

Mechanistic Insights into Nitrite Degradation by Metabolites of L. plantarum A50: An LC-MS-Based Untargeted Metabolomics Analysis

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preservation and Activation of L. plantarum A50
2.2. Research on the Growth Characteristics and Tolerance of L. plantarum A50
2.3. Research on the Ability of L. plantarum A50 Metabolites to Degrade Nitrite
2.3.1. The Capacity of L. plantarum A50 to Degrade Nitrite
2.3.2. The Influence of Varying Acidity on the Degradation of Nitrite
2.3.3. The Capacity of Metabolites Derived from L. plantarum A50 and MRS Broth in the Degradation of Nitrite
2.3.4. The Influence of Varying Levels of Acidity on the Capacity of L. plantarum A50 Metabolites to Degrade Nitrite
2.4. Conducting an Untargeted Metabolomics Analysis on the Fermentation Broth of L. plantarum A50
2.4.1. Sample Preparation
2.4.2. LC-MS/MS Data Analysis
2.4.3. Data Analysis
2.5. Statistical Analysis
3. Results
3.1. The Growth Trajectories and Physiological as well as Biochemical Markers of Lactobacillus plantarum A50
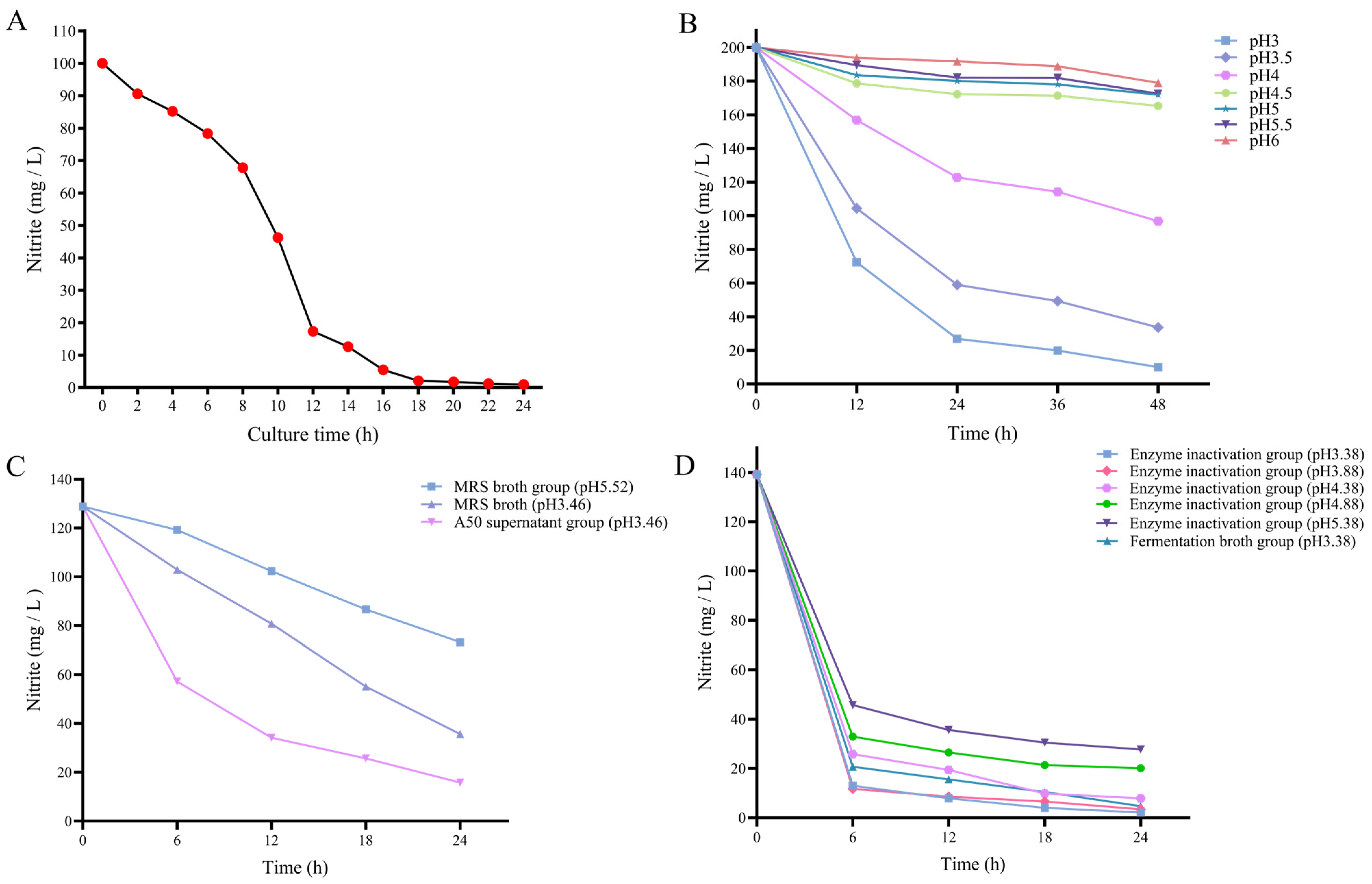
3.2. Research on the Nitrite Degradation Ability of L. plantarum A50
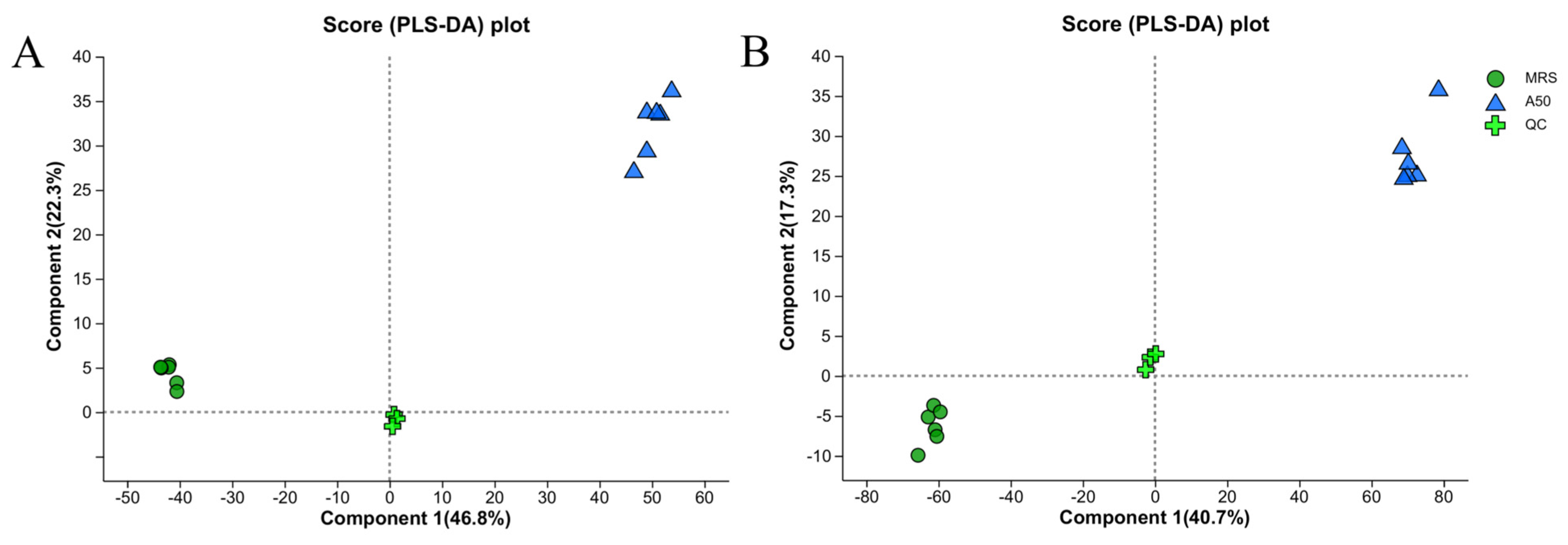
3.3. Metabolite Analysis of L. plantarum A50
4. Discussion
4.1. Physiological Characteristics of L. plantarum A50
4.2. Degradation of Nitrite by L. plantarum A50
4.3. Metabolite Analysis of L. plantarum A50
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS); Mortensen, A.; Aguilar, F.; Crebelli, R.; Di Domenico, A.; Dusemund, B.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; et al. Re-evaluation of sodium nitrate (E 251) and potassium nitrate (E 252) as food additives. EFSA J. 2017, 15, e04787. [Google Scholar] [CrossRef]
- MAC Donald, B.; Gray, J.I.; Gibbins, L.N. Role of nitrite in cured meat flavor: Antioxidant role of nitrite. J. Food Sci. 1980, 45, 893–897. [Google Scholar] [CrossRef]
- Macdougall, D.B.; Mottram, D.S.; Rhodes, D.N. Contribution of nitrite and nitrate to the colour and flavour of cured meats. J. Sci. Food Agric. 1975, 26, 1743–1754. [Google Scholar] [CrossRef]
- Shakil, M.H.; Trisha, A.T.; Rahman, M.; Talukdar, S.; Kobun, R.; Huda, N.; Zzaman, W. Nitrites in Cured Meats, Health Risk Issues, Alternatives to Nitrites: A Review. Foods 2022, 11, 3355. [Google Scholar] [CrossRef]
- Calvert, J.W.; Lefer, D.J. Clinical translation of nitrite therapy for cardiovascular diseases. Nitric Oxide 2010, 22, 91–97. [Google Scholar] [CrossRef]
- Tang, Y.; Jiang, H.; Bryan, N.S. Nitrite and nitrate: Cardiovascular risk–benefit and metabolic effect. Curr. Opin. Lipidol. 2011, 22, 11–15. [Google Scholar] [CrossRef]
- König, W.; König, E.; Weiss, K.; Tuomivirta, T.T.; Fritze, H.; Elo, K.; Vanhatalo, A.; Jaakkola, S. Impact of hexamine addition to a nitrite-based additive on fermentation quality, Clostridia and Saccharomyces cerevisiae in a white lupin-wheat silage. J. Sci. Food Agric. 2019, 99, 1492–1500. [Google Scholar] [CrossRef]
- Niklas, A.A.; Borge, G.I.A.; Rødbotten, R.; Berget, I.; Müller, M.H.B.; Herrmann, S.S.; Granby, K.; Kirkhus, B. Levels of nitrate, nitrite and nitrosamines in model sausages during heat treatment and in vitro digestion—The impact of adding nitrite and spinach (Spinacia oleracea L.). Food Res. Int. 2023, 166, 112595. [Google Scholar] [CrossRef]
- Wolff, I.A.; Wasserman, A.E. Nitrates, Nitrites, and Nitrosamines. Science 1972, 177, 15–19. [Google Scholar] [CrossRef]
- Srour, B.; Chazelas, E.; Fezeu, L.K.; Javaux, G.; Pierre, F.; Huybrechts, I.; Hercberg, S.; Deschasaux-Tanguy, M.; Kesse-Guyot, E.; Touvier, M. Nitrites, Nitrates, and Cardiovascular Outcomes: Are We Living “La Vie en Rose” with Pink Processed Meats? J. Am. Heart Assoc. 2022, 11, e027627. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, J.; He, Y.; Li, X.; Yin, S.; Chen, F.; Li, W. Association between Dietary Nitrite intake and Glioma Risk: A Systematic Review and Dose-Response Meta-Analysis of Observational Studies. Front. Oncol. 2022, 12, 910476. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, T.; Kube, K.; Gresner, N.; Südekum, K.H. Varying ensiling conditions affect the fermentation quality and abundance of bacterial key players in lucerne silages. J. Agric. Sci. 2020, 158, 297–303. [Google Scholar] [CrossRef]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Xia, L.; Qian, M.; Cheng, F.; Wang, Y.; Han, J.; Xu, Y.; Zhang, K.; Tian, J.; Jin, Y. The effect of lactic acid bacteria on lipid metabolism and flavor of fermented sausages. Food Biosci. 2023, 56, 103172. [Google Scholar] [CrossRef]
- Kim, S.-H.; Kang, K.H.; Kim, S.H.; Lee, S.; Lee, S.-H.; Ha, E.-S.; Sung, N.-J.; Kim, J.G.; Chung, M.J. Lactic acid bacteria directly degrade N-nitrosodimethylamine and increase the nitrite-scavenging ability in kimchi. Food Control 2017, 71, 101–109. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, Y. Effects of Lactic Acid Bacteria on Nitrite Degradation during Pickle Fermentation. Adv. Mater. Res. 2013, 781–784, 1656–1660. [Google Scholar] [CrossRef]
- Wei, T.; Mei, L.; Wang, Z.-G.; Xue, X.-H. Morphological and genetic responses of lactobacillus plantarum FQR to nitrite and its practical applications. J. Food Saf. 2017, 37, e12327. [Google Scholar] [CrossRef]
- Paik, H.D.; Lee, J.Y. Investigation of reduction and tolerance capability of lactic acid bacteria isolated from kimchi against nitrate and nitrite in fermented sausage condition. Meat Sci. 2014, 97, 609–614. [Google Scholar] [CrossRef]
- Merkle, A.C.; Lehnert, N. Binding and activation of nitrite and nitric oxide by copper nitrite reductase and corresponding model complexes. Dalton Trans. 2012, 41, 3355–3368. [Google Scholar] [CrossRef]
- Guo, H.; Saravanakumar, K.; Wang, M.-H. Total phenolic, flavonoid contents and free radical scavenging capacity of extracts from tubers of Stachys affinis. Biocatal. Agric. Biotechnol. 2018, 15, 235–239. [Google Scholar] [CrossRef]
- Rocha, B.S.; Gago, B.; Rui, M.B.; Laranjinha, J. Diffusion of nitric oxide through the gastric wall upon reduction of nitrite by red wine: Physiological impact. Nitric Oxide 2010, 22, 235–241. [Google Scholar] [CrossRef]
- Brandao, G.C.; Lima, D.C.; Ferreira, S.L.C. The chemical generation of NO for the determination of nitrite by high-resolution continuum source molecular absorption spectrometry. Talanta 2012, 98, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Johanningsmeier, S.D.; Price, R.; Reynolds, R.; Truong, V.-D.; Payton, S.C.; Breidt, F. Evaluation of nitrate and nitrite contents in pickled fruit and vegetable products. Food Control 2018, 90, 304–311. [Google Scholar] [CrossRef]
- Rather, I. Partially Purified Exopolysaccharide from Lactobacillus plantarum YML009 with Total Phenolic Content, Antioxidant and Free Radical Scavenging Efficacy. Indian J. Pharm. Educ. Res. 2015, 4, 282–292. [Google Scholar] [CrossRef]
- Yu, J.; Geng, Y.; Xia, H.; Ma, D.; Liu, C.; Wu, R.; Wu, J.; You, S.; Bi, Y. LAB Fermentation Improves Production of Bioactive Compounds and Antioxidant Activity of Withania somnifera Extract and Its Metabolic Signatures as Revealed by LC-MS/MS. J. Microbiol. Biotechnol. 2022, 32, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Valerio, F.; Di Biase, M.; Lattanzio, V.M.T.; Lavermicocca, P. Improvement of the antifungal activity of lactic acid bacteria by addition to the growth medium of phenylpyruvic acid, a precursor of phenyllactic acid. Int. J. Food Microbiol. 2016, 222, 1–7. [Google Scholar] [CrossRef]
- Sun, H.; Wang, H.; Zhang, A.; Yan, G.; Zhang, Y.; An, N.; Wang, X. Berberine Ameliorates Nonbacterial Prostatitis via Multi-Target Metabolic Network Regulation. OMICS J. Integr. Biol. 2015, 19, 186–195. [Google Scholar] [CrossRef]
- Guzmán-Guillén, R.; Prieto, A.I.; González, A.G.; Soria-Díaz, M.E.; Cameán, A.M. Cylindrospermopsin determination in water by LC-MS/MS: Optimization and validation of the method and application to real samples. Environ. Toxicol. Chem. 2012, 31, 2233–2238. [Google Scholar] [CrossRef]
- Wang, S.; Li, J.; Dong, Z.; Chen, L.; Yuan, X.; Shao, T. The effects of lactic acid bacteria strains isolated from various substrates on the fermentation quality of common vetch (Vicia sativa L.) in Tibet. Grass Forage Sci. 2018, 73, 639–647. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of Applying Lactic Acid Bacteria Isolated from Forage Crops on Fermentation Characteristics and Aerobic Deterioration of Silage. J. Dairy Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef]
- Griess, P. Bemerkungen zu der Abhandlung der HH. Weselsky und Benedikt “Ueber einige Azoverbindungen”. Berichte Dtsch. Chem. Ges. 1879, 12, 426–428. [Google Scholar] [CrossRef]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Kerr, K.G.; Rotowa, N.A.; Hawkey, P.M.; Lacey, R.W. Evaluation of the Mast ID and API 50CH systems for identification of Listeria spp. Appl. Environ. Microbiol. 1990, 56, 657–660. [Google Scholar] [CrossRef]
- Webster, J. The Biochemistry of Silage (Second Edition). By P. McDonald, A.R. Henderson and S. J. E. Heron. Marlow, Bucks, UK: Chalcombe Publications, (1991), pp. 340, £49.50, ISBN 0-948617-225. Exp. Agric. 1992, 28, 125. [Google Scholar] [CrossRef]
- Cai, Y.; Kumai, S.; Ogawa, M.; Benno, Y.; Nakase, T. Characterization and identification of Pediococcus species isolated from forage crops and their application for silage preparation. Appl. Environ. Microbiol. 1999, 65, 2901–2906. [Google Scholar] [CrossRef]
- Wang, S.C.; Chang, C.K.; Chan, S.C.; Shieh, J.S.; Chiu, C.K.; Duh, P.-D. Effects of lactic acid bacteria isolated from fermented mustard on lowering cholesterol. Asian Pac. J. Trop. Biomed. 2014, 4, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Shibukawa, T.; Ohira, Y.; Shimadzu, M.; Obata, E. Effect of pH on Nitrite Depletion Reaction. Kagaku Kogaku Ronbunshu 2009, 35, 613–617. [Google Scholar] [CrossRef]
- Xia, C.; Tian, Q.; Kong, L.; Sun, X.; Shi, J.; Zeng, X.; Pan, D. Metabolomics Analysis for Nitrite Degradation by the Metabolites of Limosilactobacillus fermentum RC4. Foods 2022, 11, 1009. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; de Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress Physiology of Lactic Acid Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef]
- Ye, Z.; Wang, S.; Zhang, C.; Zhao, Y. Coordinated Modulation of Energy Metabolism and Inflammation by Branched-Chain Amino Acids and Fatty Acids. Front. Endocrinol. 2020, 11, 617. [Google Scholar] [CrossRef]
- Yuan, W.; Piao, J.; Dong, Y. Advancements in the preparation methods of artificial cell membranes with lipids. Mater. Chem. Front. 2021, 5, 5233–5246. [Google Scholar] [CrossRef]
- Li, N.; Xu, C.; Li-Beisson, Y.; Philippar, K. Fatty acid and lipid transport in plant cells. Trends Plant Sci. 2016, 21, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, Y.; Woldemariam, K.Y.; Zhong, S.; Yu, Q.; Wang, J. Antioxidant effect of yeast on lipid oxidation in salami sausage. Front. Microbiol. 2023, 13, 1113848. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Y.; Liu, D.-M.; Jia, X.-Z.; Liang, M.-H.; Lu, Y.; Liu, J. Whole genome sequencing of Lactobacillus plantarum DMDL 9010 and its effect on growth phenotype under nitrite stress. LWT 2021, 149, 111778. [Google Scholar] [CrossRef]
- Poole, K. Stress responses as determinants of antimicrobial resistance in Gram-negative bacteria. Trends Microbiol. 2012, 20, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Hanking, B.M.; Roberts, S. Influence of Amino-acid Levels on Protein Synthesis in vitro. Nature 1964, 204, 1194–1195. [Google Scholar] [CrossRef] [PubMed]
- Jez, J.M. Connecting primary and specialized metabolism: Amino acid conjugation of phytohormones by GRETCHEN HAGEN 3 (GH3) acyl acid amido synthetases. Curr. Opin. Plant Biol. 2022, 66, 102194. [Google Scholar] [CrossRef]
- Kato, T.; Kikugawa, K. Proteins and amino acids as scavengers of nitrite: Inhibitory effect on the formation of nitrosodimethylamine and diazoquinone. Food Chem. Toxicol. 1992, 30, 617–626. [Google Scholar] [CrossRef]
- Fu, L.; Zhao, J.; Huang, J.; Li, N.; Dong, X.; He, Y.; Wang, W.; Wang, Y.; Qiu, J.; Guo, X. A mitochondrial STAT3-methionine metabolism axis promotes ILC2-driven allergic lung inflammation. J. Allergy Clin. Immunol. 2022, 149, 2091–2104. [Google Scholar] [CrossRef]
- Jin, P.; Ren, B.; Wang, X.C.; Jin, X.; Shi, X. Mechanism of microbial metabolic responses and ecological system conversion under different nitrogen conditions in sewers. Water Res. 2020, 186, 116312. [Google Scholar] [CrossRef]
- Zagórski, Z.P.; Sehested, K. Transients and stable radical from the deamination of α-alanine. J. Radioanal. Nucl. Chem. 1998, 232, 139–141. [Google Scholar] [CrossRef]
- Shi, J.; Xia, C.; Tian, Q.; Zeng, X.; Wu, Z.; Guo, Y.; Pan, D. Untargeted metabolomics based on LC–MS to elucidate the mechanism underlying nitrite degradation by Limosilactobacillus fermentum RC4. LWT 2022, 163, 113414. [Google Scholar] [CrossRef]
- Joshi, V.; Joung, J.-G.; Fei, Z.; Jander, G. Interdependence of threonine, methionine and isoleucine metabolism in plants: Accumulation and transcriptional regulation under abiotic stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef]
- Deber, C.M.; Stone, T.A. Relative role(s) of leucine versus isoleucine in the folding of membrane proteins. Pept. Sci. 2019, 111, e24075. [Google Scholar] [CrossRef]
- Miflin, B.J. The Location of Nitrite Reductase and Other Enzymes Related to Amino Acid Biosynthesis in the Plastids of Root and Leaves 1. Plant Physiol. 1974, 54, 550–555. [Google Scholar] [CrossRef]
- Palaiologos, G.; Hertz, L.; Schousboe, A. Role of aspartate aminotransferase and mitochondrial dicarboxylate transport for release of endogenously and exogenously supplied neurotransmitter in glutamatergic neurons. Neurochem. Res. 1989, 14, 359–366. [Google Scholar] [CrossRef]
- Weill-Thévnet, N.J.; Hermann, M.; Vandecasteele, J.-P. Lysine Transport Systems in Pseudomonas in Relation to Their Physiological Function. Microbiology 1979, 111, 263–269. [Google Scholar] [CrossRef]
- Xiao, J.; Liu, Q.-Y.; Du, J.-H.; Zhu, W.-L.; Li, Q.-Y.; Chen, X.-L.; Chen, X.-H.; Liu, H.; Zhou, X.-Y.; Zhao, Y.-Z.; et al. Integrated analysis of physiological, transcriptomic and metabolomic responses and tolerance mechanism of nitrite exposure in Litopenaeus vannamei. Sci. Total Environ. 2020, 711, 134416. [Google Scholar] [CrossRef]
- Tapia, M.I.; Liama, M.J.; Serra, J.L. Active glutamine synthetase is required for ammonium- or glutamine-promoted prevention of nitrate and nitrite reduction in the cyanobacterium Phormidium laminosum. Physiol. Plant 1995, 94, 241–246. [Google Scholar] [CrossRef]
- Lee, J.M.; Jang, W.J.; Lee, E.-W.; Kong, I.-S. β-glucooligosaccharides derived from barley β-glucan promote growth of lactic acid bacteria and enhance nisin Z secretion by Lactococcus lactis. LWT 2020, 122, 109014. [Google Scholar] [CrossRef]
- Bonestroo, M.H.; Kusters, B.J.M.; de Wit, J.C.; Rombouts, F.M. Glucose and sucrose fermenting capacity of homofermentative lactic acid bacteria used as starters in fermented salads. Int. J. Food Microbiol. 1992, 15, 365–376. [Google Scholar] [CrossRef]
- Xia, G.-H.; Wu, C.-R.; Zhang, M.-Z.; Yang, F.; Chen, C.; Hao, J. The metabolome and bacterial composition of high-moisture Italian ryegrass silage inoculated with lactic acid bacteria during ensiling. Biotechnol. Biofuels Bioprod. 2023, 16, 91. [Google Scholar] [CrossRef]
- Ma, S.; Guo, S.; Chen, J.; Sun, J.; Wang, Y.; Sheng, S. Enhancement of salt stressed-cucumber tolerance by application of glucose for regulating antioxidant capacity and nitrogen metabolism. Can. J. Plant Sci. 2020, 100, 253–263. [Google Scholar] [CrossRef]
- Li, Z.Y.; Ding, L.L.; Li, J.M.; Xu, B.L.; Yang, L.; Bi, K.S.; Wang, Z.T. ¹H-NMR and MS based metabolomics study of the intervention effect of curcumin on hyperlipidemia mice induced by high-fat diet. PLoS ONE 2015, 10, e0120950. [Google Scholar] [CrossRef] [PubMed]
- Suram, D.; Thatha, S.; Venkatapuram, P.; Adivireddy, P. Synthesis and Antimicrobial Activity of a New Class of Benzazolyl Pyrazoles. J. Heterocycl. Chem. 2017, 54, 3152–3162. [Google Scholar] [CrossRef]
- Johnson, E.M.; Richardson, M.D.; Warnock, D.W. In-vitro resistance to imidazole antifungals in Candida albicans. J. Antimicrob. Chemother. 1984, 13, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhang, Q.; Wang, Z.; Huang, G.; Li, S. Recent Advances in the Development of Indazole-based Anticancer Agents. Chemmedchem 2018, 13, 1490–1507. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-K.; Kim, S.-E.; Yeom, J.-H.; Ki, D.-W.; Lee, M.-S.; Song, J.-G.; Kim, Y.-S.; Seok, S.-J.; Yun, B.-S. Daldinan A, a novel isoindolinone antioxidant from the ascomycete Daldinia concentrica. J. Antibiot. 2012, 65, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Rajur, S.B.; Merwade, A.Y.; Basanagoudar, L.D.; Kulkarni, P.V. Synthesis and Antimicrobial Activity of New Substituted 10-Methyl-1,2-dihydro-1-oxo-1,2,4-triazino(4,5-a)indoles. J. Pharm. Sci. 1989, 78, 780–782. [Google Scholar] [CrossRef]
- Wang, Q.; Tu, J.; Yang, W.; Liang, T.; Liu, N.; Sheng, C. Discovery of Pyrazolone Carbothioamide Derivatives as Inhibitors of the Pdr1-KIX Interaction for Combinational Treatment of Azole-Resistant Candidiasis. J. Med. Chem. 2023, 66, 11893–11904. [Google Scholar] [CrossRef]
- Travis, N.; Kaiqi, S.; Julie, A.R.; Anren, S.; Tatsuro, Y.; Andrew, D.; Matthew, J.W.; Richard, O.F.; Robert, C.R.; Monika, D.; et al. Hypoxia modulates the purine salvage pathway and decreases red blood cell and supernatant levels of hypoxanthine during refrigerated storage. Haematologica 2018, 103, 361–372. [Google Scholar] [CrossRef]
- Lewis, E.B.; Chen, E.; Culyba, M.J. DNA cytosine methylation at the lexA promoter of Escherichia coli is stationary phase specific. G3 Genes|Genomes|Genetics 2021, 12, jkab409. [Google Scholar] [CrossRef] [PubMed]
- Holmsen, H.; Hindenes, J.O.; Fukami, M. Glycerophospholipid metabolism: Back to the future. Thromb. Res. 1992, 67, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.N.; Fan, T.W.-M. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [PubMed]
- Friedkin, M. Purine and pyrimidine metabolism in microorganisms. J. Cell. Comp. Physiol. 1953, 41, 261–282. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Zhang, J.; Huang, X.; Ma, M.; Zhao, K.; Yu, X.; Chen, Q.; Zhang, X.; Penttinen, P.; Gu, Y. Changes in the taxonomic and functional structures of microbial communities during vegetable waste mixed silage fermentation. Can. J. Microbiol. 2022, 68, 281–293. [Google Scholar] [CrossRef]
- Cunin, R.; Glansdorff, N.; Piérard, A.; Stalon, V. Biosynthesis and metabolism of arginine in bacteria. Microbiol. Rev. 1986, 50, 314–352. [Google Scholar] [CrossRef]
- Ling, J.; Reynolds, N.; Ibba, M. Aminoacyl-tRNA Synthesis and Translational Quality Control. Annu. Rev. Microbiol. 2009, 63, 61–78. [Google Scholar] [CrossRef]
- Miyamoto, T.; Homma, H. D-Amino acid metabolism in bacteria. J. Biochem. 2021, 170, 5–13. [Google Scholar] [CrossRef]
- Kobayashi, J. d-Amino Acids and Lactic Acid Bacteria. Microorganisms 2019, 7, 690. [Google Scholar] [CrossRef]
- Soutourina, J.; Plateau, P.; Blanquet, S. Metabolism of D-aminoacyl-tRNAs in Escherichia coli and Saccharomyces cerevisiae cells. J. Biol. Chem. 2000, 275, 32535–32542. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Yokoyama, S. Structures and functions of proteins and nucleic acids in protein biosynthesis. Int. Rev. Phys. Chem. 1989, 8, 125–145. [Google Scholar] [CrossRef]
- Park, S.G.; Kim, S. Do Aminoacyl-tRNA synthetases have biological functions other than in protein biosynthesis? IUBMB Life 2006, 58, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Chen, R.; Zhang, Y.; Zhu, Y.; Zhao, J.; Yao, H.; Ma, J. Porcine extraintestinal pathogenic Escherichia coli delivers two serine protease autotransporters coordinately optimizing the bloodstream infection. Front. Cell. Infect. Microbiol. 2023, 13, 1138801. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, J.; Sun, L.; Liu, M.; Dai, R.; Si, Q.; Ge, G.; Wang, Z.; Jia, Y. Mechanistic Insights into Nitrite Degradation by Metabolites of L. plantarum A50: An LC-MS-Based Untargeted Metabolomics Analysis. Fermentation 2024, 10, 92. https://doi.org/10.3390/fermentation10020092
An J, Sun L, Liu M, Dai R, Si Q, Ge G, Wang Z, Jia Y. Mechanistic Insights into Nitrite Degradation by Metabolites of L. plantarum A50: An LC-MS-Based Untargeted Metabolomics Analysis. Fermentation. 2024; 10(2):92. https://doi.org/10.3390/fermentation10020092
Chicago/Turabian StyleAn, Jiangbo, Lin Sun, Mingjian Liu, Rui Dai, Qiang Si, Gentu Ge, Zhijun Wang, and Yushan Jia. 2024. "Mechanistic Insights into Nitrite Degradation by Metabolites of L. plantarum A50: An LC-MS-Based Untargeted Metabolomics Analysis" Fermentation 10, no. 2: 92. https://doi.org/10.3390/fermentation10020092
APA StyleAn, J., Sun, L., Liu, M., Dai, R., Si, Q., Ge, G., Wang, Z., & Jia, Y. (2024). Mechanistic Insights into Nitrite Degradation by Metabolites of L. plantarum A50: An LC-MS-Based Untargeted Metabolomics Analysis. Fermentation, 10(2), 92. https://doi.org/10.3390/fermentation10020092