


Potential Use of Lactiplantibacillus plantarum BCC 4352 as a Functional Starter Culture for Fermenting Thai Pork Sausage (Nham)

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
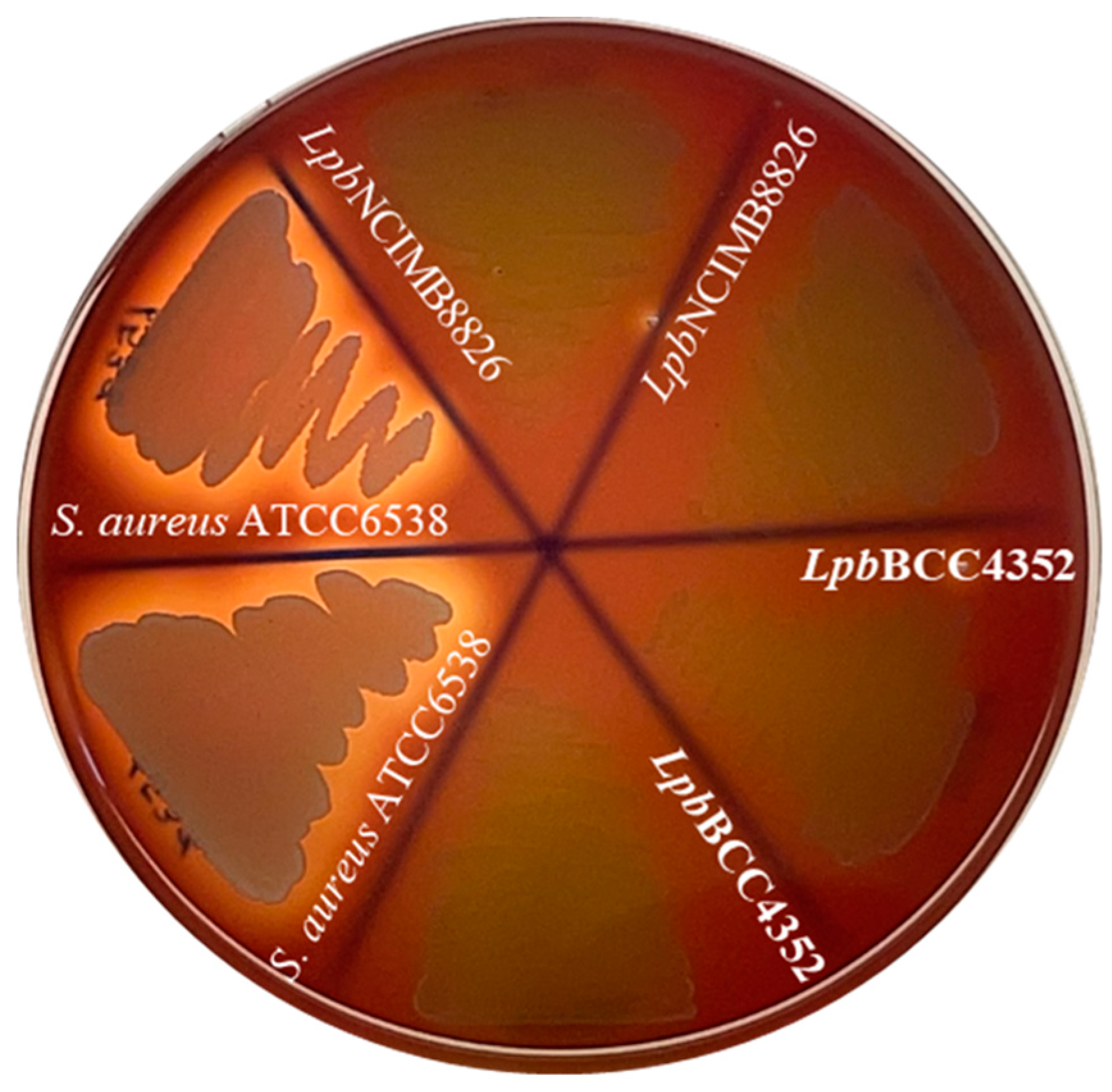
2.2. Determination of Antimicrobial Activity
2.3. Determination of Acid and Bile Tolerance
2.4. Resistance to the Simulated Digestion Conditions
2.5. Detection of the Bile Salt Hydrolase Activity
2.6. Adhesion Assay
2.6.1. Adhesion to the Human Colon Adenocarcinoma Cell Line (Caco-2)
2.6.2. Mucin Adhesion Assay
2.6.3. Detection of Binding-Related Genes
2.7. Growth of LpbBCC4352 in Prebiotic Medium
2.8. Analysis of Short Chain Fatty Acids (SCFAs)
2.9. Cholesterol Assimilation Assay
2.10. Safety Evaluation of LpBCC4352
2.10.1. Hemolytic Activity Assay
2.10.2. Antibiotic Susceptibility Test
2.11. Evaluating the Performance of LpbBCC4352 as a Starter Culture in Nham Production
2.11.1. Starter Culture Preparation
2.11.2. Nham Preparation
2.11.3. Determination of Physicochemical Properties of Nham
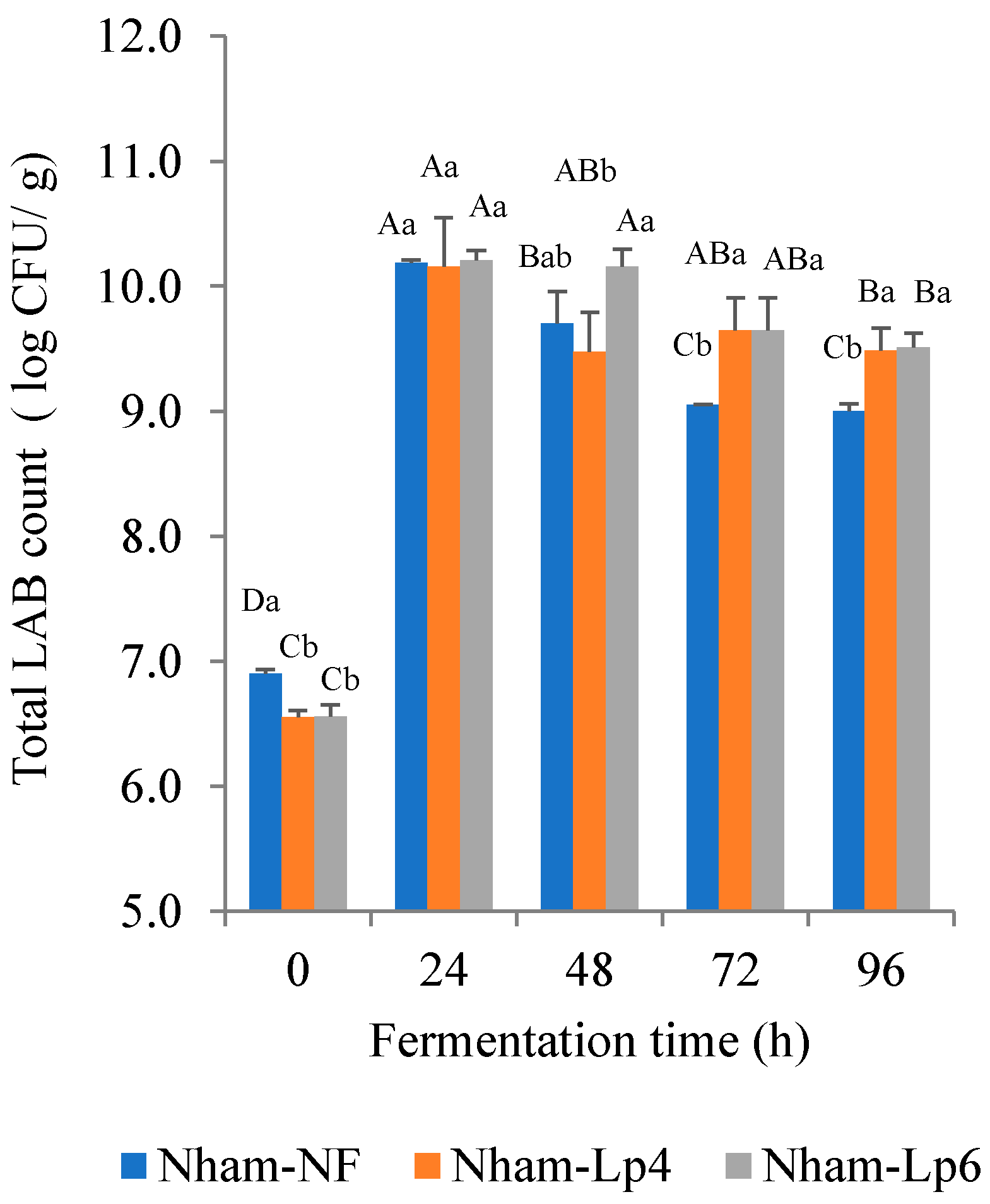
2.11.4. Microbiological Analysis of Nham
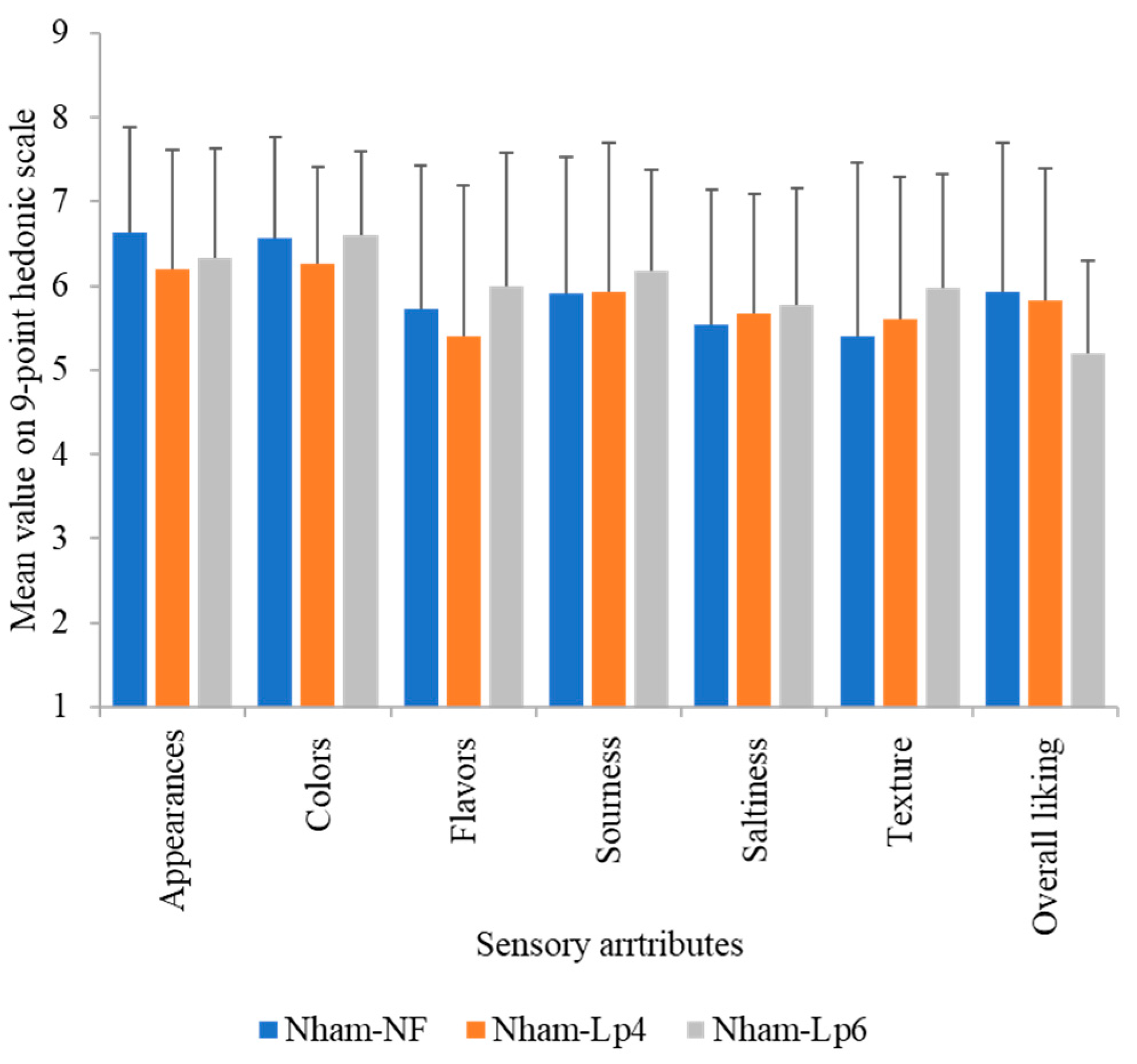
2.11.5. Sensory Analysis
2.12. Statistical Analysis
3. Results and Discussion
3.1. Antimicrobial Activity
3.2. Resistance to pH, Bile Salts, and the Simulated Digestive Conditions
3.3. Adhesion to Caco-2 Cells and Mucin
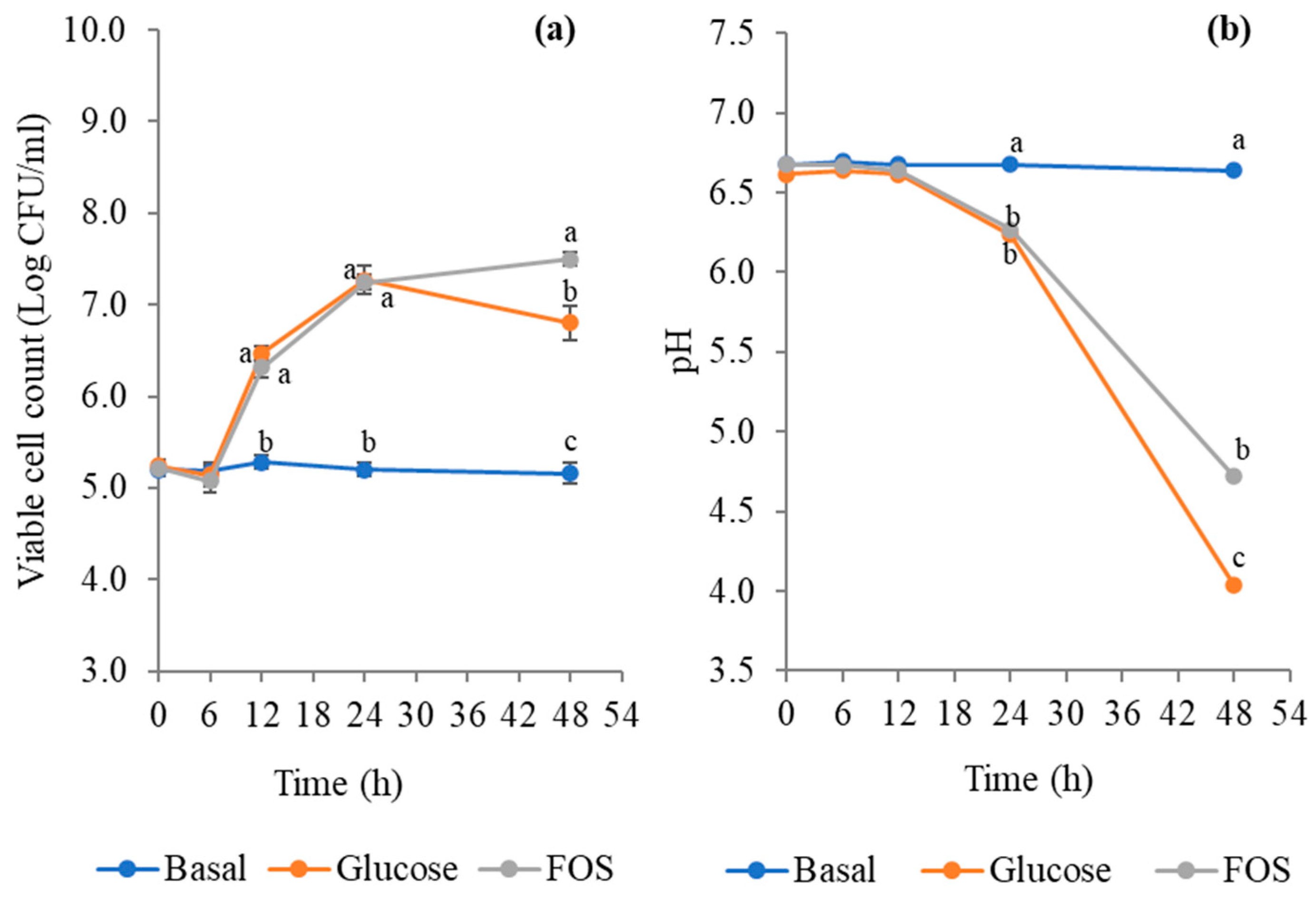
3.4. Growth in the Prebiotic Medium
3.5. Cholesterol Assimilation
3.6. Hemolytic Activity
3.7. Antibiotics Susceptibility Test
3.8. Effect of LpbBCC4352 on the Fermentation Characteristics of Nham
3.9. Acceptability Testing of the Finished Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- 2001. Available online: http://www.who.int/foodsafety/publications/fs_management/en/probiotics.pdf (accessed on 16 January 2023).
- Pieniz, S.; Andreazza, R.; Anghinoni, T.; Camargo, F.; Brandelli, A. Probiotic potential, antimicrobial and antioxidant activities of Enterococcus durans strain LAB18s. Food Control 2014, 37, 251–256. [Google Scholar] [CrossRef]
- Li, C.; Nie, S.P.; Ding, Q.; Zhu, K.X.; Wang, Z.J.; Xiong, T.; Gong, J.; Xie, M.-Y. Cholesterol-lowering effect of Lactobacillus plantarum NCU116 in a hyperlipidemic rat model. J. Funct. Foods 2014, 8, 340–347. [Google Scholar] [CrossRef]
- Hernández-Hernández, O.; Muthaiyan, A.; Moreno, F.J.; Montilla, A.; Sanz, M.L.; Ricke, S.C. Effect of prebiotic carbohydrates on the growth and tolerance of Lactobacillus. Food Microbiol. 2012, 30, 355–361. [Google Scholar] [CrossRef]
- Ji, Y.; Li, J.; Qin, Z.; Li, A.; Gu, Z.; Liu, X.; Lin, L.; Zhou, Y. Contribution of nuclease to the pathogenesis of Aeromonas hydrophila. Virulence 2015, 6, 515–522. [Google Scholar] [CrossRef]
- Rodgers, S. Novel applications of live bacteria in food services: Probiotics and protective cultures. Trends Food Sci. Technol. 2008, 19, 188–197. [Google Scholar] [CrossRef]
- Buntin, N.; Chanthachum, S.; Hongpattarakere, T. Screening of lactic acid bacteria from gastrointestinal tracts of marine fish for their potential use as probiotics. Songklanakarin J. Sci. Technol. 2008, 30 (Suppl. S1), 141–148. [Google Scholar]
- De Vos, P.; Marijke, M.F.; Milica, S.; Jan, S. Encapsulation for preservation of functionality and targeted delivery of bioactive food components. Int. Dairy J. 2010, 20, 292–302. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Ouwehand, A.C.; Isolauri, E. Clinical applications of probiotic bacteria. Int. Dairy J. 1998, 8, 563–572. [Google Scholar] [CrossRef]
- Borriello, S.P.; Hammes, W.P.; Holzapfel, W.; Marteau, P.; Schrezenmeir, J.; Vaara, M.; Valtonen, V. Safety of probiotics that contain lactobacilli or bifidobacteria. Clin. Infect. Dis. 2003, 36, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Bernardeau, M.; Vernoux, J.P.; Henri-Dubernet, S.; Guéguen, M. Safety assessment of dairy microorganisms: The Lactobacillus genus. Int. J. Food Microbiol. 2008, 126, 278–285. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on the maintenance of the list of QPS biological agents intentionally added to food and feed. EFSA J. 2012, 10, 3020. [Google Scholar] [CrossRef]
- Salminen, S.; Playne, M.; Lee, Y.K. Successful probiotic lactobacilli: Human studies on probiotic efficacy. In Handbook of Functional Dairy Products; Shortt, C., O’Brien, J., Eds.; CRC Press: New York, NY, USA, 2004. [Google Scholar]
- Ngamsomchat, A.; Kaewkod, T.; Konkit, M.; Tragoolpua, Y.; Bovonsombut, S.; Chitov, T. Characterisation of Lactobacillus plantarum of dairy-product origin for probiotic chèvre cheese production. Foods 2022, 11, 934. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Wen, J.; Wu, Z.; Pan, D.; Wang, L. Evaluation of probiotic yoghurt by the mixed culture with Lactobacillus plantarum A3. Food Sci. Hum. Wellness 2022, 11, 323–331. [Google Scholar] [CrossRef]
- El-Sayed, M.I.; Aly, E.; El-Dee, A.M. Improving the physicochemical and antioxidative properties of fermented goat milk using carob molasses and some probiotic Strains. Food Sci. Biotechnol. 2023, 33, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Valyasevi, R.; Rolle, R.S. An overview of small-scale food fermentation technologies in developing countries with special reference to Thailand: Scope for their improvement. Int. J. Food Microbiol. 2002, 75, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Visessanguan, W.; Benjakul, S.; Smitinont, T.; Kittikun, C.; Thepkasikul, P.; Panya, A. Changes in microbiological, biochemical and physical-chemical properties of Nham inoculated with different inoculum levels of Lactobacillu curvatus. LWT-Food Sci. Technol. 2006, 39, 814–826. [Google Scholar] [CrossRef]
- Kingcha, Y.; Tosukhowong, A.; Zendo, T.; Roytrakul, S.; Luxananil, P.; Chareonpornsook, K.; Valyasevi, R.; Sonomoto, K.; Visessanguan, W. Anti-listeria activity of Pediococcus pentosaceus BCC 3772 and application as starter culture for Nham, a traditional fermented pork sausage. Food Control 2012, 25, 190–196. [Google Scholar] [CrossRef]
- Ba, H.V.; Seo, H.W.; Seong, P.N.; Kang, S.M.; Kim, Y.S.; Cho, S.-H.; Park, B.-Y.; Ham, J.-S.; Kim, J.-H. Lactobacillus plantarum (KACC 92189) as a Potential Probiotic Starter Culture for Quality Improvement of Fermented Sausages. Korean J. Food Sci. Anim. Resour. 2018, 38, 189–202. [Google Scholar] [CrossRef]
- Beganović, J.; Pavunc, A.L.; Gjuračić, K.; Špoljarec, M.; Šušković, J.; Kos, B. Improved sauerkraut production with probiotic strain Lactobacillus plantarum L4 and Leuconostoc mesenteroides LMG 7954. J. Food Sci. 2011, 76, M124–M129. [Google Scholar] [CrossRef]
- Islam, S.; Biswas, S.; Jabin, T.; Moniruzzaman, M.; Biswas, J.; Uddin, S.; Ekram, A.E.; Elgorban, A.M.; Ghodake, G.; Syed, A.; et al. Probiotic potential of Lactobacillus plantarum DMR14 for preserving and extending shelf life of fruits and fruit juice. Heliyon 2023, 9, e17382. [Google Scholar] [CrossRef]
- Atsadawut, A.; Apinun, W.; Jantima, T.; Dolnapa, K.; Chiu-Hsia, C. Potential of infrared drying and cell-protective agent efficiency on survival of Lactobacillus plantarum probiotic in fermented soybean meal. Biocatal. Agric. Biotechnol. 2023, 53, 102843. [Google Scholar] [CrossRef]
- Baruzzi, F.; Poltronieri, P.; Quero, G.M.; Morea, M.; Morelli, L. An in vitro protocol for direct isolation of potential probiotic lactobacilli from raw bovine milk and traditional fermented milks. Appl. Microbiol. Biotechnol. 2011, 90, 331–342. [Google Scholar] [CrossRef]
- Zago, M.; Fornasari, M.E.; Carminati, D.; Burns, P.; Suàrez, V.; Vinderola, G.; Reinheimer, J.; Giraffa, G. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol. 2011, 28, 1033–1040. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Jofré, A.; Aymerich, T.; Garriga, M. Probiotic fermented sausages: Myth or reality? Procedia Food Sci. 2015, 5, 133–136. [Google Scholar] [CrossRef]
- Trabelsi, I.; Slima, S.B.; Ktari, N.; Triki, M.; Abdehedi, R.; Abaza, W.; Moussa, H.; Abdeslam, A.; Salah, R.B. Incorporation of probiotic strain in raw minced beef meat: Study of textural modification, lipid and protein oxidation and color parameters during refrigerated storage. Meat Sci. 2019, 154, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Blaiotta, G.; Murru, N.; Di Cerbo, A.; Romano, R.; Aponte, M. Production of probiotic bovine salami using Lactobacillus plantarum 299v as adjunct. J. Sci. Food Agric. 2018, 98, 2285–2294. [Google Scholar] [CrossRef] [PubMed]
- Pavli, F.G.; Argyri, A.A.; Chorianopoulos, N.G.; Nychas, G.J.E.; Tassou, C.C. Effect of Lactobacillus plantarum L125 strain with probiotic potential on physicochemical, microbiological, and sensorial characteristics of dry-fermented sausages. LWT 2020, 118, 108810. [Google Scholar] [CrossRef]
- Cavalheiro, C.P.; Ruiz-Capillas, C.; Herrero, A.M.; Jiménez-Colmenero, F.; Pintado, T.; de Menezes, C.R.; Fries, L.L.M. Effect of encapsulated Lactobacillus plantarum as probiotic on dry sausages during chilled storage. Int. J. Food Sci. Technol. 2020, 55, 3613–3621. [Google Scholar] [CrossRef]
- Sirini, N.; Loyeau, P.; Ruiz, M.; Stegmayer, M.; Soto, L.; Werning, M.; Frizzo, L.; Ordoñez, V.; Fernández-López, J.; Rosmini, M. Development of probiotic fermented sausages and viability monitoring of supplemented Lactiplantibacillus plantarum BFL strain. Fermentation 2022, 8, 526. [Google Scholar] [CrossRef]
- Klingberg, T.D.; Budde, B.B. The survival and persistence in the human gastrointestinal tract of five potential probiotic lactobacilli consumed as freeze-dried cultures or as probiotic sausage. Int. J. Food Microbiol. 2006, 109, 157–159. [Google Scholar] [CrossRef]
- Tanasupawat, S.; Komagata, K. Lactic acid bacteria in fermented foods in Thailand. World J. Microbiol. Biotechnol. 1995, 11, 253–256. [Google Scholar] [CrossRef]
- Visessanguan, W.; Benjakul, S.; Riebroy, S.; Thepkasikul, P. Changes in composition and functional properties of proteins and their contributions to Nham characteristics. Meat Sci. 2004, 66, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Phupaboon, S.; Kontongdee, P.; Hashim, F.J.; Kanpipit, N.; Matra, M.; Totakul, P.; Prommachart, R.; Phesatcha, B.; Wanapat, M. Bioaccessibility and Microencapsulation of Lactobacillus sp. to Enhance Nham Protein Hydrolysates in Thai Fermented Sausage. Foods 2022, 11, 3846. [Google Scholar] [CrossRef] [PubMed]
- Zendo, T.; Fukao, M.; Ueda, K.; Higuchi, T.; Nakayama, J.; Sonomoto, K. Identification of the lantibiotic nisin Q, a new natural nisin variant produced by Lactococcus lactis 61-14 isolated from a river in Japan. Biosci. Biotechnol. Biochem. 2003, 67, 1616–1619. [Google Scholar] [CrossRef]
- Vijayakumar, M.; Ilavenil, S.; Kim, D.H.; Arasu, M.V.; Priya, K.; Choi, K.C. In vitro assessment of the probiotic potential of Lactobacillus plantarum KCC-24 isolated from Italian rye-grass (Lolium multiflorum) forage. Anaerobe 2015, 32, 90–97. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardized static in vitro digestion method suitable for food–an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Allain, T.; Chaouch, S.; Thomas, M.; Travers, M.A.; Valle, I.; Langella, P.; Grellier, P.; Polack, B.; Florent, I.; Bermúdez-Humarán, L.G. Bile salt hydrolase activities: A novel target to screen anti-Giardia lactobacilli. Front. Microbiol. 2018, 9, 89. [Google Scholar] [CrossRef]
- Klingberg, T.D.; Axelsson, L.; Naterstad, K.; Elsser, D.; Budde, B.B. Identification of potential probiotic starter cultures for Scandinavian-type fermented sausages. Int. J. Food Microbiol. 2005, 105, 419–431. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Grootaert, C.; Possemiers, S.; Verstraete, W.; Verbeken, K.; Van de Wiele, T. In vitro model to study the modulation of the mucin-adhered bacterial community. Appl. Microbiol. Biotechnol. 2009, 83, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Turpin, W.; Humblot, C.; Noordine, M.L.; Thomas, M.; Guyot, J.P. Lactobacillaceae and Cell Adhesion: Genomic and Functional Screening. PLoS ONE 2012, 7, e38034. [Google Scholar] [CrossRef]
- Gibson, G.R.; Cummings, J.H.; Macfarlane, G.T. Use of a three-stage continuous culture system to study the effect of mucin on dissimilatory sulfate reduction and methanogenesis by mixed populations of human gut bacteria. Appl. Environ. Microbiol. 1988, 54, 2750–2755. [Google Scholar] [CrossRef]
- Gamonpilas, C.; Buathongjan, C.; Sangwan, W.; Rattanaprasert, M.; Weizman, K.C.; Klomtun, M.; Phonsatta, N.; Methacanon, P. Production of low molecular weight pectins via electron beam irradiation and their potential prebiotic functionality. Food Hydrocoll. 2021, 113, 106551. [Google Scholar] [CrossRef]
- Schooley, D.L.; Kubiak, F.M.; Evans, J.V. Capillary gas chromatographic analysis of volatile and non-volatile organic acids from biological samples as the t-butyldimethylsilyl derivatives. J. Chromatogr. Sci. 1985, 23, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Liong, M.T.; Shah, N.P. Optimization of cholesterol removal by probiotics in the presence of prebiotics by using a response surface method. Appl. Environ. Microbiol. 2005, 71, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Rudel, L.L.; Morris, M.D. Determination of cholesterol using O-phthalaldehyde. J. Lipid Res. 1973, 14, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Speck, S.; Wenke, C.; Feßler, A.T.; Kacza, J.; Geber, F.; Scholtzek, A.D.; Hanke, D.; Eichhorn, I.; Schwarz, S.; Rosolowski, M.; et al. Borderline resistance to oxacillin in Staphylococcus aureus after treatment with sub-lethal sodium hypochlorite concentrations. Heliyon 2020, 6, e04070. [Google Scholar] [CrossRef] [PubMed]
- Vesa, T.; Pochart, P.; Marteau, P. Pharmacokinetics of Lactobacillus plantarum NCIMB 8826, Lactobacillus fermentum KLD, and Lactococcus lactis MG 1363 in the human gastrointestinal tract. Aliment. Pharmacol. Ther. 2000, 14, 823–828. [Google Scholar] [CrossRef] [PubMed]
- ISO 10932:2010; Milk and Milk Products-Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-Enterococcal Lactic Acid Bacteria (LAB). International Organization for Standardization: Geneva, Switzerland, 2010.
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis International, 17th ed.; AOAC: Washington, DC, USA, 2000. [Google Scholar]
- Sharma, G. Digital color imaging. IEEE Trans. Image Process. 1997, 6, 901–932. [Google Scholar] [CrossRef] [PubMed]
- Nespolo, C.R.; Brandelli, A. Production of bacteriocin-like substances by lactic acid bacteria isolated from regional ovine cheese. Braz. J. Microbiol. 2010, 41, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Bezares, B.; Saenz, Y.; Navarro, L.; Zarazaga, M.; Ruiz-Larrea, F.; Torres, C. Coculture-inducible bacteriocin activity of Lactobacillus plantarum strain J23 isolated from grape must. Food Microbiol. 2007, 24, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Tabasco, R.; García-Cayuela, T.; Peláez, C.; Requena, T. Lactobacillus acidophilus La-5 increases lactacin B production when it senses live target bacteria. Int. J. Food Microbiol. 2009, 132, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; Calasso, M.; Vincentini, O.; Vernocchi, P.; Ndagijimana, M.; De Vincenzi, M.; Dessì, M.R.; Guerzoni, M.E.; Gobbetti, M. Quorum sensing in sourdough Lactobacillus plantarum DC400: Induction of plantaricin A (PlnA) under co-cultivation with other lactic acid bacteria and effect of PlnA on bacterial and Caco-2 cells. Proteomics 2010, 10, 2175–2190. [Google Scholar] [CrossRef]
- Maldonado-Barragán, A.; Caballero-Guerrero, B.; Luis Ruiz-Barba, J. Induction of bacteriocin production by coculture is widespread among plantaricin-producing Lactobacillus plantarum strains with different regulatory operons. Food Microbiol. 2013, 33, 40–47. [Google Scholar] [CrossRef]
- Ding, T.; Li, Y. Study on the promotive effect and mechanism of exogenous nucleotides on Lactobacillus casei antagonism against Salmonella enterica by coculture. LWT 2023, 182, 114821. [Google Scholar] [CrossRef]
- Fongcom, A.; Pruksakorn, S.; Mongkol, R.; Tharavichitkul, P.; Yoonim, N. Streptococcus suis infection in northern Thailand. J. Med. Assoc. Thail. 2001, 84, 1502–1508. [Google Scholar]
- Khadthasrima, N.; Hannwong, T.; Thammawitjaya, P.; Pingsusean, D.; Akkanij, B.; Jaikhar, A.; Paungmali, P.; Yudee, P.; Wongyai, S.; Samerchea, S.; et al. Human Streptococcus suis Outbreak in Phayao Province, Thailand, 2007. Outbreak Surveill. Investig. Rep. (OSIR) e-J. 2008, 1, 4–7. Available online: http://www.osirjournal.net/issue.php?id=3 (accessed on 16 January 2023). [CrossRef]
- Navacharoen, N.; Chantharochavong, V.; Hanprasertpong, C.; Kangsanarak, J.; Lekagul, S. Hearing and vestibular loss in Streptococcus suis infection from swine and traditional raw pork exposure in northern Thailand. J. Laryngol. Otol. 2009, 123, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Noppon, B.; Khaeng-air, S.; Sopa, A.; Phuaram, P.; Wongsan, R.; Laohasinnurak, T. Streptococcus suis serotype 2 in Uncooked Pork Meat Products in Khon Kaen, Northeastern Thailand, and their Antimicrobial Profiles. Int. J. Sci. Eng. Res. 2014, 5, 1130–1133, ISSN 2229-5518. [Google Scholar]
- Burintramart, U.; Chantarasakha, K.; Chokesajjawatee, N. Diversity of lactic acid bacteria in Thai fermented pork (Nham) during fermentation assessed by Denaturing Gradient Gel Electrophoresis. KKU Res. J. 2014, 19, 19–25. [Google Scholar]
- Wongnak, P.; Wiratsudakul, A.; Nuanualsuwan, S. A risk assessment of pathogenic Streptococcus suis in pork supply chains and markets in Thailand. Food Control 2020, 118, 1071432. [Google Scholar] [CrossRef]
- Schillinger, U.; Guigas, C.; Holzapfel, W.H. In vitro adherence and other properties of lactobacilli used in probiotic yoghurt-like products. Int. Dairy J. 2005, 15, 1289–1297. [Google Scholar] [CrossRef]
- Nannen, N.L.; Hutkins, R.W. Proton-translocating adenosine triphosphate activity in lactic acid bacteria. J. Dairy Sci. 1991, 74, 747–751. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef] [PubMed]
- Jaichumjai, P.; Valyasevi, R.; Assavanig, A.; Kurdi, P. Isolation and characterization of acid-sensitive Lactobacillus plantarum with application as starter culture for Nham production. Food Microbiol. 2010, 27, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Tian, X.; Huang, R.; Tao, X.; Shah, N.P.; Wei, H.; Wan, C. A physiological comparative study of acid tolerance of Lactobacillus plantarum ZDY 2013 and L. plantarum ATCC 8014 at membrane and cytoplasm levels. Ann. Microbiol. 2017, 67, 669–677. [Google Scholar] [CrossRef]
- Noriega, L.; Gueimonde, M.; Sánchez, B.; Margolles, A.; delosReyes-Gavilán, C.G. Effect of the adaptation to high bile salts concentrations on glycosidic activity, survival at low pH and cross-resistance to bile salts in Bifidobacteirum. Int. J. Food Microbiol. 2004, 94, 79–86. [Google Scholar] [CrossRef]
- Kheroua, O.; Belleville, J. Behaviour of digestive enzymes in the pancreatic juice and pancreas of rats fed on a low-protein diet than on a balanced diet. Reprod. Nutr. Dev. 1981, 21, 901–917. [Google Scholar] [CrossRef]
- De Smet, I.; Van Hoorde, L.; Vande Woestyne, M.; Christiaens, H.; Verstraete, W. Significance of bile salt hydrolytic activities of Lactobacilli. J. Appl. Bacteriol. 1995, 79, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Moser, S.A.; Savage, D.C. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in Lactobacilli. Appl. Environ. Microbiol. 2001, 67, 3476–3480. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Gahan, C.G.M.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [PubMed]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C.; et al. In vitro selection criteria for probiotic bacteria of human origin: Correlation with in vivo findings. Am. J. Clin. Nutr. 2001, 73, 386s–392s. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Surono, I.; Meriluoto, J.; Salminen, S. Indigenous dadih lactic acid bacteria: Cell-surface properties and interactions with pathogens. J. Food Sci. 2007, 72, M89–M93. [Google Scholar] [CrossRef] [PubMed]
- Bergillos-Meca, T.; Costabile, A.; Walton, G.; Moreno-Montoro, M.; Ruiz-Bravo, A.; Ruiz-López, M.D. In vitro evaluation of the fermentation properties and potential probiotic activity of Lactobacillus plantarum C4 in batch culture systems. LWT-Food Sci. Technol. 2015, 60, 420–426. [Google Scholar] [CrossRef]
- Buck, B.L.; Altermann, E.; Svingerud, T.; Klaenhammer, T.R. Functional analysis of putative adhesion factors in Lactobacillus acidophilus NCFM, Appl. Environ. Microbiol. 2005, 71, 8344–8351. [Google Scholar] [CrossRef]
- Ashida, N.; Yanagihara, S.; Shinoda, T.; Yamamoto, N. Characterization of adhesive molecule with affinity to Caco-2 cells in Lactobacillus acidophilus by proteome analysis. J. Biosci. Bioeng. 2011, 112, 333–337. [Google Scholar] [CrossRef]
- Yuille, S.; Reichardt, N.; Panda, S.; Dunbar, H.; Mulder, I.E. Human gut bacteria as potent class I histone deacetylase inhibitors in vitro through production of butyric acid and valeric acid. PLoS ONE 2018, 13, e0201073. [Google Scholar] [CrossRef]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef]
- Lye, H.; Khoo, B.Y.; Karim, A.A.; Rusul, G.; Liong, M.T. Growth properties and cholesterol removal ability of electroporated Lactobacillus acidophilus BT 1088. J. Microbiol. Biotechnol. 2012, 22, 981–989. [Google Scholar] [CrossRef]
- Kingkaew, E.; Konno, H.; Hosaka, Y.; Phongsopitanun, W.; Tanasupawat, S. Characterization of lactic acid bacteria from fermented fish (pla-paeng-daeng) and their cholesterol-lowering and immunomodulatory effects. Microbes Environ. 2023, 38, ME22044. [Google Scholar] [CrossRef]
- Lin, M.Y.; Chen, T.W. Reduction of Cholesterol by Lactobacillus acidophilus in Culture Broth. J. Food Drug Anal. 2000, 8, 4. [Google Scholar] [CrossRef]
- Lye, H.S.; Rahmat-Ali, G.R.; Liong, M.T. Mechanisms of cholesterol removal by lactobacilli under conditions that mimic the human gastrointestinal tract. Int. Dairy J. 2010, 20, 169–175. [Google Scholar] [CrossRef]
- Lye, H.S.; Rusul, G.; Liong, M.T. Removal of cholesterol by lactobacilli via incorporation and conversion to coprostanol. J. Dairy Sci. 2010, 93, 1383–1392. [Google Scholar] [CrossRef]
- Kriaa, A.; Bourgin, M.; Potiron, A.; Mkaouar, H.; Jablaoui, A.; Gérard, P.; Maguin, E.; Rhimi, M. Microbial impact on cholesterol and bile acid metabolism: Current status and future prospects. J. Lipid Res. 2019, 60, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Liong, M.T.; Shah, N.P. Effects of a Lactobacillus casei synbiotic on serum lipoprotein, intestinal microflora, and organic acids in rats. J. Dairy Sci. 2006, 89, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.M.; Bongers, R.S.; de Vos, W.M.; Kleerebezem, M. Functional analysis of four bile salt hydrolase and penicillin acylase family members in Lactobacillus plantarum WCFS1. Appl. Environ. Microbiol. 2008, 74, 4719–4726. [Google Scholar] [CrossRef] [PubMed]
- De Preter, V.; Vanhoutte, T.; Huys, G.; Swings, J.; De Vuyst, L.; Rutgeerts, P.; Verbeke, K. Effects of Lactobacillus casei Shirota, Bifidobacterium breve, and oligofructose-enriched inulin on colonic nitrogen-protein metabolism in healthy humans. Am. J. Physiol.-Gastrointest. Liver Physiol. 2007, 292, G358–G368. [Google Scholar] [CrossRef] [PubMed]
- Osmanagaoglu, O.; Kiran, F.; Ataoglu, H. Evaluation of in vitro probiotic potential of Pediococcus pentosaceus OZF isolated from human breast milk. Probiotics Antimicrob. Proteins 2010, 2, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, D.; Liu, S.; Liu, Y. Probiotic potential of L. sake C2 isolated from traditional Chinese fermented cabbage. Eur. Food Res. Technol. 2012, 234, 45–51. [Google Scholar] [CrossRef]
- FAO/WHO. Joint FAO/WHO Working Group Report on Draft Guidelines for the Evaluation of Probiotics in Food; FAO/WHO: London, ON, Canada, 2002. Available online: https://www.mhlw.go.jp/file/05-Shingikai-11121000-Iyakushokuhinkyoku-Soumuka/0000197343.pdf (accessed on 16 January 2023).
- EFSA. Update of the criteria used in the assessment of bacterial resistance to antibiotics of human or veterinary importance. EFSA J. 2008, 6, 732. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator Strains | Antagonistic Activity | |
|---|---|---|
| Cell-to-Cells | Cells Free Supernatant (CFS) | |
| Gram-positive bacteria | ||
| Bacillus circulans JCM 2504T | - | - |
| Bacillus coagulans JCM 2257T | - | - |
| Bacillus subtilis ssp. subtilis JCM 1465T | - | - |
| Enterococcus faecalis JCM 5803T | - | - |
| Enterococcus faecium JCM 5804T | - | - |
| Kocuria rhizophila NBRC 12708 | + | - |
| Lactiplantibacillus plantarum ATCC 14917T | + | - |
| Lactiplantibacillus plantarum NCIMB 8826 | - | - |
| Latilactobacillus sakei ssp. sakei JCM 1157T | + | - |
| Lactococcus garvieae BCC 43578 | - | - |
| Leuconostoc mescenteroides ssp. mes JCM. 6124T | - | - |
| Listeria innocua ATCC 33090T | - | - |
| Pediococcus dextrinicus JCM 5887T | - | - |
| Pediococcus pentosaceus JCM 5885 | + | - |
| Staphylococcus aureus ATCC 23235 | - | - |
| Staphylococcus aureus ATCC 25923 | - | - |
| Staphylococcus aureus ATCC 6512 | - | - |
| Staphylococcus aureus ATCC 29213 | - | - |
| Streptococcus gordonii DMST 35778 | - | - |
| Streptococcus mutan DMST 18777 | - | - |
| Streptococcus pyrogenes DMST 17020 | - | - |
| Streptococcus suis P1/7 * | + | - |
| Weissella paramesenteroides JCM 9890T | - | - |
| Propionibacterium acnes ATCC 11827 | - | - |
| Propionibacterium acnes DSM 1897 | - | - |
| Propionibacterium acnes DSM 16379 | - | - |
| Propionibacterium acnes JCM 6473 | - | - |
| Gram-negative bacteria | ||
| Escherichia coli ATCC 25922 | - | - |
| Escherichia coli O157:H7 ATCC 43888 | - | - |
| Salmonella Typhimurium ATCC 13311 | - | - |
| Strains | Binding-Related Genes | References | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| apf | cbsA | cnb | fpbA | mapA | gtf | msa | mub1 | mub2 | slpA | ||
| L. plantarum BCC4352 (LpbBCC4352) | + | - | + | + | + | + | + | - | + | + | This study |
| L. plantarum NCIMB 8826 (LpbNCIMB8826) | + | + | + | + | + | + | + | + | + | - | This study |
| L. plantarum 11.3 | + | - | + | + | + | - | + | + | + | - | [45] |
| L. plantarum 2.1 | + | - | - | + | + | - | - | + | + | - | [45] |
| L. plantarum A6 | + | - | + | + | + | - | + | + | + | - | [45] |
| Samples | Residual Cholesterol (μg/mL) | Cholesterol Reduction (μg/mL) | Cholesterol Assimilation (%) | |
|---|---|---|---|---|
| 0 h | 72 h | |||
| LpbNCIMB8826 | 20.70 ± 0.92 a | 9.14 ± 1.21 a | 12.20 ± 1.21 a | 55.85 ± 7.80 a |
| LpbBCC4352 | 20.16 ± 0.80 a | 11.64 ± 0.80 b | 9.09 ± 0.80 b | 42.27 ± 1.69 b |
| Antimicrobials | Drug Class | Test Range (mg/L) | MIC (mg/L) | Interpretation | |
|---|---|---|---|---|---|
| Cut Off [97] | LpbBCC4352 | ||||
| Ampicillin | Beta-lactam | 0.06–8 | 2 | 0.5 | Susceptible |
| Chloramphenicol | Phenicol | 0.25–32 | 8 | 8 | Susceptible |
| Clindamycin | Lincomycin | 0.13–16 | 4 | <0.13 | Susceptible |
| Erythromycin | Macrolide | 0.13–16 | 1 | 1 | Susceptible |
| Gentamicin | Aminoglycoside | 0.5–64 | 16 | 8 | Susceptible |
| Kanamycin | Aminoglycoside | 4–512 | 64 | 64 | Susceptible |
| Tetracycline | Tetracycline | 1–128 | 32 | 16 | Susceptible |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kingcha, Y.; Pumpuang, L.; Adunphatcharaphon, S.; Chantarasakha, K.; Santiyanont, P.; Klomtun, M.; Janyaphisan, T.; Kongtong, K.; Phonsatta, N.; Panya, A.; et al. Potential Use of Lactiplantibacillus plantarum BCC 4352 as a Functional Starter Culture for Fermenting Thai Pork Sausage (Nham). Fermentation 2024, 10, 145. https://doi.org/10.3390/fermentation10030145
Kingcha Y, Pumpuang L, Adunphatcharaphon S, Chantarasakha K, Santiyanont P, Klomtun M, Janyaphisan T, Kongtong K, Phonsatta N, Panya A, et al. Potential Use of Lactiplantibacillus plantarum BCC 4352 as a Functional Starter Culture for Fermenting Thai Pork Sausage (Nham). Fermentation. 2024; 10(3):145. https://doi.org/10.3390/fermentation10030145
Chicago/Turabian StyleKingcha, Yutthana, Laphaslada Pumpuang, Saowalak Adunphatcharaphon, Kanittha Chantarasakha, Pannita Santiyanont, Manadsaree Klomtun, Thitiphorn Janyaphisan, Kittima Kongtong, Natthaporn Phonsatta, Atikorn Panya, and et al. 2024. "Potential Use of Lactiplantibacillus plantarum BCC 4352 as a Functional Starter Culture for Fermenting Thai Pork Sausage (Nham)" Fermentation 10, no. 3: 145. https://doi.org/10.3390/fermentation10030145
APA StyleKingcha, Y., Pumpuang, L., Adunphatcharaphon, S., Chantarasakha, K., Santiyanont, P., Klomtun, M., Janyaphisan, T., Kongtong, K., Phonsatta, N., Panya, A., Visessanguan, W., Petchkongkaew, A., & Woraprayote, W. (2024). Potential Use of Lactiplantibacillus plantarum BCC 4352 as a Functional Starter Culture for Fermenting Thai Pork Sausage (Nham). Fermentation, 10(3), 145. https://doi.org/10.3390/fermentation10030145