

Ameliorative Effects of Camel Milk and Fermented Camel Milk on Acute Alcoholic Liver Injury

Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Animals and Experimental Design
3. Methodologies
3.1. Liver Tissue and Serum Biochemical Analyses
3.2. Detection of Hepatic Cytokines via Real-Time Quantitative PCR
3.3. Histopathologic Analysis
3.4. Sequencing and Analysis of Fecal Bacterial 16S rRNA Genes
3.4.1. DNA Extraction and PCR Amplification
3.4.2. Bioinformatics Analysis
3.5. Metabolomic Analysis
3.6. Analyzing Statistics
4. Results
4.1. Behavioral Aspects of the Gavage Dose in Mice
4.2. CM and FCM Ameliorate Alcohol-Induced Oxidative Damage
4.3. Effects of CM and FCM on Liver Histopathology in Mice with Alcohol-Induced Liver Injury
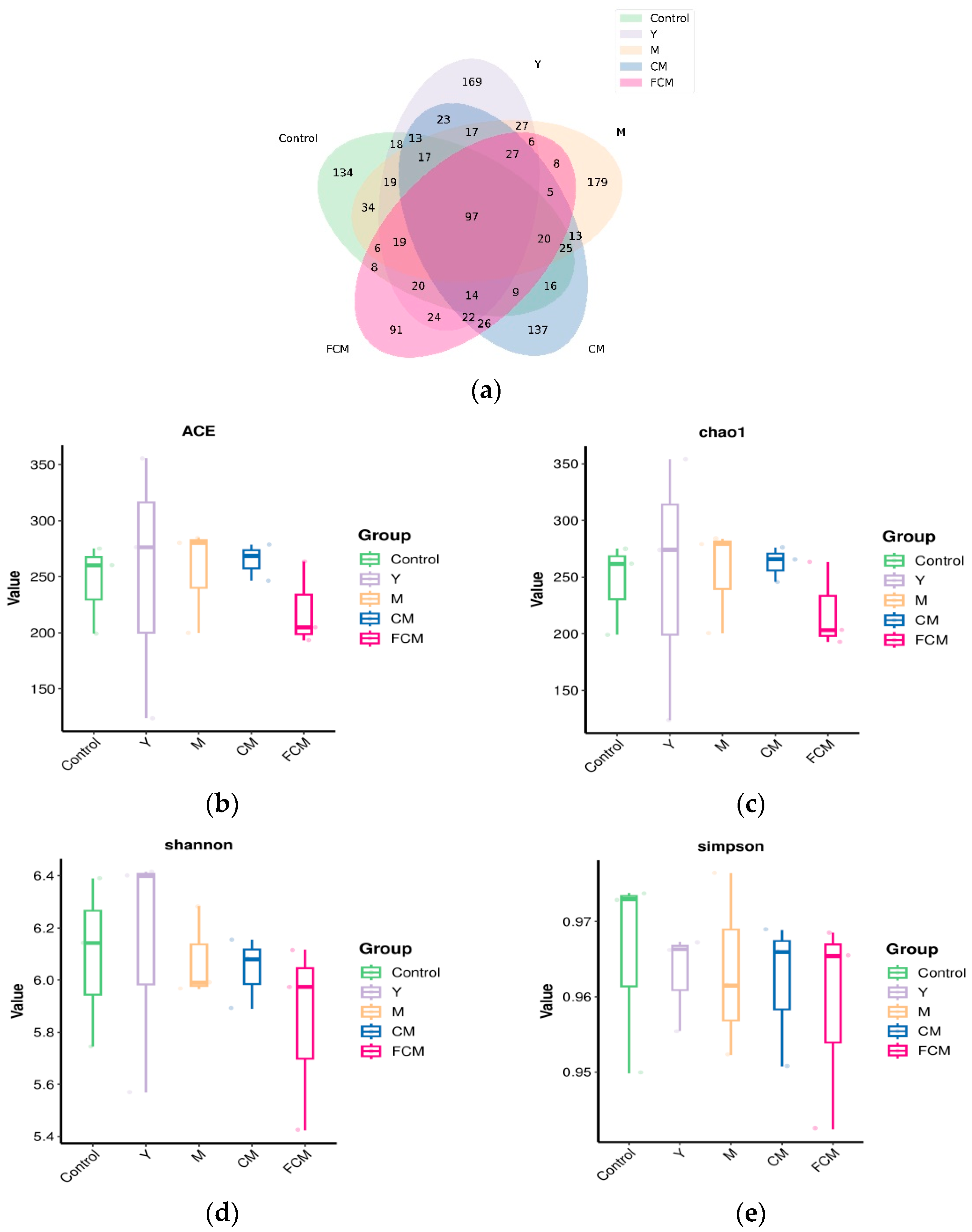
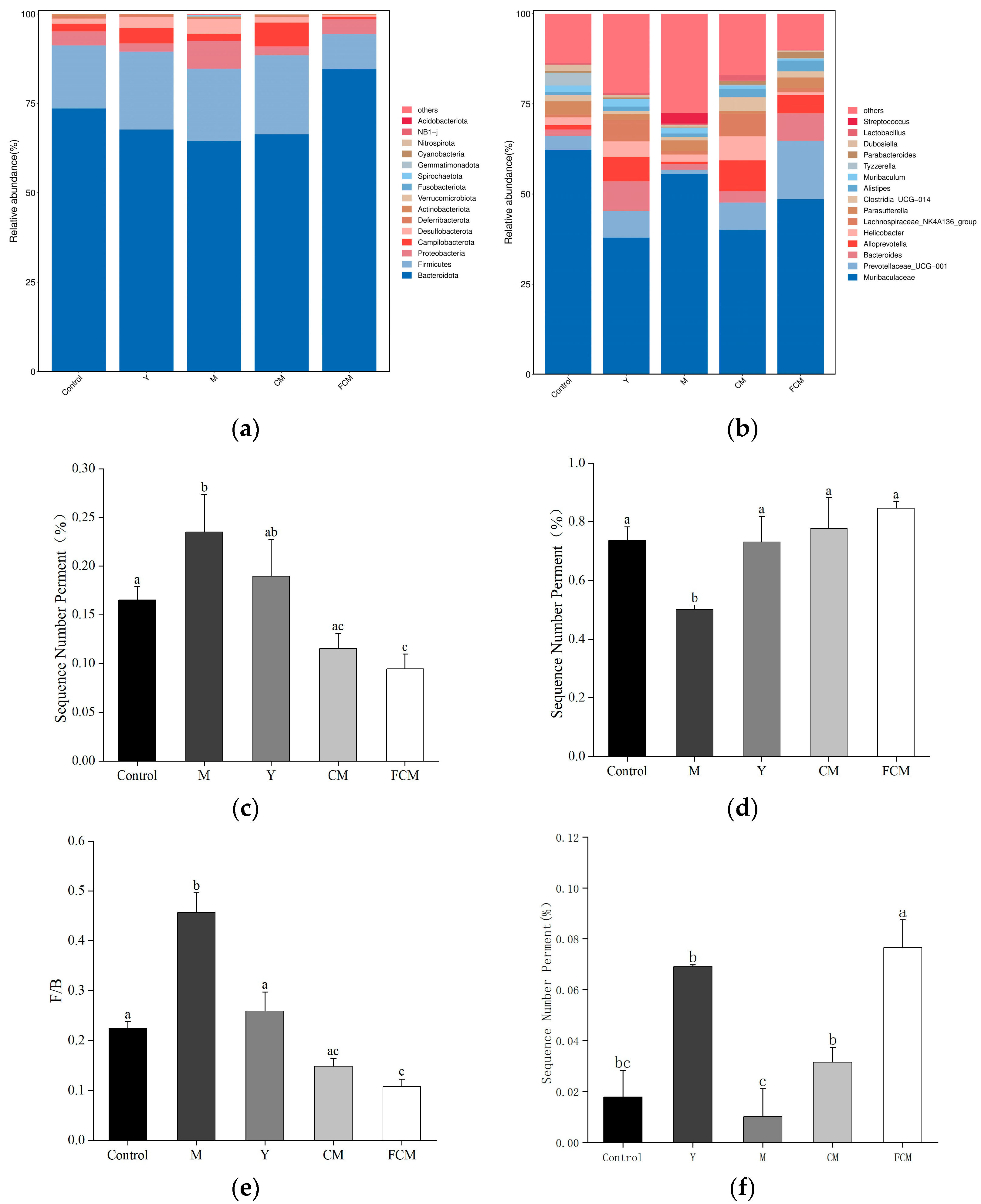
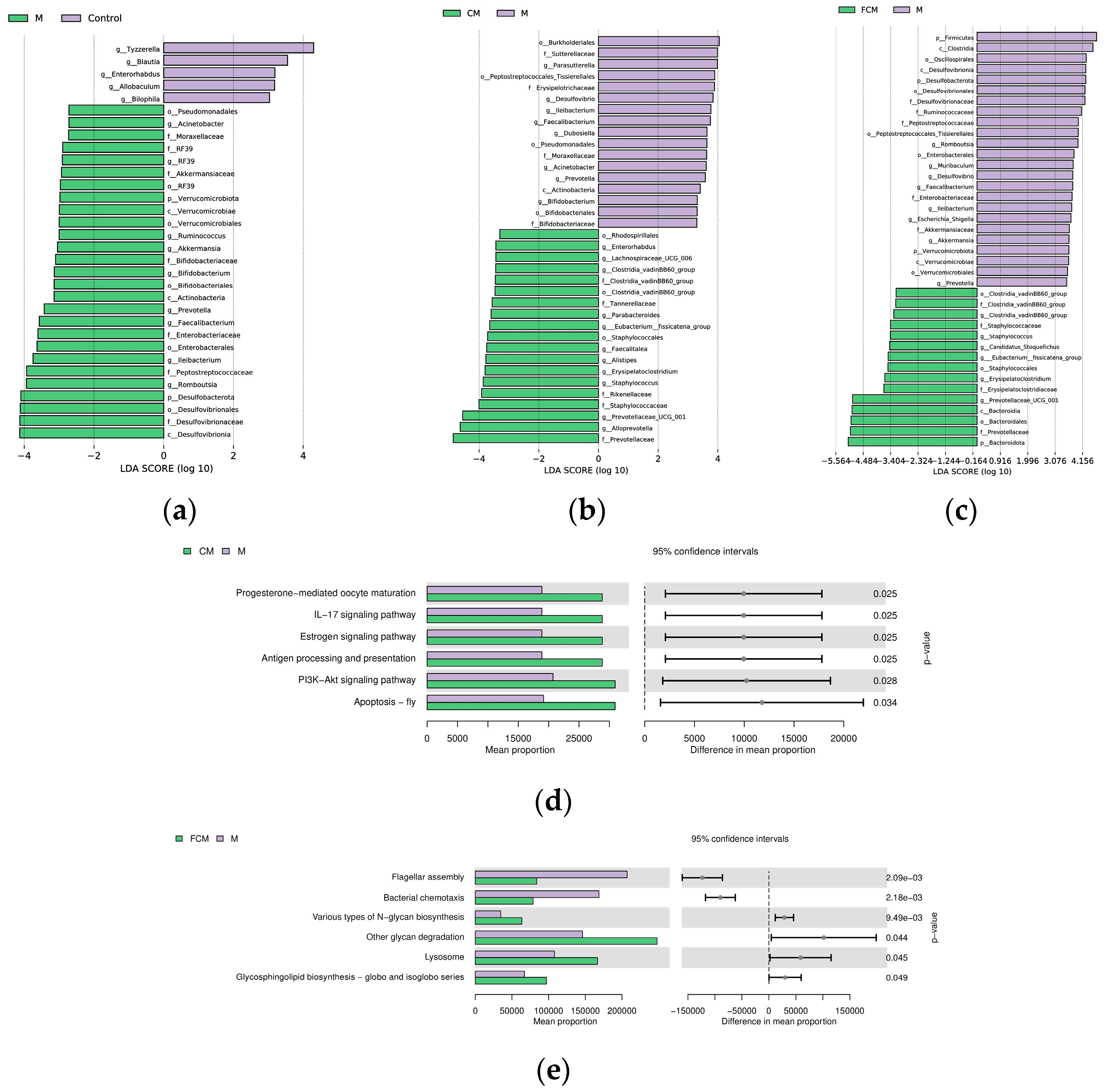
4.4. Results of CM and FCM on the Microbiota of the Intestines of Mice Subjected to Alcohol-Induced Liver Dysfunction
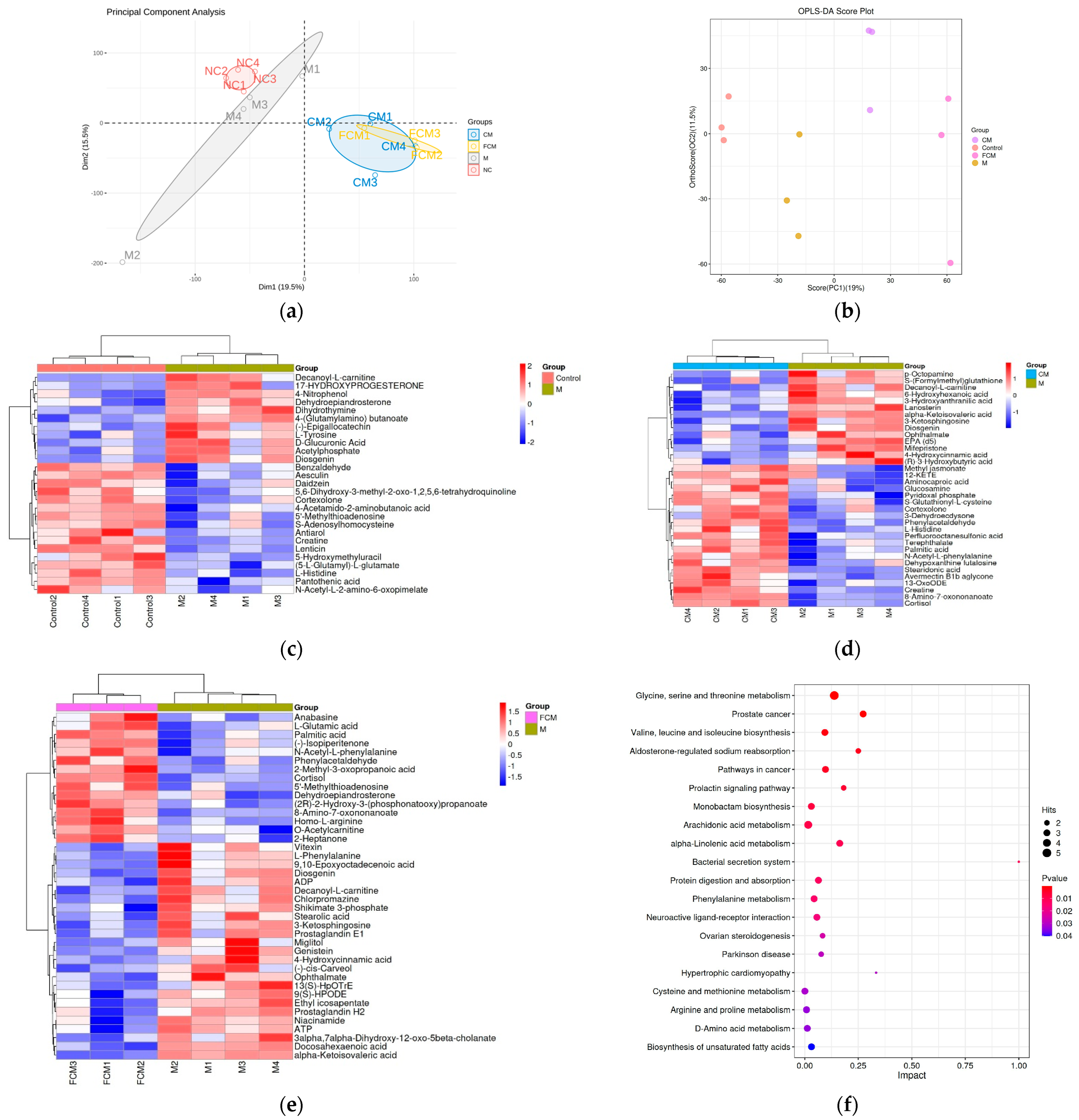
4.5. Effects of CM and FCM on Liver Metabolites in Mice
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Subramaniyan, V.; Chakravarthi, S.; Jegasothy, R.; Seng, W.Y.; Fuloria, N.K.; Fuloria, S.; Hazarika, I.; Das, A. Alcohol-Associated Liver Disease: A Review on Its Pathophysiology, Diagnosis and Drug Therapy. Toxicol. Rep. 2021, 8, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Ayares, G.; Idalsoaga, F.; Arnold, J.; Fuentes-López, E.; Arab, J.P.; Díaz, L.A. Public Health Measures and Prevention of Alcohol-Associated Liver Disease. J. Clin. Exp. Hepatol. 2022, 12, 1480–1491. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Wang, Y.; You, Y.; Ai, Z.; Dai, W.; Piao, C.; Liu, J.; Wang, Y. Fermented Ginseng Improved Alcohol Liver Injury in Association with Changes in the Gut Microbiota of Mice. Food Funct. 2019, 10, 5566–5573. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.-L.; Shen, Y.-H.; Song, Y.-J.; Xu, Y.; Xu, H.-X.; Prunella Vulgaris, L. Attenuates Gut Dysbiosis and Endotoxin Leakage against Alcoholic Liver Disease. J. Ethnopharmacol. 2024, 319, 117237. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Li, W.; Chen, G.; Yang, Z.; Lv, X.; Zheng, L.; Sun, J.; Ai, L.; Sun, B.; Ni, L. Ameliorative Effects of Monascin from Red Mold Rice on Alcoholic Liver Injury and Intestinal Microbiota Dysbiosis in Mice. Food Biosci. 2022, 50, 102079. [Google Scholar] [CrossRef]
- De Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut Microbiome and Health: Mechanistic Insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Meena, A.S.; Shukla, P.K.; Bell, B.; Giorgianni, F.; Caires, R.; Fernández-Peña, C.; Beranova, S.; Aihara, E.; Montrose, M.H.; Chaib, M.; et al. TRPV6 Channel Mediates Alcohol-Induced Gut Barrier Dysfunction and Systemic Response. Cell Rep. 2022, 39, 110937. [Google Scholar] [CrossRef]
- Liu, H.; Kang, X.; Yang, X.; Yang, H.; Kuang, X.; Ren, P.; Yan, H.; Shen, X.; Kang, Y.; Li, L.; et al. Compound Probiotic Ameliorates Acute Alcoholic Liver Disease in Mice by Modulating Gut Microbiota and Maintaining Intestinal Barrier. Probiotics Antimicrob. Proteins 2023, 15, 185–201. [Google Scholar] [CrossRef]
- Sang, T.; Guo, C.; Guo, D.; Wu, J.; Wang, Y.; Wang, Y.; Chen, J.; Chen, C.; Wu, K.; Na, K.; et al. Suppression of Obesity and Inflammation by Polysaccharide from Sporoderm-Broken Spore of Ganoderma Lucidum via Gut Microbiota Regulation. Carbohydr. Polym. 2021, 256, 117594. [Google Scholar] [CrossRef]
- Sun, X.; Shi, J.; Kong, L.; Shen, Q.; Zeng, X.; Wu, Z.; Guo, Y.; Pan, D. Recent Insights into the Hepatoprotective Effects of Lactic Acid Bacteria in Alcoholic Liver Disease. Trends Food Sci. Technol. 2022, 125, 91–99. [Google Scholar] [CrossRef]
- Cao, H.; Zhou, T.; Tang, H.; Feng, P.; Ali, G.; Liu, P.; Li, X. Genetically Encoded Probiotic EcN 1917 Alleviates Alcohol-Induced Acute Liver Injury and Restore Gut Microbiota Homeostasis. J. Funct. Foods 2021, 85, 104661. [Google Scholar] [CrossRef]
- Tian, X.; Li, R.; Jiang, Y.; Zhao, F.; Yu, Z.; Wang, Y.; Dong, Z.; Liu, P.; Li, X. Bifidobacterium breve ATCC15700 Pretreatment Prevents Alcoholic Liver Disease through Modulating Gut Microbiota in Mice Exposed to Chronic Alcohol Intake. J. Funct. Foods 2020, 72, 104045. [Google Scholar] [CrossRef]
- Xiong, S.-Y.; Wu, G.-S.; Li, C.; Ma, W.; Luo, H.-R. Clinical Efficacy of Probiotics in the Treatment of Alcoholic Liver Disease: A Systematic Review and Meta-Analysis. Front. Cell. Infect. Microbiol. 2024, 14, 1358063. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Woo, D.; Nam, Y.; Baek, J.; Lee, J.-Y.; Kim, W. Probiotic Cheese Improves Alcohol Metabolism and Alleviates Alcohol-Induced Liver Injury via the SIRT1/AMPK Signaling Pathway. J. Funct. Foods 2023, 108, 105736. [Google Scholar] [CrossRef]
- Nam, Y.; Kim, J.-H.; Konkit, M.; Kim, W. Hepatoprotective Effects of Lactococcus Chungangensis CAU 1447 in Alcoholic Liver Disease. J. Dairy Sci. 2019, 102, 10737–10747. [Google Scholar] [CrossRef]
- Ayyash, M.; Abdalla, A.; Alhammadi, A.; Senaka Ranadheera, C.; Affan Baig, M.; Al-Ramadi, B.; Chen, G.; Kamal-Eldin, A.; Huppertz, T. Probiotic Survival, Biological Functionality and Untargeted Metabolomics of the Bioaccessible Compounds in Fermented Camel and Bovine Milk after in Vitro Digestion. Food Chem. 2021, 363, 130243. [Google Scholar] [CrossRef]
- Bao, W.; He, Y.; Yu, J.; Yang, X.; Liu, M.; Ji, R. Diversity Analysis and Gene Function Prediction of Bacteria and Fungi of Bactrian Camel Milk and Naturally Fermented Camel Milk from Alxa in Inner Mongolia. LWT 2022, 169, 114001. [Google Scholar] [CrossRef]
- He, J.; Guo, K.; Chen, Q.; Wang, Y. Jirimutu Camel Milk Modulates the Gut Microbiota and Has Anti-Inflammatory Effects in a Mouse Model of Colitis. J. Dairy Sci. 2022, 105, 3782–3793. [Google Scholar] [CrossRef]
- Al-Anazi, M.S.; El-Zahar, K.M.; Rabie, N.A.-H. Nutritional and Therapeutic Properties of Fermented Camel Milk Fortified with Red Chenopodium Quinoa Flour on Hypercholesterolemia Rats. Molecules 2022, 27, 7695. [Google Scholar] [CrossRef]
- Ayoub, M.A.; Palakkott, A.R.; Ashraf, A.; Iratni, R. The Molecular Basis of the Anti-Diabetic Properties of Camel Milk. Diabetes Res. Clin. Pract. 2018, 146, 305–312. [Google Scholar] [CrossRef]
- Han, B.; Zhang, L.; Hou, Y.; Zhong, J.; Hettinga, K.; Zhou, P. Phosphoproteomics Reveals That Camel and Goat Milk Improve Glucose Homeostasis in HDF/STZ-Induced Diabetic Rats through Activation of Hepatic AMPK and GSK3-GYS Axis. Food Res. Int. 2022, 157, 111254. [Google Scholar] [CrossRef] [PubMed]
- Korish, A.A.; Abdel Gader, A.G.M.; Alhaider, A.A. Comparison of the Hypoglycemic and Antithrombotic (Anticoagulant) Actions of Whole Bovine and Camel Milk in Streptozotocin-Induced Diabetes Mellitus in Rats. J. Dairy Sci. 2020, 103, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.; Ge, W.; Song, Y.; He, R.; Wang, Z.; Zhao, L. Camel Milk Peptides Alleviate Hyperglycemia by Regulating Gut Microbiota and Metabolites in Type 2 Diabetic Mice. Food Res. Int. 2023, 173, 113278. [Google Scholar] [CrossRef] [PubMed]
- Darwish, H.A.; Abd Raboh, N.R.; Mahdy, A. Camel’s Milk Alleviates Alcohol-Induced Liver Injury in Rats. Food Chem. Toxicol. 2012, 50, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.; Qiao, X.; Yi, L.; Siren, D.; He, J.; Hai, L.; Guo, F.; Xiao, Y.; Ji, R. Camel Milk Modulates Ethanol-Induced Changes in the Gut Microbiome and Transcriptome in a Mouse Model of Acute Alcoholic Liver Disease. J. Dairy Sci. 2020, 103, 3937–3949. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.; Qi, B.; Hao, S.; Ji, R. Camel milk ameliorates inflammatory mechanisms in an alcohol-induced liver injury mouse model. Sci. Rep. 2021, 11, 22811. [Google Scholar] [CrossRef]
- Qi, B.; Ji Ri, M.; Ming, L. The intervention of camel milk on acute alcoholic liver injury in mice. J. Chin. Inst. Food Sci. Technol. 2023, 23, 147–158. [Google Scholar] [CrossRef]
- Liu, M.; Liu, M.; Yang, S.; Shen, C.; Wang, X.; Liu, W.; Guo, Y. Fermented Milk of Cheese-Derived Lactobacillus Delbrueckii Subsp. Bulgaricus Displays Potentials in Alleviating Alcohol-Induced Hepatic Injury and Gut Dysbiosis in Mice. Food Res. Int. 2022, 157, 111283. [Google Scholar] [CrossRef]
- Han, X.; Liu, J.; Bai, Y.; Hang, A.; Lu, T.; Mao, C. An Iridoid Glycoside from Cornus Officinalis Balances Intestinal Microbiome Disorder and Alleviates Alcohol-Induced Liver Injury. J. Funct. Foods 2021, 82, 104488. [Google Scholar] [CrossRef]
- Ma, T.; Li, Y.; Zhu, Y.; Jiang, S.; Cheng, C.; Peng, Z.; Xu, L. Differential Metabolic Pathways and Metabolites in a C57BL/6J Mouse Model of Alcoholic Liver Disease. Med. Sci. Monit. 2020, 26, e924602. [Google Scholar] [CrossRef]
- Fairfield, B. Gut Dysbiosis as a Driver in Alcohol-Induced Liver Injury. Jhep Rep. 2021, 3, 100220. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.; Cai, Z.; Yang, Q.; Liu, J.; Yu, Y.; Pan, F.; Zhang, T. Soybean Meal Peptide Gly-Thr-Tyr-Trp Could Protect Mice from Acute Alcoholic Liver Damage: A Study of Protein-Protein Interaction and Proteomic Analysis. Food Chem. 2024, 451, 139337. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, H.; Liang, J.; Sun, C.; Wang, D.; Chen, K.; Zhao, J.; Ji, S.; Ma, C.; Ye, X.; et al. Hepatoprotective Potential of Four Fruit Extracts Rich in Different Structural Flavonoids against Alcohol-Induced Liver Injury via Gut Microbiota-Liver Axis. Food Chem. 2024, 460, 140460. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, Z.; Wang, X.; Li, M.; Zhou, B.; Zhang, X. Solanum nigrum L. Berries Extract Ameliorated the Alcoholic Liver Injury by Regulating Gut Microbiota, Lipid Metabolism, Inflammation, and Oxidative Stress. Food Res. Int. 2024, 188, 114489. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Al-Nuaimi, A.K.; Al-Mahadin, S.; Liu, S.-Q. In Vitro Investigation of Anticancer and ACE-Inhibiting Activity, α-Amylase and α-Glucosidase Inhibition, and Antioxidant Activity of Camel Milk Fermented with Camel Milk Probiotic: A Comparative Study with Fermented Bovine Milk. Food Chem. 2018, 239, 588–597. [Google Scholar] [CrossRef]
- Ayyash, M.; Al-Dhaheri, A.S.; Al Mahadin, S.; Kizhakkayil, J.; Abushelaibi, A. In Vitro Investigation of Anticancer, Antihypertensive, Antidiabetic, and Antioxidant Activities of Camel Milk Fermented with Camel Milk Probiotic: A Comparative Study with Fermented Bovine Milk. J. Dairy Sci. 2018, 101, 900–911. [Google Scholar] [CrossRef]
- Khakhariya, R.; Sakure, A.A.; Maurya, R.; Bishnoi, M.; Kondepudi, K.K.; Padhi, S.; Rai, A.K.; Liu, Z.; Patil, G.B.; Mankad, M.; et al. A Comparative Study of Fermented Buffalo and Camel Milk with Anti-Inflammatory, ACE-Inhibitory and Anti-Diabetic Properties and Release of Bio Active Peptides with Molecular Interactions: In Vitro, in Silico and Molecular Study. Food Biosci. 2023, 52, 102373. [Google Scholar] [CrossRef]
- Aljutaily, T.; Rehan, M.; Moustafa, M.M.A.; Barakat, H. Effect of Intermittent Fasting, Probiotic-Fermented Camel Milk, and Probiotic-Fermented Camel Milk Incorporating Sukkari Date on Diet-Induced Obesity in Rats. Fermentation 2022, 8, 619. [Google Scholar] [CrossRef]
- Berhe, T.; Ipsen, R.; Seifu, E.; Kurtu, M.Y.; Eshetu, M.; Hansen, E.B. Comparison of the Acidification Activities of Commercial Starter Cultures in Camel and Bovine Milk. LWT 2018, 89, 123–127. [Google Scholar] [CrossRef]
- Atwaa, E.S.H.; Shahein, M.R.; Raya-Alvarez, E.; Abd El-Sattar, E.S.; Hassan, M.A.A.; Hashim, M.A.; Dahran, N.; El-Khadragy, M.F.; Agil, A.; Elmahallawy, E.K. Assessment of the Physicochemical and Sensory Characteristics of Fermented Camel Milk Fortified with Cordia Myxa and Its Biological Effects against Oxidative Stress and Hyperlipidemia in Rats. Front. Nutr. 2023, 10, 1130224. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of Potential Probiotic Lactic Acid Bacteria Isolated from Camel Milk. LWT—Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Lai, W.; Zhou, S.; Bai, Y.; Che, Q.; Cao, H.; Guo, J.; Su, Z. Glucosamine Attenuates Alcohol-Induced Acute Liver Injury via Inhibiting Oxidative Stress and Inflammation. Curr. Res. Food Sci. 2024, 8, 100699. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Liu, W.; Chen, L.; Ni, L.; Zou, X.; Ye, M.; Zhang, Z.; Zou, S. Flavonoid and Chromone-Rich Extract from Euscaphis Konishii Hayata Leaf Attenuated Alcoholic Liver Injury in Mice. J. Ethnopharmacol. 2022, 295, 115455. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Li, S.; Song, C.; Sun, X.; Liu, X. Black Soybean-Derived Peptides Exerted Protective Effect against Alcohol-Induced Liver Injury in Mice. J. Funct. Foods 2021, 87, 104828. [Google Scholar] [CrossRef]
- Wu, K.; Li, J.; Zhou, X.; Zhou, F.; Tang, S.; Yi, L.; Wu, Y.; Tian, S. NADH and NRH as Potential Dietary Supplements or Pharmacological Agents for Early Liver Injury Caused by Acute Alcohol Exposure. J. Funct. Foods 2021, 87, 104852. [Google Scholar] [CrossRef]
- Ding, Q.; Cao, F.; Lai, S.; Zhuge, H.; Chang, K.; Valencak, T.G.; Liu, J.; Li, S.; Ren, D. Lactobacillus Plantarum ZY08 Relieves Chronic Alcohol-Induced Hepatic Steatosis and Liver Injury in Mice via Restoring Intestinal Flora Homeostasis. Food Res. Int. 2022, 157, 111259. [Google Scholar] [CrossRef]
- Shu, X.; Wang, J.; Zhao, L.; Wang, J.; Wang, P.; Zhang, F.; Wang, R. Bifidobacterium Lactis TY-S01 Protects against Alcoholic Liver Injury in Mice by Regulating Intestinal Barrier Function and Gut Microbiota. Heliyon 2023, 9, e17878. [Google Scholar] [CrossRef]
- Guo, W.-L.; Cao, Y.-J.; You, S.-Z.; Wu, Q.; Zhang, F.; Han, J.-Z.; Lv, X.-C.; Rao, P.-F.; Ai, L.-Z.; Ni, L. Ganoderic Acids-Rich Ethanol Extract from Ganoderma Lucidum Protects against Alcoholic Liver Injury and Modulates Intestinal Microbiota in Mice with Excessive Alcohol Intake. Curr. Res. Food Sci. 2022, 5, 515–530. [Google Scholar] [CrossRef]
- Wang, W.; Yang, S.; Song, S.; Zhang, J.; Jia, F. Flammulina Velutipes Mycorrhizae Dietary Fiber Improves Lipid Metabolism Disorders in Obese Mice through Activating AMPK Signaling Pathway Mediated by Gut Microbiota. Food Biosci. 2021, 43, 101246. [Google Scholar] [CrossRef]
- Song, X.; Cui, W.; Meng, F.; Xia, Q.; Li, X.; Hou, M.; Jia, L.; Zhang, J. Glucopyranose from Pleurotus geesteranus Prevent Alcoholic Liver Diseases by Regulating Nrf2/HO-1-TLR4/NF-κB Signalling Pathways and Gut Microbiota. Food Funct. 2022, 13, 2441–2455. [Google Scholar] [CrossRef]
- Zeng, S.; Cao, J.; Wei, C.; Chen, Y.; Liu, Q.; Li, C.; Zhang, Y.; Zhu, K.; Wu, G.; Tan, L. Polysaccharides from Artocarpus Heterophyllus Lam. (Jackfruit) Pulp Alleviate Obesity by Modulating Gut Microbiota in High Fat Diet-Induced Rats. Food Hydrocoll. 2023, 139, 108521. [Google Scholar] [CrossRef]
- Tao, K.; Peng, H.; Bi, X.; Wang, X.; Tian, Y.; Zhang, H.; Sun, Y. Effect of Tetrastigma Hemsleyanum Leaves and Extract Supplementation on Liver Metabolomics and the Gut Microbiota in Alcohol-Induced Liver Injury. Food Biosci. 2024, 58, 103617. [Google Scholar] [CrossRef]
- Baweja, S.; Mittal, A.; Thangariyal, S.; Subudhi, P.D.; Gautam, S.; Kaul, R. Unveiling the Effect of Estrogen Receptors in Alcoholic Liver Disease: A Novel Outlook. Liver Res. 2023, 7, 333–341. [Google Scholar] [CrossRef]
- He, Q.; Yang, C.; Kang, X.; Chen, Y.; Zhang, T.; Zhang, H.; Kwok, L.-Y. Intake of Bifidobacterium Lactis Probio-M8 Fermented Milk Protects against Alcoholic Liver Disease. J. Dairy Sci. 2022, 105, 2908–2921. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| IL-6 | TTCTCTGGGAAATCGTGGAAA | TGCAAGTGCATCATCGTTGT |
| IL-1β | GCAACTGTTCCTGAACTCAACT | ATCTTTTGGGGTCCGTCAACT |
| PPAR-α | CCTGGAAAGTCCCTTATCT | GCCCTTACAGCCTTCACAT |
| Group | M | Y | CM | FCM |
|---|---|---|---|---|
| Gavage time | 14:50–14:56 | 15:48–15:52 | 12:07–12:16 | 11:58–12:02 |
| Sober time | 15:27–16:10 | 16:10–18:00 | 13:45–12:29 | 12:13–12:37 |
| Mean time (min) | 73.50 ± 21.96 b | 38.00 ± 8.49 a | 16.25 ± 2.65 a | 27.00 ± 4.51 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Sun, W.; Luo, Y. Ameliorative Effects of Camel Milk and Fermented Camel Milk on Acute Alcoholic Liver Injury. Fermentation 2024, 10, 493. https://doi.org/10.3390/fermentation10100493
Zhu C, Sun W, Luo Y. Ameliorative Effects of Camel Milk and Fermented Camel Milk on Acute Alcoholic Liver Injury. Fermentation. 2024; 10(10):493. https://doi.org/10.3390/fermentation10100493
Chicago/Turabian StyleZhu, Chunxia, Wancheng Sun, and Yihao Luo. 2024. "Ameliorative Effects of Camel Milk and Fermented Camel Milk on Acute Alcoholic Liver Injury" Fermentation 10, no. 10: 493. https://doi.org/10.3390/fermentation10100493
APA StyleZhu, C., Sun, W., & Luo, Y. (2024). Ameliorative Effects of Camel Milk and Fermented Camel Milk on Acute Alcoholic Liver Injury. Fermentation, 10(10), 493. https://doi.org/10.3390/fermentation10100493