ↑ | circZEB1/miR-448/eEF2K | Cell proliferation. | [41] |
| circSKA3 | ↑ | - | Tumor progression. Poor prognosis. Exosomal transmission. | [42] |
| circ-PDCD11 | ↑ | circ-PDCD11/miR-512-3p/CDCA3 | Accelerated glucose uptake, lactate production, ATP generation, and extracellular acidification. Poor prognosis of TNBC. | [44] |
| circRNA-UCK2 | ↑ | circRNA-UCK2/miR-767-5p/TET1 | Tumor progression. Poor prognosis. | [43] |
| circANKS1B | ↑ | circANKS1B/miR-148a-3p/miR-152-3p/USF1 | Metastasis. | [45] |
| circGFRA1 | circGFRA1/miR-361-5p/TLR4 | Resistance to paclitaxel. | [46] | |
| circEPSTI1 | ↑ | circEPSTI1/miR-4753/BCL11A | Cell proliferation. | [47] |
| circHIF1A | ↑ | circHIF1A//NFIB/FUScircHIF1A/miR-580-5p/CD44 | Packaged into exosomes. Metastasis and poor prognosis of TNBC. | [48,49] |
| circBACH2 | ↑ | circBACH2/miR-186-5p/miR-548c-3p/CXCR4 | Cell proliferation, invasion, and metastasis. | [50] |
| circSEPT9 | ↑ | circSEPT9/miR-637/LIF | Facilitation of the carcinogenesis and development of TNBC. | [51] |
| circAGFG1 | ↑ | circAGFG1/miR-195-5p/CCNE1 | Cancer progression. | [52] |
| circ-HER2 | ↑ | - | Cell proliferation, invasion, and metastasis. | [53] |
| circRPPH1 | ↑ | circRPPH1/miR-556-5p/YAP1 | Cell proliferation, invasion, and metastasis. | [54] |
| hsa_circ_0131242 | ↑ | has_circ_0131242/miR-2682 | Cancer progression. | [55] |
| hsa_circ_0000520 | ↑ | hsa_circ_0000520/miR-1296/ZFX | Regulation of transcription. | [56] |
| circGNB1 | ↓ | circGNB1/miR-141-5p/IGF1R | Regulation of tumor progression. | [57] |
| circUSP42 | ↑ | circUBE2D2/miR-512-3p/CDCA3 | Cell proliferation, metastasis, and chemoresistance. | [58] |
| circCDYL | ↓ | circCDYL/miR-190a-3p/TP53INP1 | Tumor suppression. | [59] |
| circNR3C2 | ↓ | circNR3C2/miR-513a-3p/HRD1 | Inhibition of cell proliferation, migration, invasion, and EMT process. | [60] |
| circAHNAK1 | ↑ | circAHNAK1/miR-421/RASA1 | Cell proliferation. | [61] |
| hsa_circ_0091074 | ↑ | hsa_circ_0091074/miR-1297/TAZ | Regulation of cell cycle. | [62] |
| circUBE2D2 | ↑ | circUBE2D2/miR-512-3p/CDCA3 | Cell proliferation, metastasis, and chemoresistance. | [63] |
| hsa_circ_0092276 | hsa_circ_0092276/miR-384/ATG7 | Autophagy and DOX resistance. | [64] | |
| CircKDM4C | ↓ | CircKDM4C/miR-548p | Inhibited DOX resistance. | [65] |
| circSMARCA5 | ↓ | circSMARCA5/miR-548p/SMARCA5 | Prevention of chemoresistance. | [66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zepeda-Enríquez, P.; Silva-Cázares, M.B.; López-Camarillo, C. Novel Insights into Circular RNAs in Metastasis in Breast Cancer: An Update. Non-Coding RNA 2023, 9, 55. https://doi.org/10.3390/ncrna9050055
Zepeda-Enríquez P, Silva-Cázares MB, López-Camarillo C. Novel Insights into Circular RNAs in Metastasis in Breast Cancer: An Update. Non-Coding RNA. 2023; 9(5):55. https://doi.org/10.3390/ncrna9050055
Chicago/Turabian StyleZepeda-Enríquez, Paola, Macrina B. Silva-Cázares, and César López-Camarillo. 2023. "Novel Insights into Circular RNAs in Metastasis in Breast Cancer: An Update" Non-Coding RNA 9, no. 5: 55. https://doi.org/10.3390/ncrna9050055
APA StyleZepeda-Enríquez, P., Silva-Cázares, M. B., & López-Camarillo, C. (2023). Novel Insights into Circular RNAs in Metastasis in Breast Cancer: An Update. Non-Coding RNA, 9(5), 55. https://doi.org/10.3390/ncrna9050055