
Identifying a Role of Polysaccharides from Agaricus Blazei Murill in Combating Skin Photoaging: The Effect of Antioxidants on Fibroblast Behavior

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. The Preparation of AB-J and AB-K
2.3. Physical and Chemical Properties of AB-J and AB-K
2.4. Determination of Antioxidant Activity In Vitro
2.5. Cell Culture and Cytotoxicity Assay
2.6. Model and Sample Treatment
2.7. Determination of Intracellular ROS, GSH-Px and CAT
2.8. Determination of COL-I, ELN, and Matrix Metalloproteinase (MMPs)
2.9. Relative mRNA Expressions of Key Genes
2.10. Red Blood Cell (RBC) Hemolysis Assay
2.11. Eye Irritation Determination: HET-CAM
2.12. UV-Visible and FT-IR Spectrum Spectra
2.13. Monosaccharide Composition Analysis, GPC, SEM and AFM
2.14. Statistics Analysis
3. Results
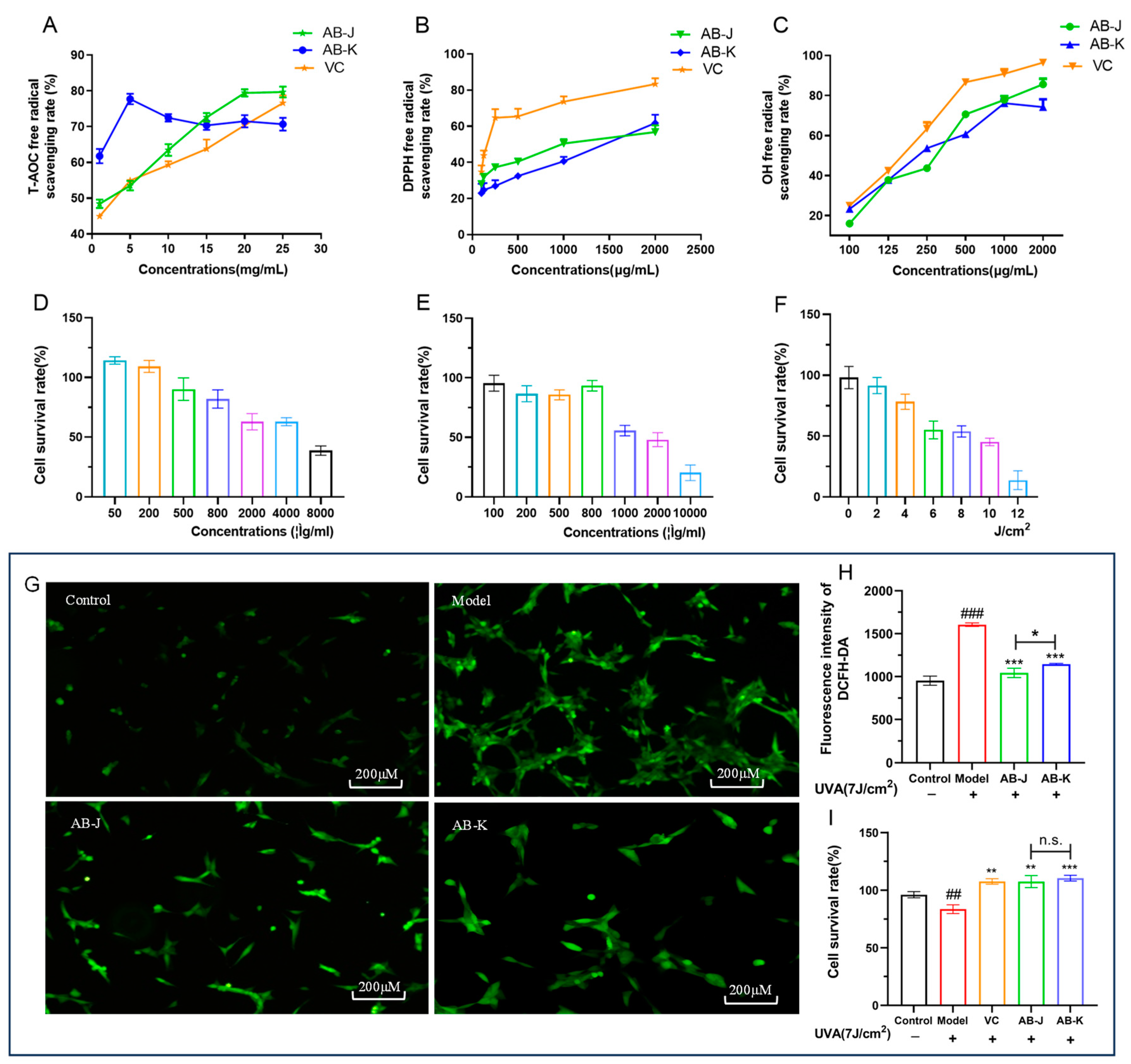
3.1. Content Index of AB-J and AB-K and Their Antioxidant Capacities In Vitro
3.2. AB-J and AB-K Can Accelerate Keap-1-Nrf2/ARE Signaling Pathway
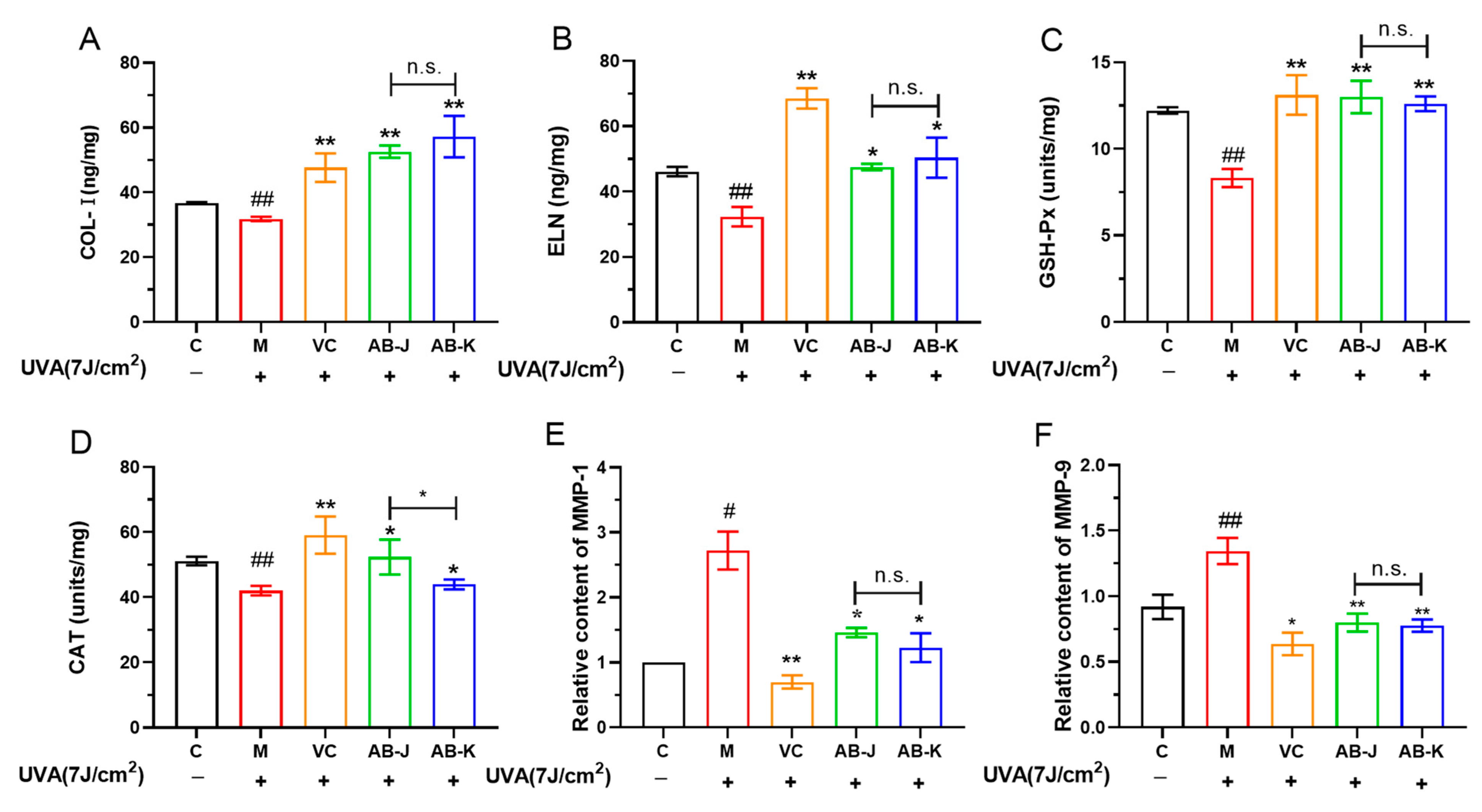
3.3. AB-J and AB-K Can Affect the Antioxidant Enzymes and Extracellular Matrix Levels
3.4. Safety Evaluation
3.5. AB-J Showed a Relatively Better Effect Than AB-K
3.6. Physical Properties of AB-J
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gonzaga, M.L.; Ricardo, N.M.; Heatley, F.; Soares, S. Isolation and characterization of polysaccharides from Agaricus blazei Murill. Carbohydr. Polym. 2005, 60, 43–49. [Google Scholar] [CrossRef]
- Stojkovi, D.; Reis, F.; Glamolija, J.; Ciric, A.; Barros, L.; Van Griensven, L.J.; Ferreira, I.C.; Sokovic, M. Cultivated strains of Agaricus bisporus and A. brasiliensis: Chemical characterization and evaluation of antioxidant and antimicrobial properties for the final healthy product – natural preservatives in yoghurt. Food Funct. 2014, 5, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.; Correa, V.; Gonalves, G.; Soares, A.; Bracht, A.; Peralta, R. Agaricus blazei bioactive compounds and their effects on human health: Benefits and controversies. Curr. Pharm. Des. 2017, 23, 2807–2834. [Google Scholar] [CrossRef]
- De Sá-Nakanishi, A.B.; Soares, A.A.; Natali, M.R.M.; Comar, J.F.; Peralta, R.M.; Bracht, A. Effects of the Continuous Administration of an Agaricus blazei Extract to Rats on Oxidative Parameters of the Brain and Liver during Aging. Molecules 2014, 19, 18590–18603. [Google Scholar] [CrossRef]
- Liu, P.; Yuan, J.; Jiang, Z.; Wang, Y.; Wen, B.; Li, G. A lower cadmium accumulating strain of Agaricus brasiliensis produced by 60 Co-γ-irradiation. LWT 2024, 114, 108370. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, D.; Fang, L.; Zhao, X.; Zhou, A.; Xie, J. A galactomannoglucan derived from Agaricus brasiliensis: Purification, characterization and macrophage activation via MAPK and I kappa B/NF kappa B pathways. Food Chem. 2018, 239, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Galhardi, L.C.; Rincao, V.; Soares, S.; Vieira, I.G.; Ricardo, N.M.; Nozawa, C.; Linhares, R.E. Antiherpetic activity of an Agaricus brasiliensis polysaccharide, its sulfated derivative and fractions. Int. J. Biol. Macromol. 2013, 52, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Lu, X.; Yuan, G.; Zhang, Q.; An, L. Effects of Agaricus blazei polypeptide on cell senescence by regulation of Keap1/Nrf2/ARE and TLR4/NF-κBp65 signaling pathways and its mechanism in D-gal-induced NIH/3T3 cells. J. Funct. Foods 2020, 72, 104037. [Google Scholar] [CrossRef]
- Gonzaga, M.L.; Campelo, M.; Saraiva, K.; Santos, A.Q.; Leal, L.K.; Ricardo, N.M.; Soares, S.; Riberiro, M.E. Chitosan and Agaricus brasiliensis Polysaccharides Films: A Preliminary Study. J. Braz. Chem. Soc. 2020, 31, 990–998. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Y. Structural analysis of an alkali-extractable and water-soluble polysaccharide (ABP-AW1) from the fruiting bodies of Agaricus blazei Murill. Carbohydr. Polym. 2011, 86, 429–432. [Google Scholar] [CrossRef]
- Lv, J.; Bai, F.; Wei, P. Effects of Agaricus blazei polysaccharide on oxygen free radicals and inflammatory related factors in diabetes rats. Guizhou Med. 2017, 41, 2. [Google Scholar] [CrossRef]
- Liu, W.; Yang, J.; Ren, J.; Zhang, D.; Ning, Z. Structural characterization and anti-inflammatory activity of ABD polysaccharides from Agaricus blazei. Mod. Food Technol. 2017, 33, 7. [Google Scholar] [CrossRef]
- Cheng, F.; Yan, X.; Zhang, M.; Chang, M.; Yun, S.; Meng, J.; Liu, J.; Feng, C. Regulation of RAW 264.7 cell-mediated immunity by polysaccharides from Agaricus blazei Murill via the MAPK signal transduction pathway. Food Funct. 2017, 8, 1475–1480. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Cheng, W.; Wang, Q.; Zhang, J.; Wang, C.; Li, M.; Zhao, D.; Wang, D.; An, Q. Exploring the Protective and Reparative Mechanisms of G. lucidum Polysaccharides Against H2O2-Induced Oxidative Stress in Human Skin Fibroblasts. Clin. Cosmet. Investig. Dermatol. 2021, 14, 1481–1496. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Gong, X.; Wei, D.; Liu, Y.; Zhao, M.; Yu, X.; Olatunji, O.; Jiao, X.; Zhen, O. Antioxidant Activity and Preliminary Structure Analysis of Polysaccharides from Cordyceps cicadas. Food Sci. 2016, 37, 19–24. [Google Scholar] [CrossRef]
- Cheng, W.; Yang, Y.; Zhang, J.; Di, F.; Li, L.; Wang, C.; Li, M.; Zhao, D.; Shi, X.; Huo, T.; et al. Protective mechanisms of intra- and extracellular polysaccharides from Schizophyllum commune on H2O2-induced oxidative damage of human skin fibroblasts. Food Agric. Immunol. 2023, 34, 2203407. [Google Scholar] [CrossRef]
- Wang, M. Recent developments in Hericium erinaceus polysaccharides: Extraction, purification, structural characteristics and biological activities. Crit. Rev. Food Sci. Nutr. 2019, 59, 96–115. [Google Scholar] [CrossRef] [PubMed]
- Florence, D.; Cedric, L.; Alix, V.; Olivier, T. UV, stress and aging. Derm. Endocrinol. 2014, 4, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Agar, N.; Halliday, G.; Barnetson, R.; Ananthaswamy, H.; Wheeler, M.; Jones, A. The basal layer in human squamous tumors harbors more UVA than UVB fingerprint mutations: A role for UVA in human skin carcinogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 4954–4959. [Google Scholar] [CrossRef]
- Li, Q.; Bai, D.; Qin, L.; Shao, M.; Zhang, S.; Yan, C.; Yu, G.; Hao, J. Protective effect of D-tetramannuronic acid tetrasodium salt on UVA-induced photo-aging in HaCaT cells. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 126, 110094. [Google Scholar] [CrossRef]
- Jaszewska, E.; Soin, M.; Filipek, A.; Naruszewicz, M. UVA-induced ROS generation inhibition by Oenothera paradoxa defatted seeds extract and subsequent cell death in human dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2013, 126, 42–46. [Google Scholar] [CrossRef]
- Lee, H.; Hong, Y.; Tran, Q.; Cho, H.; Kim, M.; Kim, C.; Kwon, S.; Park, S.; Park, J.; Park, J. A new role for the ginsenoside RG3 in antiaging via mitochondria function in ultraviolet-irradiated human dermal fibroblasts. J. Ginseng. Res. 2019, 43, 431–441. [Google Scholar] [CrossRef]
- Varani, J.; Warner, R.; Gharaee-Kermani, M.; Phan, S.; Kang, S.; Chung, J.; Wang, Z.; Datta, S.; Fisher, S.; Voorhees, J. Vitamin A Antagonizes Decreased Cell Growth and Elevated Collagen-Degrading Matrix Metalloproteinases and Stimulates Collagen Accumulation in Naturally Aged Human Skin1. J. Investig. Dermatol. 2000, 114, 480–486. [Google Scholar] [CrossRef]
- Kim, J.; Kim, D.; Kim, H.; Jang, A. Protection effect of donkey hide gelatin hydrolysates on UVB-induced photoaging of human skin fibroblasts. Process Biochem. 2018, 67, 118–126. [Google Scholar] [CrossRef]
- Lee, K.; Park, J.; Jung, E.; Ryu, J.; Kim, Y.; Youm, J.; Kang, S. A study of facial wrinkles improvement effect of veratric acid from cauliflower mushroom through photo-protective mechanisms against UVB irradiation. Arch. Dermatol. Res. 2016, 308, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.; Chang, C.; Gowrisankar, Y.; Chen, X.; Lin, H.; Yen, H.; Yang, H. Zerumbone Exhibits Antiphotoaging and Dermatoprotective Properties in Ultraviolet A-Irradiated Human Skin Fibroblast Cells via the Activation of Nrf2/ARE Defensive Pathway. Oxidative Med. Cell. Longev. 2019, 4098674. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Zheng, C.; Zheng, Q.; Chen, S.; Li, W.; Shang, Z.; Zhang, H. Salvianolic acid A attenuates early brain injury after subarachnoid hemorrhage in rats by regulating ERK/P38/Nrf2 signaling. Am. J. Transl. Res. 2017, 9, 5643–5652. [Google Scholar]
- Sevag, M.; Lackman, D.; Smolens, J. The isolation of components of Streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.; Zheng, Y.; He, Z.; Guan, P.; He, X.; Hui, L.; Dai, Y. Study of Schiff base formation between dialdehyde cellulose and proteins, and its application for the deproteinization of crude polysaccharide extracts. Ind. Crops Prod. 2018, 112, 532–540. [Google Scholar] [CrossRef]
- You, S.; Wang, C.; Zhang, J.; Li, M.; Zhao, D.; An, Q. An Preliminary purification and anti-aging activity evaluation of total flavonoids from seabuckthorn seed meal prepared by fermentation method. Dly. Chem. Ind. 2019, 49, 7. [Google Scholar]
- Mu, H.; Gao, H.; Chen, H.; Fang, X.; Ge, L. Determination of total flavanoid content in Asparagus officinalis Linn by spectrofluorimetry. J. Chin. Inst. Food Sci. Technol. 2010, 10, 201–205. [Google Scholar] [CrossRef]
- Cao, H.; Liu, C.; Li, S. Congo Red Method for Quantitative Detection of Yeast β- Research on the Methods of Dextran in the Food and Fermentation Industry. Food Ferment. Ind. 2022, 48, 261–266. [Google Scholar] [CrossRef]
- Tu, J.; Zhang, X.; Yu, L.; Liu, H.; Wu, F.; Liu, X.; Ma, X. Studies on rapid determination of beta-glucan content in grain products by flow-injecting Congo red spectrophotometry. Cereal Feed. Ind. 2009, 46–48. [Google Scholar]
- GB5009.91-2017; Food Safety National Standard—Determination of Potassium and Sodium in Food. Standardization Administration of China: Beijing, China, 2017.
- Liu, Z.; Jiao, Y.; Lu, H.; Shu, X.; Chen, Q. Chemical characterization, antioxidant properties and anticancer activity of exopolysaccharides from Floccularia luteovirens. Carbohydr. Polym. 2019, 229, 115432. [Google Scholar] [CrossRef]
- Min, W.; Fang, X.; Wu, T.; Fang, L.; Liu, C.; Wang, J. Characterization and antioxidant activity of an acidic exopolysaccharide from Lactobacillus plantarum JLAU103. J. Bioence Bioeng. 2018, 127, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Han, A.; Park, S.; Cho, C.; Rhee, Y.; Hong, H. Effect of enzyme-assisted extraction on the physicochemical properties and bioactive potential of lotus leaf polysaccharides. Int. J. Biol. Macromol. 2020, 153, 169–179. [Google Scholar] [CrossRef]
- Hseu, Y.; Lo, H.; Korivi, M.; Tsai, Y.; Tang, M.; Yang, H. Dermato-protective properties of ergothioneine through induction of Nrf2/ARE-mediated antioxidant genes in UVA-irradiated Human keratinocytes. Free Radic. Biol. Med. 2015, 86, 102–117. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Singleton, B.; Libretto, P.; Sibley, C.V.; Andrews, C. An in vitro haemolysis test as an alternative to the draize test for ocular irritation. Comp. Haematol. Int. 1994, 4, 49–54. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, X.; Zhang, W.; Cai, J.; Xue, J.; Yang, G.; Tan, X.; Xie, X.; Xiong, X.; Huang, J. Combined in vitro tests as an alternative to in vivo eye irritation tests. Altern. Lab. Anim. Atla 2010, 38, 303–314. [Google Scholar] [CrossRef]
- Chen, Z.; Yin, C.; Fan, X.; Ma, K.; Yao, F.; Zhou, R.; Shi, D.; Cheng, W.; Gao, H. Characterization of physicochemical and biological properties of Schizophyllum commune polysaccharide extracted with different methods. Int. J. Biol. Macromol. 2020, 156, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Safaryan, M.; Ganjloo, A.; Bimakr, M.; Zarringhalami, S. Optimization of Ultrasound-Assisted Extraction, Preliminary Characterization and In Vitro Antioxidant Activity of Polysaccharides from Green Pea Pods. Foods 2016, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; You, S.; Wang, D.; Zhao, D.; Zhang, J.; An, Q.; Li, M.; Wang, C. Fermented Dendrobium officinale polysaccharides protect UVA-induced photoaging of human skin fibroblasts. Food Sci. Nutr. 2022, 10, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Natural Nrf2 Modulators for Skin Protection. Antioxidants 2020, 9, 812. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Mockabee-macias, A.; Jiang, C.; Falzone, A.; Prieto-Farigua, N.; Stone, E.; Harris, I.; Denicola, G.; Mockabee-Macias, A.; Jiang, C.; et al. Non-canonical glutamate-cysteine ligase activity protects against ferroptosis. Cell Metab. 2020, 33, 174. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Jinlin, D.; Wang, F.; Yuan, Z.; Xue, J.; Lu, T.; Huang, W.; Liu, Y.; Zhang, Y. GSTM3 deficiency impedes DNA mismatch repair to promote gastric tumorigenesis via CAND1/NRF2-KEAP1 signaling. Cancer Lett. 2022, 538, 215692. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.; Jaiswal, A. Antioxidant-induced Phosphorylation of Tyrosine 486 Leads to Rapid Nuclear Export of Bach1 That Allows Nrf2 to Bind to the Antioxidant Response Element and Activate Defensive Gene Expression. J. Biol. Chem. 2010, 285, 153–162. [Google Scholar] [CrossRef]
- Xu, Y.; Cui, Y.; Wang, X.; Yue, F.; Shan, Y.; Liu, B.; Zhou, Y.; Yi, Y.; Lu, X. Purification, characterization and bioactivity of exopolysaccharides produced by Lactobacillus plantarum KX041. Int. J. Biol. Macromol. 2019, 128, 480–492. [Google Scholar] [CrossRef]
- Zhao, D.; Jiang, J.; Du, R.; Guo, S.; Ping, W.; Ling, H.; Ge, J. Purification and characterization of an exopolysaccharide from Leuconostoc lactis L2. Int. J. Biol. Macromol. Struct. Funct. Interact. 2019, 139, 1224–1231. [Google Scholar] [CrossRef]
- Asgher, M.; Urooj, Y.; Qamar, S.; Khalid, N. Improved exopolysaccharide production from Bacillus licheniformis MS3: Optimization and structural/functional characterization. Int. J. Biol. Macromol. 2020, 151, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Bhandary, T.; Kurian, C.; Muthu, M.; Anand, A.; Anand, T.; Paari, K. Exopolysaccharides Derived from Probiotic Bacteria and their Health Benefits. J. Pure Appl. Microbiol. 2023, 17, 35–40. [Google Scholar] [CrossRef]
- Shao, L.; Wu, Z.; Zhang, H.; Chen, W.; Guo, B. Observation of Exopolysaccharide S2 from Lactobacillus rhamnosus KF5 Using Atomic Force Microscopy. Food Sci. 2015, 36, 43–47. [Google Scholar] [CrossRef]
- Hyde, K.; Bahkali, A.; Moslem, M. Fungi-an unusual source for cosmetics. Fungal Divers. 2010, 43, 1–9. [Google Scholar] [CrossRef]
- Jiang, C.; Ge, J.; He, B.; Zeng, B. Glycosphingolipids in Filamentous Fungi: Biological Roles and Potential Applications in Cosmetics and Health Foods. Front. Microbiol. 2021, 12, 690211. [Google Scholar] [CrossRef]
- Taofiq, O.; Rodrigues, F.; Barros, L.; Peralta, R.; Barreiro, M.; Feeewira, I.C.; Oliveira, M.B. Agaricus blazei Murrill from Brazil: An ingredient for nutraceutical and cosmeceutical applications. Food Funct. 2019, 10, 565–572. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, R.; Xie, Y.; Wang, H.; Ge, M. The Protective Effects of Polysaccharides from Agaricus blazei Murill Against Cadmium-Induced Oxidant Stress and Inflammatory Damage in Chicken Livers. Biol. Trace Elem. Res. 2016, 178, 117–126. [Google Scholar] [CrossRef]
- Al-Dbass, A.; Al-Daihan, S.; Bhat, R. Agaricus blazei Murill as an efficient hepatoprotective and antioxidant agent against CCl4 -induced liver injury in rats. Saudi J. Biol. Sci. 2012, 19, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, O.M.; Vellosa, J.C.; Fernandes, A.; Buffa-Fiho, W.; Hakime-Silva, R.; Furlan, M.; Brunetti, I. Antioxidant activity of Agaricus blazei. Fitoterapia 2007, 78, 263–264. [Google Scholar] [CrossRef]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef]
- Bhatti, J.; Bhatti, G.; Reddy, P. Mitochondrial dysfunction and oxidative stress in metabolic disorders — A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta (BBA) - Mol. Basis Dis. 2016, 1863, 1066–1077. [Google Scholar] [CrossRef]
- Xu, H. The role of oxidative stress in the pathogenesis of diabetes related periodontitis. Int. J. Stomatol. 2011, 38, 4. [Google Scholar] [CrossRef]
- Fang, J.; Sun, Q.; Wang, Z.; Song, Z.; Wang, C.; Li, M.; Wang, D. Enhancement of Human Epidermal Cell Defense against UVB Damage by Fermentation of Passiflora edulis Sims Peel with Saccharomyces cerevisiae. Nutrients 2023, 15, 501. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.; Shao, Y.; Wu, Y.; Hsu, W.; Cheng, K.; Yu, C.; Chou, C.; Hsieh, C. Physicochemical properties and protective effects on UVA-induced photoaging in Hs68 cells of Pleurotus ostreatus polysaccharides by fractional precipitation. Int. J. Biol. Macromol. 2023, 228, 537–547. [Google Scholar] [CrossRef]
- Chen, B.; Huang, H.; Tsai, K.; Wu, J.; Chang, Y.; Chang, M.; Lu, C.; Yang, S.; Huang, H. Protective Effect of a Water-Soluble Carotenoid-Rich Extract of Cordyceps militaris against Light-Evoked Functional Vision Deterioration in Mice. Nutrients 2022, 14, 1675. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Stress-sensing mechanisms and the physiological roles of the Keap1–Nrf2 system during cellular stress. J. Biol. Chem. 2017, 292, 16817–16824. [Google Scholar] [CrossRef] [PubMed]
- Canning, P.; Sorrell, F.; Bullock, A. Structural basis of Keap1 interactions with Nrf2. Free Radic. Biol. Med. 2015, 88, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, W.; Feng, X.; Yang, F.; Qin, H.; Wu, S.; Hou, D.; Chen, J. Nrf2-ARE Signaling Acts as Master Pathway for the Cellular Antioxidant Activity of Fisetin. Molecules 2019, 24, 708. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, J.; Sanapalli, B.K.; Bano, M.; Singh, S.; Gulati, M.; Karri, V.V. Nanostructured Lipid Carriers of Pioglitazone Loaded Collagen/Chitosan Composite Scaffold for Diabetic Wound Healing. Adv. Wound Care 2019, 8, 499–513. [Google Scholar] [CrossRef]
- Aluri, H.; Kublin, C.; Thotakura, S.; Armaos, H.; Samizadeh, M.; Hawley, D.; Thomas, W.; Leavis, P.; Makarenkova, H.; Zoukhri, D. Role of Matrix Metalloproteinases 2 and 9 in Lacrimal Gland Disease in Animal Models of Sjögren's Syndrome. Investig. Opthalmology Vis. Sci. 2015, 56, 5218–5228. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Kim, S.; Chung, H.; Pae, H. Chapter Two - Reactive Oxygen Species in the Activation of MAP Kinases. Methods Enzymol. 2013, 528, 27–48. [Google Scholar] [PubMed]
- Liu, Q.; Xiao, X.; Hu, L.; Jie, H.; Wang, Y.; Ye, W.; Li, M.; Liu, Z.; Anhuienoside, C. Ameliorates Collagen-Induced Arthritis through Inhibition of MAPK and NF-κB Signaling Pathways. Front. Pharmacol. 2017, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Guo, W.; Zhang, W.; Xu, J.; Qian, M.; Bai, W.; Zhang, Y.; Rao, P.; Ni, L.; Lv, X. Grifola frondosa polysaccharides ameliorate lipid metabolic disorders and gut microbiota dysbiosis in high-fat diet fed rats. Food Funct. 2019, 10, 2560–2572. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.; Tsai, Y.; Li, I.; Chen, M.; Huang, Y. Hp-s1 Ganglioside Suppresses Proinflammatory Responses by Inhibiting MyD88-Dependent NF-κB and JNK/p38 MAPK Pathways in Lipopolysaccharide-Stimulated Microglial Cells. Mar. Drugs 2020, 18. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.H.; Yen, G.C. Differential expressions of antioxidant status in aging rats: The role of transcriptional factor Nrf2 and MAPK signaling pathway. Biogerontroly 2007, 8, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, Y.; Sun, J.; Ma, J.; Cheng, C. Protective role of quercetin against lead-induced inflammatory response in rat kidney through the ROS-mediated MAPKs and NF-κB pathway. Biochim. Biophys. Acta 2012, 1820, 1693–1703. [Google Scholar] [CrossRef]
- Wyatt, L.; Luz, A.; Cao, X.; Maurer, L.; Blawas, A.; Aballay, A.; Pan, W.K.; Meyer, J. Effects of methyl and inorganic mercury exposure on genome homeostasis and mitochondrial function in Caenorhabditis elegans. DNA Repair. 2017, 52, 31–48. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Primer Pair Sequence (5′→3′) |
|---|---|---|
| β-actin | F R | TGGCACCCAGCACAATGAA CTAAGTCATAGTCCGCCTAGAAGCA |
| Nrf2 | F R | CAACTCAGCACCTTGTATC TTCTTAGTATCTGGCTTCTT |
| Keap-1 | F R | GGAGGCGGAGCCCGA GATGCCCTCAATGGACACCA |
| P38 | F R | TTAACAGGATGCCAAGCCATGA GGCACCAATAAATACATTCGCAAAG |
| Jnk1 | F R | CTGTGTGGAATCAAGCACCTTCA CTGGCCAGACCGAAGTCAAGA |
| ERK | F R | CCGAGTGACGAGCCCAT CTCCCTTGCTAGAGCTCACT |
| NQO1 | F R | CAGCCAATCAGCGTTCGGTA CTTCATGGCGTAGTTGAATGATGTC |
| HO-1 | F R | CAAGCGCTATGTTCAGCGAC GCTTGAACTTGGTGGCACTG |
| MMP-1 | F R | GCA TATCGATGCTGCTCTTTC GATAACCTGGATCCATAGATCGTT |
| MMP-9 | F R | GTACTCGACCACTTCTTGTCG AGAAGCCCCACTTCTTGTCG |
| Stimulus Score | Classification |
|---|---|
| IS < 1 | No irritation |
| 1 ≤ IS < 5 | Mid irritation |
| 5 ≤ IS < 9 | Moderate irritation |
| IS ≥ 9 | Strong irritation |
| Sample | AB-J | AB-K | |
|---|---|---|---|
| Content (%) | |||
| fattiness | (0.04 ± 3.2)% | (0.5 ± 4.1)% | |
| protein | (23.7 ± 3.1)% | (8.69 ± 2.1)% | |
| ash | (2.8 ± 1.6)% | (0.36 ± 1.8)% | |
| moisture | (22.2 ± 2.3)% | (17.2 ± 2.1)% | |
| carbohydrate | (51.3 ± 2.3)% | (73.2 ± 4.1)% | |
| Sample | Parallel | Bleeding | Blood Vessel | Angiolysis | ES/Point | IS |
|---|---|---|---|---|---|---|
| AB-J | 1 | 0 | 0 | 0 | 0 | 0.08 |
| 2 | 0 | 0 | 0 | |||
| 3 | 0 | 0 | 0 | |||
| 4 | 0 | 0 | 0 | |||
| 5 | 0 | 0 | 0 | |||
| 6 | 0 | 0 | 0 | |||
| AB-K | 1 | 0 | 0 | 0 | 0 | 0.08 |
| 2 | 0 | 0 | 0 | |||
| 3 | 0 | 0 | 0 | |||
| 4 | 0 | 0 | 0 | |||
| 5 | 0 | 0 | 0 | |||
| 6 | 0 | 0 | 0 | |||
| Negative Control (0.9% NaCl) | 1 | 0 | 0 | 0 | 0 | 0.07 |
| 2 | 0 | 0 | 0 | |||
| 3 | 0 | 0 | 0 | |||
| 4 | 0 | 0 | 0 | |||
| 5 | 0 | 0 | 0 | |||
| 6 | 0 | 0 | 0 | |||
| Positive Control (0.1 mol/L NaOH) | 1 | 2 | 2 | 1 | 12 | 17.42 |
| 2 | 2 | 1 | 1 | |||
| 3 | 2 | 2 | 1 | |||
| 4 | 2 | 1 | 1 | |||
| 5 | 2 | 1 | 1 | |||
| 6 | 2 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di, F.; Cheng, W.; Li, L.; Pu, C.; Sun, R.; Zhang, J.; Wang, C.; Li, M. Identifying a Role of Polysaccharides from Agaricus Blazei Murill in Combating Skin Photoaging: The Effect of Antioxidants on Fibroblast Behavior. Fermentation 2024, 10, 292. https://doi.org/10.3390/fermentation10060292
Di F, Cheng W, Li L, Pu C, Sun R, Zhang J, Wang C, Li M. Identifying a Role of Polysaccharides from Agaricus Blazei Murill in Combating Skin Photoaging: The Effect of Antioxidants on Fibroblast Behavior. Fermentation. 2024; 10(6):292. https://doi.org/10.3390/fermentation10060292
Chicago/Turabian StyleDi, Feiqian, Wenjing Cheng, Luyao Li, Chunhong Pu, Ruiliang Sun, Jiachan Zhang, Changtao Wang, and Meng Li. 2024. "Identifying a Role of Polysaccharides from Agaricus Blazei Murill in Combating Skin Photoaging: The Effect of Antioxidants on Fibroblast Behavior" Fermentation 10, no. 6: 292. https://doi.org/10.3390/fermentation10060292