

Optimization of Biodegradation of Common Bean Biomass for Fermentation Using Trichoderma asperellum WNZ-21 and Artificial Neural Networks
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of RCBBs
2.2. Cultivation Medium
2.3. Modeling of AA Production
2.3.1. Screening the Medium Components Using the DSD
2.3.2. The Architecture of the ANN
2.3.3. Software and Statistical Procedure
2.4. Chemical Tests
2.4.1. AA Detection
2.4.2. Phytochemical Analysis
2.4.3. High-Performance Liquid Chromatography (HPLC) Analysis
2.4.4. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.4.5. Fourier Transform Infrared (FT-IR) Spectroscopy
2.5. High-Resolution Transmission Electron Microscopy (HR-TEM)
2.6. Biological Tests
2.6.1. Antioxidant Activity
2.6.2. Antibacterial Activity
2.7. Anticancer Activity
2.7.1. MTT Assay
2.7.2. Gene Expression in Cancer Cells
2.7.3. Data Analysis
3. Results and Discussion
3.1. DSD Paradigm for Screening the Medium Components
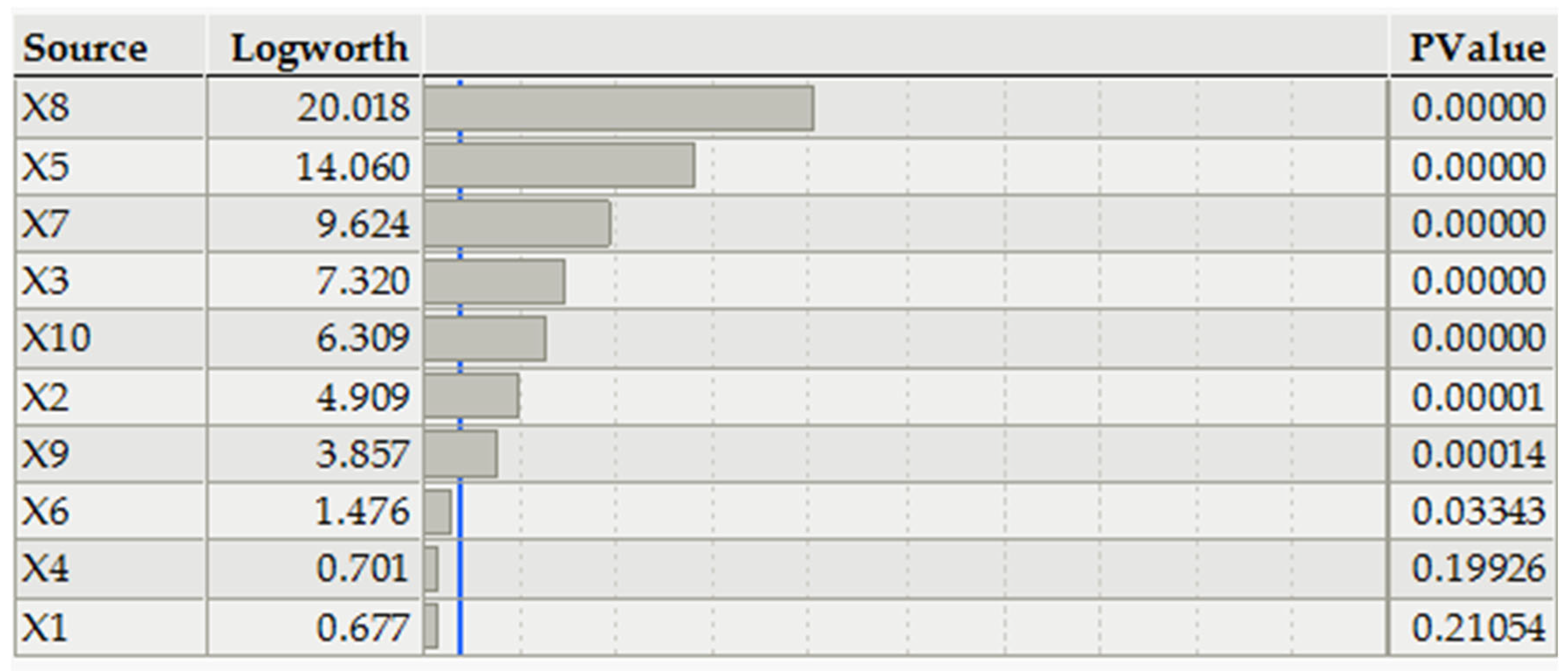
3.1.1. Coefficients and ANOVA
3.1.2. Adequacy of DSD
3.1.3. Residual Analysis
3.2. Machine Learning for Modeling AA Production
3.2.1. The Architecture of the ANN
3.2.2. Training and Validation Processes
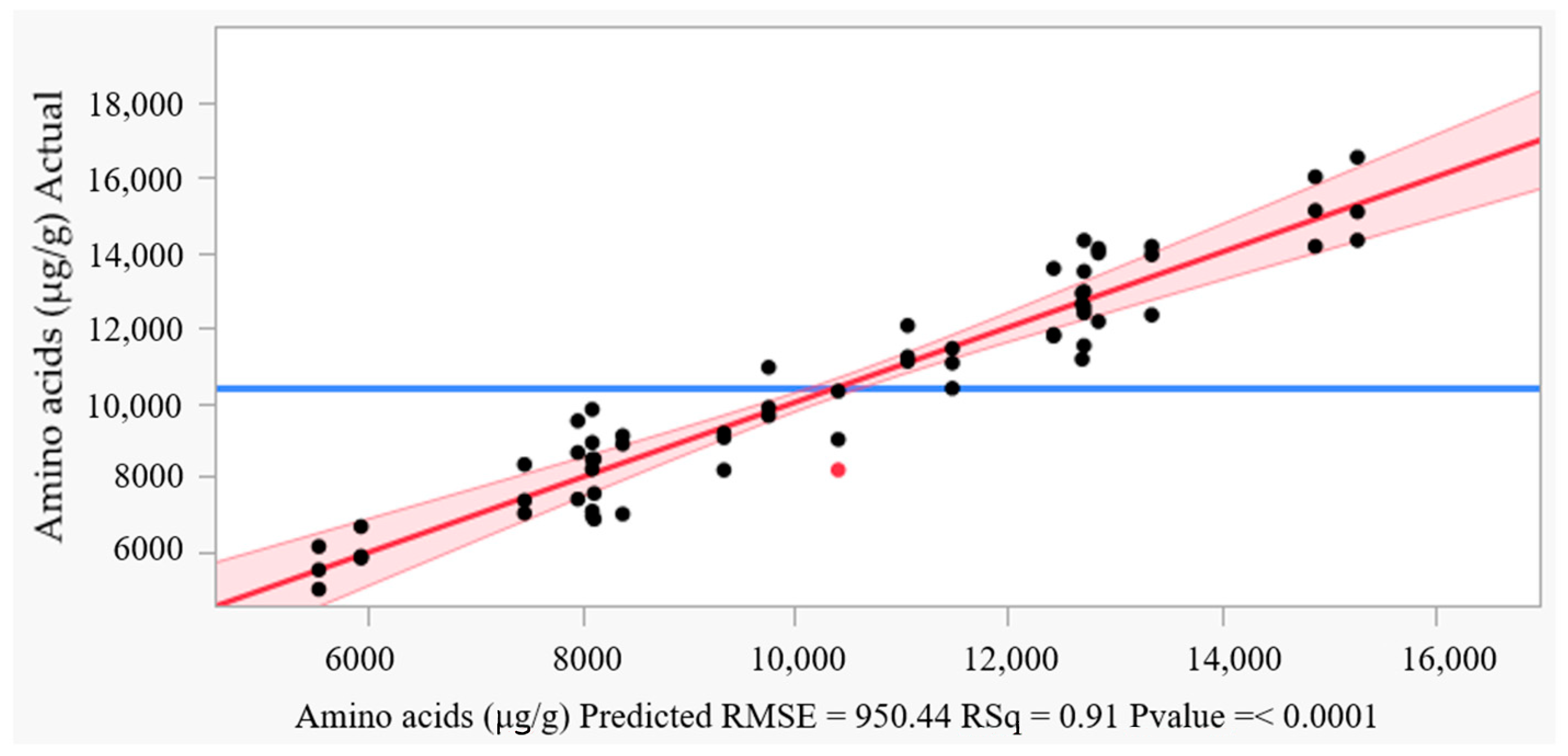
3.2.3. Prediction and Residual Analysis
3.2.4. The Experimental Testing of the ANN Model
3.3. Biochemical Composition of Fungal Filtrate
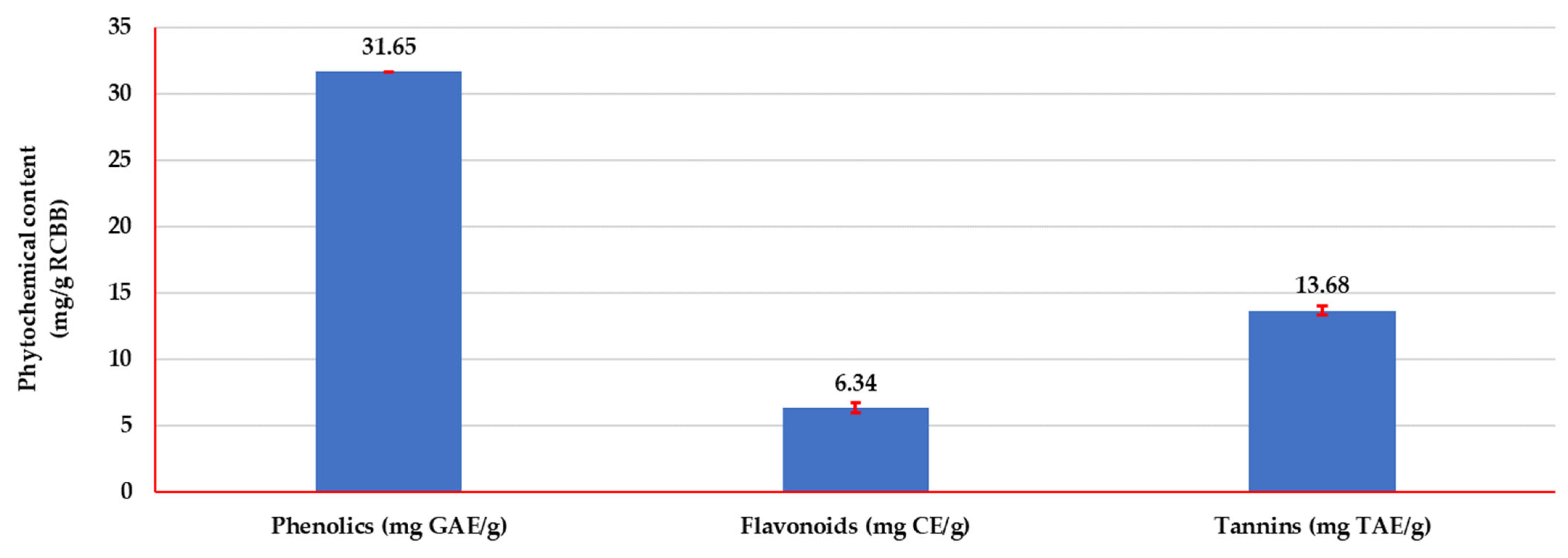
3.3.1. Phytochemical Analysis
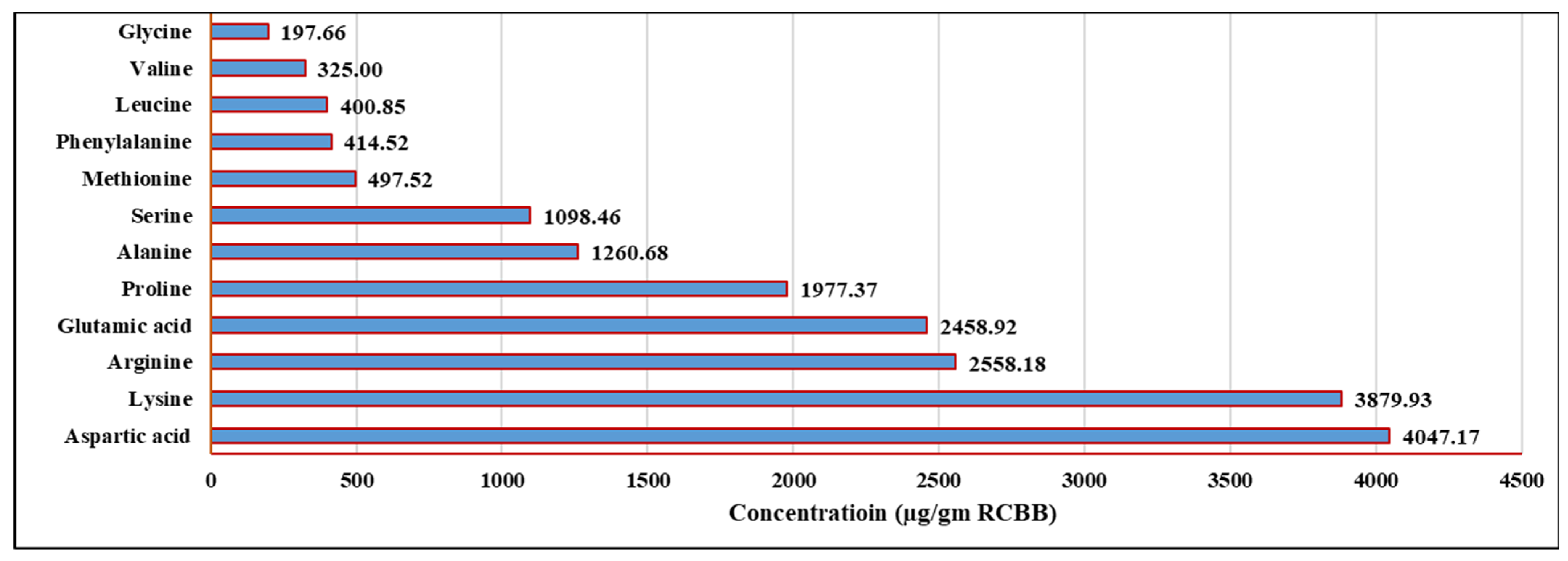
3.3.2. HPLC Profile of AAs
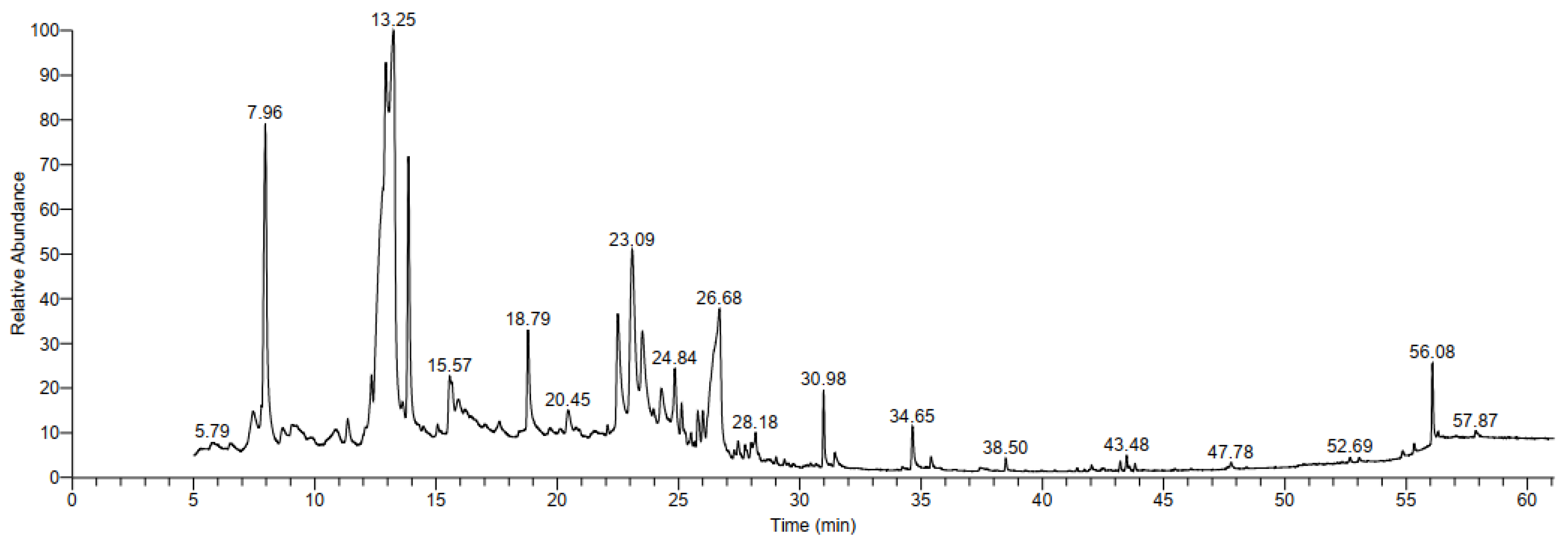
3.3.3. GC-MS Analysis
3.3.4. FT-IR Spectral Analysis
3.4. Structural Investigation
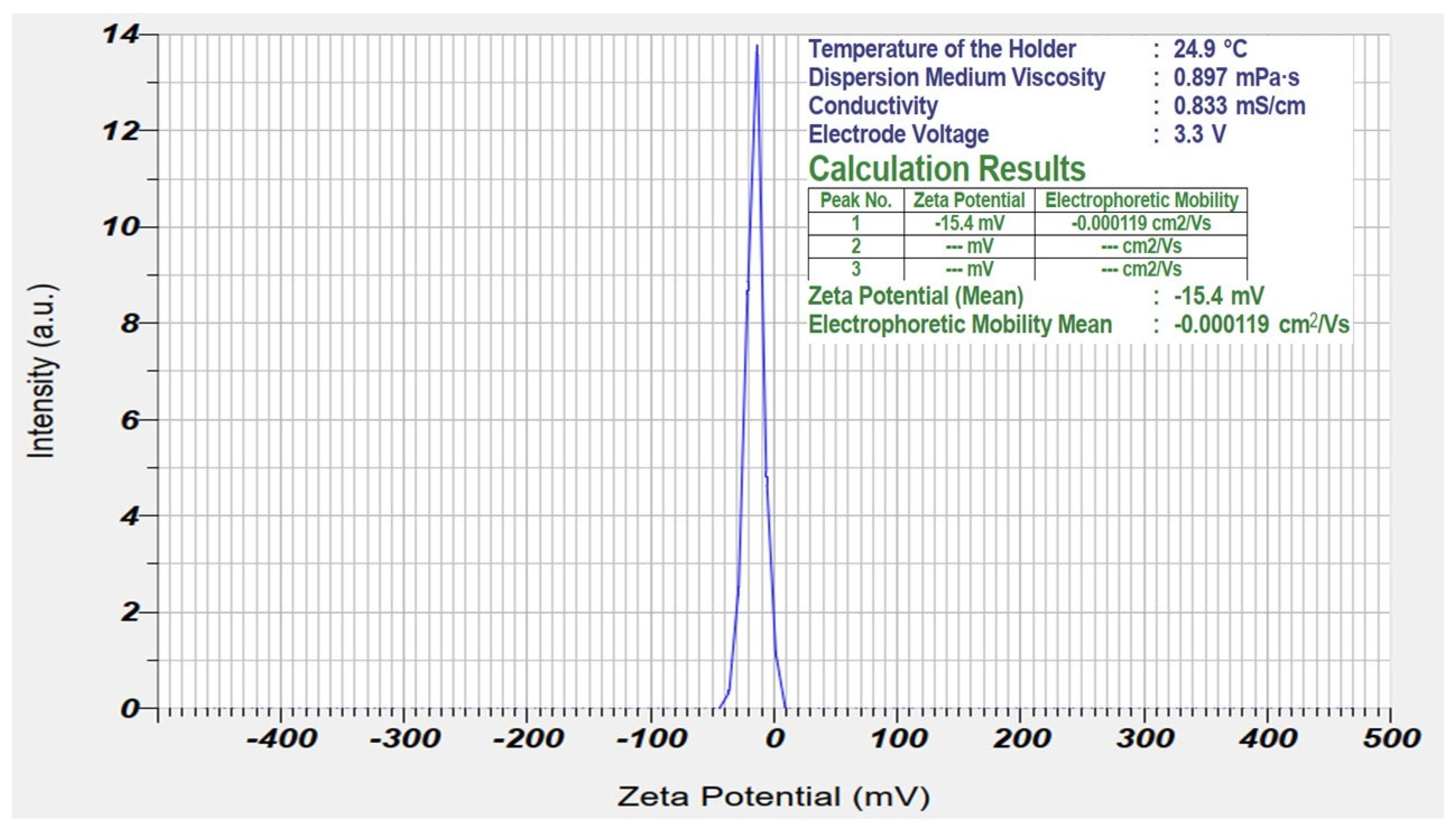
3.4.1. Zeta Potential Analysis
3.4.2. HR-TEM Investigation
3.5. Biological Activity of Fungal Filtrate
3.5.1. Antioxidant Activity
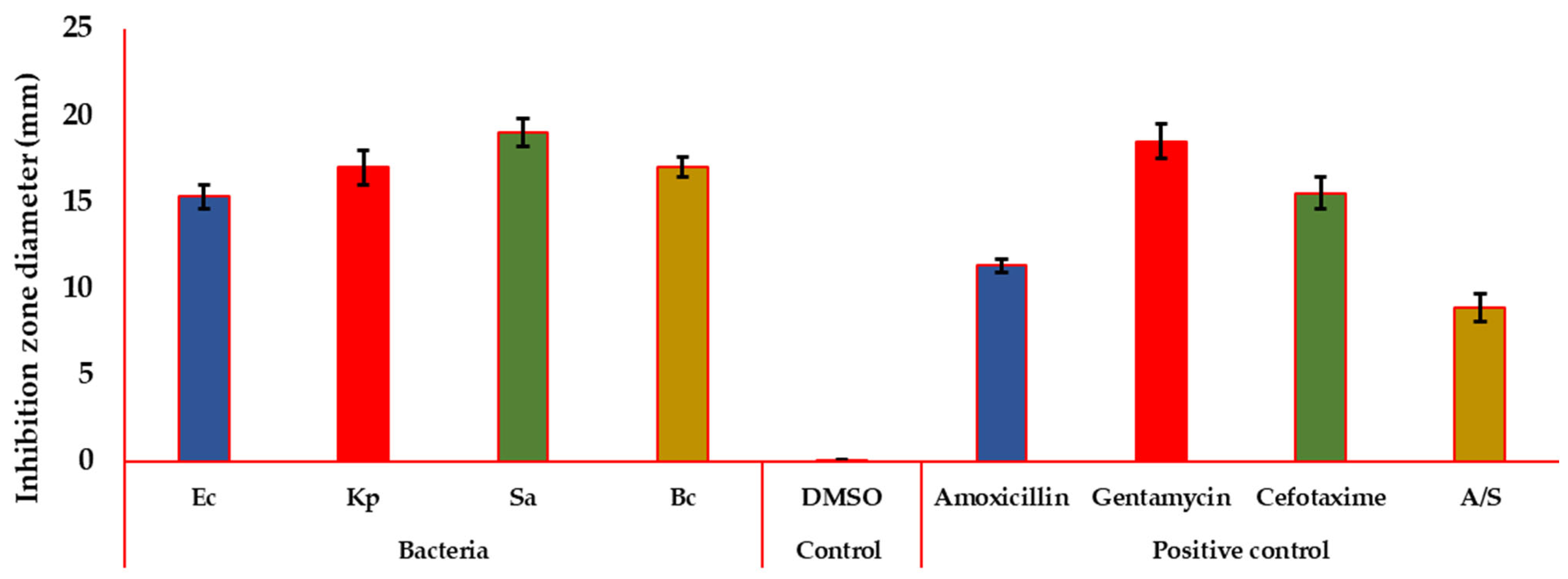
3.5.2. Antibacterial Activity
3.6. Anticancer Activity
3.6.1. Cell Viability Assay
3.6.2. Apoptotic Modulation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gimenes, N.C.; Silveira, E.; Tambourgi, E.B. An overview of proteases: Production, downstream processes and industrial applications. Sep. Purif. Rev. 2021, 50, 223–243. [Google Scholar] [CrossRef]
- Verma, N.; Kumar, V.; Bansal, M. Valorization of waste biomass in fermentative production of cellulases: A review. Waste Biomass Valorization 2021, 12, 613–640. [Google Scholar] [CrossRef]
- Moussa, Z.; Darwish, D.B.; Alrdahe, S.S.; Saber, W.I.A. Innovative Artificial-Intelligence-Based Approach for the Biodegradation of Feather Keratin by Bacillus paramycoides, and Cytotoxicity of the Resulting Amino Acids. Front. Microbiol. 2021, 12, 731262. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M. The Chemistry of Protein Oxidation in Food. Angew. Chem. Int. Ed. Engl. 2019, 58, 16742–16763. [Google Scholar] [CrossRef] [PubMed]
- Vandana, U.K.; Rajkumari, J.; Singha, L.P.; Satish, L.; Alavilli, H.; Sudheer, P.D.; Chauhan, S.; Ratnala, R.; Satturu, V.; Mazumder, P.B. The endophytic microbiome as a hotspot of synergistic interactions, with prospects of plant growth promotion. Biology 2021, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.K.; Dey, Y.N.; Madhavan, Y.; Maity, A. Fungal Endophytes: A Storehouse of Bioactive Compounds. Mini Rev. Med. Chem. 2023, 23, 978–991. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Meshram, V.; Gupta, M.; Goyal, S.; Qureshi, K.A.; Jaremko, M.; Shukla, K.K. Fungal Endophytes: Microfactories of Novel Bioactive Compounds with Therapeutic Interventions; A Comprehensive Review on the Biotechnological Developments in the Field of Fungal Endophytic Biology over the Last Decade. Biomolecules 2023, 13, 1038. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.H.; Attia, M.S.; Kandil, E.K.; Fawzi, M.M.; Abdelrahman, A.S.; Khader, M.S.; Khodaira, M.A.; Emam, A.E.; Goma, M.A.; Abdelaziz, A.M. Bioactive compounds and biomedical applications of endophytic fungi: A recent review. Microb. Cell Factories 2023, 22, 107. [Google Scholar] [CrossRef] [PubMed]
- Omomowo, I.O.; Amao, J.A.; Abubakar, A.; Ogundola, A.F.; Ezediuno, L.O.; Bamigboye, C.O. A review on the trends of endophytic fungi bioactivities. Sci. Afr. 2023, 20, e01594. [Google Scholar] [CrossRef]
- Elsayed, M.S.; Eldadamony, N.M.; Alrdahe, S.S.T.; Saber, W.I.A. Definitive Screening Design and Artificial Neural Network for Modeling a Rapid Biodegradation of Date Palm Fronds by a New Trichoderma sp. PWN6 into Citric Acid. Molecules 2021, 26, 5048. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.; Moussa, Z.; Alrdahe, S.S.; Alharbi, M.M.; Ghoniem, A.A.; El-Khateeb, A.Y.; Saber, W.I.A. Optimization of Heavy Metals Biosorption via Artificial Neural Network: A Case Study of Cobalt (II) Sorption by Pseudomonas alcaliphila NEWG-2. Front. Microbiol. 2022, 13, 893603. [Google Scholar] [CrossRef] [PubMed]
- Fakhry, H.; Ghoniem, A.A.; Al-Otibi, F.O.; Helmy, Y.A.; El Hersh, M.S.; Elattar, K.M.; Saber, W.I.A.; Elsayed, A. A Comparative Study of Cr(VI) Sorption by Aureobasidium pullulans AKW Biomass and Its Extracellular Melanin: Complementary Modeling with Equilibrium Isotherms, Kinetic Studies, and Decision Tree Modeling. Polymers 2023, 15, 3754. [Google Scholar] [CrossRef] [PubMed]
- El-Metwally, M.M.; Abdel-Fattah, G.M.; Al-Otibi, F.O.; Khatieb, D.; Helmy, Y.A.; Mohammed, Y.M.M.; Saber, W.I.A. Application of artificial neural networks for enhancing Aspergillus flavipes lipase synthesis for green biodiesel production. Heliyon 2023, 9, e20063. [Google Scholar] [CrossRef] [PubMed]
- Saber, W.I.A.; Al-Askar, A.A.; Ghoneem, K.M. Exclusive Biosynthesis of Pullulan Using Taguchi’s Approach and Decision Tree Learning Algorithm by a Novel Endophytic Aureobasidium pullulans Strain. Polymers 2023, 15, 1419. [Google Scholar] [CrossRef] [PubMed]
- Tunga, R.; Banerjee, R.; Bhattacharyya, B. Optimizing some factors affecting protease production under solid state fermentation. Bioprocess Eng. 1998, 19, 187–190. [Google Scholar] [CrossRef]
- Cupp-Enyard, C. Sigma’s Non-specific Protease Activity Assay—Casein as a Substrate. J. Vis. Exp. 2008, e899. [Google Scholar] [CrossRef]
- Aberoumand, A. Nutritional evaluation of edible Portulaca oleracia as plant food. Food Anal. Methods 2009, 2, 204–207. [Google Scholar] [CrossRef]
- Burlingame, B. Wild nutrition. J. Food Compos. Anal. 2000, 2, 99–100. [Google Scholar] [CrossRef]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Jajić, I.; Krstović, S.; Glamočić, D.; Jakšić, S.; Abramović, B. Validation of an HPLC method for the determination of amino acids in feed. J. Serbian Chem. Soc. 2013, 78, 839–850. [Google Scholar] [CrossRef]
- Kitts, D.D.; Wijewickreme, A.N.; Hu, C. Antioxidant properties of a North American ginseng extract. Mol. Cell. Biochem. 2000, 203, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Parejo, I.; Codina, C.; Petrakis, C.; Kefalas, P. Evaluation of scavenging activity assessed by Co(II)/EDTA-induced luminol chemiluminescence and DPPH·(2,2-diphenyl-1-picrylhydrazyl) free radical assay. J. Pharmacol. Toxicol. Methods 2000, 44, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Boyanova, L.; Gergova, G.; Nikolov, R.; Derejian, S.; Lazarova, E.; Katsarov, N.; Mitov, I.; Krastev, Z. Activity of Bulgarian propolis against 94 Helicobacter pylori strains in vitro by agar-well diffusion, agar dilution and disc diffusion methods. J. Med. Microbiol. 2005, 54, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Owuama, C.I. Determination of minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) using a novel dilution tube method. Afr. J. Microbiol. Res. 2017, 11, 977–980. [Google Scholar]
- Thabrew, M.I.; Hughes, R.D.; McFarlane, I.G. Screening of hepatoprotective plant components using a HepG2 cell cytotoxicity assay. J. Pharm. Pharmacol. 1997, 49, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.A.; Shawkey, A.E.; Rabeh, M.A.; Abdellatif, A.O. Expression of P53, BAX, and BCL-2 in human malignant melanoma and squamous cell carcinoma cells after tea tree oil treatment in vitro. Cytotechnology 2019, 71, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.P.; Morris, J.C.; Montori, V.M. Thyroid cancer: Zealous imaging has increased detection and treatment of low risk tumours. BMJ 2013, 347, f4706. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.-A.; Saber, W.I.; Zweil, A.M.; Bashir, S.I. An innovative green synthesis approach of chitosan nanoparticles and their inhibitory activity against phytopathogenic Botrytis cinerea on strawberry leaves. Sci. Rep. 2022, 12, 3515. [Google Scholar] [CrossRef] [PubMed]
- Lakhdari, W.; Benyahia, I.; Bouhenna, M.M.; Bendif, H.; Khelafi, H.; Bachir, H.; Ladjal, A.; Hammi, H.; Mouhoubi, D.; Khelil, H.; et al. Exploration and Evaluation of Secondary Metabolites from Trichoderma harzianum: GC-MS Analysis, Phytochemical Profiling, Antifungal and Antioxidant Activity Assessment. Molecules 2023, 28, 5025. [Google Scholar] [CrossRef]
- Omomowo, I.; Fadiji, A.; Omomowo, O. Antifungal evaluation and phytochemical profile of Trichoderma harzianum and Glomus versiforme secondary metabolites on cowpea pathogens. Asian J. Microbiol. Biotechnol. Environ. Sci. 2020, 22, 265–272. [Google Scholar]
- Sumilat, D.A.; Lintang, R.A.J.; Undap, S.L.; Adam, A.A.; Tallei, T.E. Phytochemical, antioxidant, and antimicrobial analysis of Trichoderma asperellum isolated from ascidian Eudistoma sp. J. Appl. Pharm. Sci. 2022, 12, 090–095. [Google Scholar] [CrossRef]
- Ciurko, D.P.; Łaba, W.; Piegza, M.; Juszczyk, P.; Choińska-Pulit, A.; Sobolczyk-Bednarek, J. Enzymatic bioconversion of feather waste with keratinases of PCM 2849. Pol. J. Chem. Technol. 2019, 21, 53–59. [Google Scholar] [CrossRef]
- Moussa, Z.; Alanazi, Y.F.; Khateb, A.M.; Eldadamony, N.M.; Ismail, M.M.; Saber, W.I.A.; Darwish, D.B.E. Domiciliation of Trichoderma asperellum Suppresses Globiosporangium ultimum and Promotes Pea Growth, Ultrastructure, and Metabolic Features. Microorganisms 2023, 11, 198. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, S.; Parkin, K.L. Amino acids, peptides, and proteins. In Fennema’s Food Chemistry; CRC Press: Boca Raton, FL, USA, 2017; pp. 235–356. [Google Scholar]
- Murdock, J.N.; Wetzel, D.L. FT-IR microspectroscopy enhances biological and ecological analysis of algae. Appl. Spectrosc. Rev. 2009, 44, 335–361. [Google Scholar] [CrossRef]
- Adochitei, A.; Drochioiu, G. Rapid characterization of peptide secondary structure by FT-IR spectroscopy. Rev. Roum. Chim. 2011, 56, 783–791. [Google Scholar]
- Honary, S.; Zahir, F. Effect of zeta potential on the properties of nano-drug delivery systems-a review (Part 1). Trop. J. Pharm. Res. 2013, 12, 255–264. [Google Scholar]
- Tao, B.; Ren, C.; Li, H.; Liu, B.; Jia, X.; Dong, X.; Zhang, S.; Chang, H. Thio-/LISICON and LGPS-Type Solid Electrolytes for All-Solid-State Lithium-Ion Batteries. Adv. Funct. Mater. 2022, 32, 2203551. [Google Scholar] [CrossRef]
- Loget, G.; Kuhn, A. Electric field-induced chemical locomotion of conducting objects. Nat. Commun. 2011, 2, 535. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Zhong, L.; Sun, Y.; Sun, N.; Zhang, L.; Liu, W.; Qu, Y.; Zhong, Y. Enhancement of Cellulase Production in Trichoderma reesei via Disruption of Multiple Protease Genes Identified by Comparative Secretomics. Front. Microbiol. 2019, 10, 2784. [Google Scholar] [CrossRef]
- Cherif, M.; Benhamou, N. Cytochemical aspects of chitin breakdown during the parasitic action of a Trichoderma sp. on Fusarium oxysporum f. sp. radicis-lycopersici. Phytopathology 1990, 80, 1406–1414. [Google Scholar] [CrossRef]
- Cardoso, M.M.; Barradas, M.J.; Kroner, K.H.; Crespo, J.G. Amino acid solubilization in cationic reversed micelles: Factors affecting amino acid and water transfer. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean. Technol. 1999, 74, 801–811. [Google Scholar] [CrossRef]
- Mohamed, S.A.; Saleh, R.M.; Kabli, S.A.; Al-Garni, S.M. Influence of solid state fermentation by Trichoderma spp. on solubility, phenolic content, antioxidant, and antimicrobial activities of commercial turmeric. Biosci. Biotechnol. Biochem. 2016, 80, 920–928. [Google Scholar] [CrossRef]
- Konappa, N.; Udayashankar, A.C.; Dhamodaran, N.; Krishnamurthy, S.; Jagannath, S.; Uzma, F.; Pradeep, C.K.; De Britto, S.; Chowdappa, S.; Jogaiah, S. Ameliorated antibacterial and antioxidant properties by Trichoderma harzianum mediated green synthesis of silver nanoparticles. Biomolecules 2021, 11, 535. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Heo, Y.M.; Jang, S.; Lee, H.; Kwon, S.-L.; Park, M.S.; Lim, Y.W.; Kim, J.-J. Diversity of Trichoderma spp. in marine environments and their biological potential for sustainable industrial applications. Sustainability 2020, 12, 4327. [Google Scholar] [CrossRef]
- Nagarajan, S.; Nagarajan, R.; Kumar, J.; Salemme, A.; Togna, A.R.; Saso, L.; Bruno, F. Antioxidant Activity of Synthetic Polymers of Phenolic Compounds. Polymers 2020, 12, 1646. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Aboul-Enein, B.H.; Duchnik, E.; Marchlewicz, M. Antioxidative properties of phenolic compounds and their effect on oxidative stress induced by severe physical exercise. J. Physiol. Sci. 2022, 72, 19. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Chen, G.; Liu, H. Antioxidative Categorization of Twenty Amino Acids Based on Experimental Evaluation. Molecules 2017, 22, 2066. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Jang, H.-J.; Cho, W.-Y.; Yeon, S.-J.; Lee, C.-H. In Vitro antioxidant actions of sulfur-containing amino acids. Arab. J. Chem. 2020, 13, 1678–1684. [Google Scholar] [CrossRef]
- Matemu, A.; Nakamura, S.; Katayama, S. Health Benefits of Antioxidative Peptides Derived from Legume Proteins with a High Amino Acid Score. Antioxidants 2021, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.S.; Paiva-Martins, F. Amino Acids, Amino Acid Derivatives and Peptides as Antioxidants. In Lipid Oxidation in Food and Biological Systems: A Physical Chemistry Perspective; Springer: Berlin/Heidelberg, Germany, 2022; pp. 381–404. [Google Scholar]
- Kavaz, D.; Faraj, R.E. Investigation of composition, antioxidant, antimicrobial and cytotoxic characteristics from Juniperus sabina and Ferula communis extracts. Sci. Rep. 2023, 13, 7193. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zeng, W.; Li, Z.; Peng, Y. Mannose regulates water balance, leaf senescence, and genes related to stress tolerance in white clover under osmotic stress. Biol. Plant. 2020, 64, 406–416. [Google Scholar] [CrossRef]
- Tian, D.; Qiao, Y.; Peng, Q.; Zhang, Y.; Gong, Y.; Shi, L.; Xiong, X.; He, M.; Xu, X.; Shi, B. A Poly-D-Mannose Synthesized by a One-Pot Method Exhibits Anti-Biofilm, Antioxidant, and Anti-Inflammatory Properties In Vitro. Antioxidants 2023, 12, 1579. [Google Scholar] [CrossRef]
- Fernandes, P.A.R.; Coimbra, M.A. The antioxidant activity of polysaccharides: A structure-function relationship overview. Carbohydr. Polym. 2023, 314, 120965. [Google Scholar] [CrossRef] [PubMed]
- Mu, S.; Yang, W.; Huang, G. Antioxidant activities and mechanisms of polysaccharides. Chem. Biol. Drug Des. 2021, 97, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Ecevit, K.; Barros, A.A.; Silva, J.M.; Reis, R.L. Preventing microbial infections with natural phenolic compounds. Future Pharmacol. 2022, 2, 460–498. [Google Scholar] [CrossRef]
- Lobiuc, A.; Paval, N.E.; Mangalagiu, I.I.; Gheorghita, R.; Teliban, G.C.; Amariucai-Mantu, D.; Stoleru, V. Future Antimicrobials: Natural and Functionalized Phenolics. Molecules 2023, 28, 1114. [Google Scholar] [CrossRef] [PubMed]
- Idrees, M.; Mohammad, A.R.; Karodia, N.; Rahman, A. Multimodal Role of Amino Acids in Microbial Control and Drug Development. Antibiotics 2020, 9, 330. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.G.; Skwarecki, A.S.; Milewska, M.J. Amino Acid Based Antimicrobial Agents—Synthesis and Properties. ChemMedChem 2021, 16, 3513–3544. [Google Scholar] [CrossRef] [PubMed]
- Elshaer, A.; Hanson, P.; Worthington, T.; Lambert, P.; Mohammed, A.R. Preparation and characterization of amino acids-based trimethoprim salts. Pharmaceutics 2012, 4, 179–196. [Google Scholar] [CrossRef] [PubMed]
- Patyal, M.; Kaur, K.; Gupta, N.; Kaur, R.; Malik, A.K. Optical and Antimicrobial Activity of Nanostructured Mn (II) and Cu (II) Macrocyclic Complexes Derived from Aspartic Acid. Prot. Met. Phys. Chem. Surf. 2023, 59, 169–178. [Google Scholar] [CrossRef]
- Wu, Y.; He, Q.; Che, X.; Liu, F.; Lu, J.; Kong, X. Effect of number of lysine motifs on the bactericidal and hemolytic activity of short cationic antimicrobial peptides. Biochem. Biophys. Res. Commun. 2023, 648, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Rakhmawatie, M.D.; Marfu’ati, N.; Barsaliputri, B.; FikrIyah, A.Z.; Ethica, S.N. Antibacterial activity and GC-MS profile of secondary metabolites of Bacillus subtilis subsp. subtilis HSFI-9 associated with Holothuria scabra. Biodiversitas J. Biol. Divers. 2023, 24, 2843–2849. [Google Scholar] [CrossRef]
- Hamad, G.M.; Abd El-Baky, N.; Sharaf, M.M.; Amara, A.A. Volatile Compounds, Fatty Acids Constituents, and Antimicrobial Activity of Cultured Spirulina (Arthrospira fusiformis) Isolated from Lake Mariout in Egypt. Sci. World J. 2023, 2023, 9919814. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Chen, T.; Chen, X. A review of the antibacterial activity and mechanisms of plant polysaccharides. Trends Food Sci. Technol. 2022, 123, 264–280. [Google Scholar] [CrossRef]
- Bhatia, S.; Miller, N.J.; Lu, H.; Longino, N.V.; Ibrani, D.; Shinohara, M.M.; Byrd, D.R.; Parvathaneni, U.; Kulikauskas, R.; Ter Meulen, J.; et al. Intratumoral G100, a TLR4 Agonist, Induces Antitumor Immune Responses and Tumor Regression in Patients with Merkel Cell Carcinoma. Clin. Cancer Res. 2019, 25, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Bakrim, S.; El Omari, N.; El Hachlafi, N.; Bakri, Y.; Lee, L.-H.; Bouyahya, A. Dietary Phenolic Compounds as Anticancer Natural Drugs: Recent Update on Molecular Mechanisms and Clinical Trials. Foods 2022, 11, 3323. [Google Scholar] [CrossRef]
- Endo, H.; Inoue, I.; Masunaka, K.; Tanaka, M.; Yano, M. Curcumin induces apoptosis in lung cancer cells by 14-3-3 protein-mediated activation of Bad. Biosci. Biotechnol. Biochem. 2020, 84, 2440–2447. [Google Scholar] [CrossRef]
- Li, J.; Fan, Y.; Zhang, Y.; Liu, Y.; Yu, Y.; Ma, M. Resveratrol Induces Autophagy and Apoptosis in Non-Small-Cell Lung Cancer Cells by Activating the NGFR-AMPK-mTOR Pathway. Nutrients 2022, 14, 2413. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Liu, Z.; Wang, Y.; Zhang, Q.; Li, J.; Zhong, P.; Xie, Z.; Ji, A.; Li, Y. Epigallocatechin-3-gallate alleviates high-fat diet-induced nonalcoholic fatty liver disease via inhibition of apoptosis and promotion of autophagy through the ROS/MAPK signaling pathway. Oxid. Med. Cell. Longev. 2021, 2021, 5599997. [Google Scholar] [CrossRef] [PubMed]
- Sair, A.T.; Liu, R.H. Molecular regulation of phenolic compounds on IGF-1 signaling cascade in breast cancer. Food Funct. 2022, 13, 3170–3184. [Google Scholar] [CrossRef] [PubMed]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino acids in cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Pranzini, E.; Pardella, E.; Paoli, P.; Fendt, S.M.; Taddei, M.L. Metabolic Reprogramming in Anticancer Drug Resistance: A Focus on Amino Acids. Trends Cancer 2021, 7, 682–699. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.; Haq, E.A.; Dwiranti, A.; Prasedya, E.S.; Kitamoto, Y. Bifunctional folic-conjugated aspartic-modified Fe3O4 nanocarriers for efficient targeted anticancer drug delivery. RSC Adv. 2022, 12, 4961–4971. [Google Scholar] [CrossRef] [PubMed]
- Aslani, R.; Namazi, H. Simple fabrication of multifunctional hyperbranched copolymer based on l-lysine and citric acid for co-delivery of anticancer drugs to breast cancer cells. React. Funct. Polym. 2022, 170, 105101. [Google Scholar] [CrossRef]
- Han, N.-R.; Park, H.-J.; Ko, S.-G.; Moon, P.-D. Maltol has anti-cancer effects via modulating PD-L1 signaling pathway in B16F10 cells. Front. Pharmacol. 2023, 14, 1255586. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, C.; Bao, N. Molecular mechanism of palmitic acid and its derivatives in tumor progression. Front. Oncol. 2023, 13, 1224125. [Google Scholar] [CrossRef] [PubMed]
- Ju, H.; Yu, C.; Zhang, X.-D.; Liu, W.; Wu, Y.-C.; Gong, P.-X.; Li, H.-H.; Liu, Y.; Li, H.-J. Recent trends in anti-cancer activities of terrestrial plants-based polysaccharides: A review. Carbohydr. Polym. Technol. Appl. 2023, 6, 100341. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Level | ||||
|---|---|---|---|---|---|
| Name | Symbol | Unit | Low (−1) | Center (0) | High (+1) |
| NaH2PO4 | X1 | mg/g RCBB | 10.80 | 12.80 | 14.80 |
| KH2PO4 | X2 | 2.00 | 3.00 | 4.00 | |
| NaCl | X3 | 0.30 | 0.50 | 0.70 | |
| NH4Cl | X4 | 0.50 | 1.00 | 1.50 | |
| MgSO4·7H2O | X5 | 0.30 | 0.50 | 0.70 | |
| CaCl2·2H2O | X6 | 0.005 | 0.010 | 0.015 | |
| pH | X7 | 5.50 | 6.00 | 6.50 | |
| Time | X8 | Day | 7.0 | 9.0 | 11.0 |
| Inoculation | X9 | Spore/g RCBB | 1 × 106 | 2 × 106 | 3 × 106 |
| Temperature | X10 | °C | 25 | 30 | 35 |
| Run | Block | The Coded Level of the Independent Variable | Amino Acids (µg/g) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | DSD | ANN | ||||||||||||||
| X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | Predicted | Error | Predicted | Error | |||
| 1 * | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 14,151.58 | 14,872.12 | −720.54 | 15,113.26 | −241.14 |
| 2 | 1 | 0 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 5856.41 | 5938.34 | −81.93 | 6549.77 | −611.43 |
| 3 | 1 | 1 | 0 | −1 | −1 | 1 | −1 | 1 | 1 | 1 | −1 | 14,313.57 | 15,266.91 | −953.34 | 15,368.87 | −101.96 |
| 4 | 1 | −1 | 0 | 1 | 1 | −1 | 1 | −1 | −1 | −1 | 1 | 5027.50 | 5543.55 | −516.05 | 5588.06 | −44.51 |
| 5 * | 1 | 1 | −1 | 0 | −1 | 1 | 1 | −1 | 1 | −1 | 1 | 14,310.57 | 12,707.83 | 1602.74 | 13,498.79 | −790.96 |
| 6 | 1 | −1 | 1 | 0 | 1 | −1 | −1 | 1 | −1 | 1 | −1 | 8222.18 | 8102.63 | 119.55 | 8039.90 | 62.73 |
| 7 | 1 | 1 | −1 | −1 | 0 | −1 | 1 | 1 | −1 | 1 | 1 | 8932.11 | 8103.29 | 828.82 | 7250.51 | 852.78 |
| 8 | 1 | −1 | 1 | 1 | 0 | 1 | −1 | −1 | 1 | −1 | −1 | 11,509.85 | 12,707.16 | −1197.31 | 11,727.83 | 979.33 |
| 9 * | 1 | 1 | 1 | 1 | −1 | 0 | −1 | −1 | −1 | 1 | 1 | 7051.29 | 7469.19 | −417.90 | 7743.02 | −273.83 |
| 10 * | 1 | −1 | −1 | −1 | 1 | 0 | 1 | 1 | 1 | −1 | −1 | 14,151.58 | 13,341.27 | 810.31 | 13,442.04 | −100.77 |
| 11 * | 1 | 1 | −1 | 1 | 1 | −1 | 0 | −1 | 1 | 1 | −1 | 12,609.74 | 12,689.00 | −79.26 | 12,096.10 | 592.90 |
| 12 * | 1 | −1 | 1 | −1 | −1 | 1 | 0 | 1 | −1 | −1 | 1 | 7577.24 | 8121.46 | −544.22 | 8016.38 | 105.08 |
| 13 * | 1 | 1 | 1 | −1 | 1 | −1 | −1 | 0 | 1 | −1 | 1 | 9510.05 | 7968.64 | 1541.41 | 8555.07 | −586.43 |
| 14 | 1 | −1 | −1 | 1 | −1 | 1 | 1 | 0 | −1 | 1 | −1 | 12,151.78 | 12,841.82 | −690.04 | 13,376.95 | −535.13 |
| 15 * | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | 0 | −1 | −1 | 12,043.80 | 11,056.00 | 987.80 | 11,216.67 | −160.67 |
| 16 | 1 | −1 | −1 | −1 | 1 | 1 | −1 | −1 | 0 | 1 | 1 | 9864.01 | 9754.46 | 109.55 | 9718.07 | 36.39 |
| 17 | 1 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | 0 | −1 | 7030.30 | 8387.93 | −1357.63 | 8094.15 | 293.78 |
| 18 | 1 | −1 | −1 | 1 | −1 | −1 | −1 | 1 | 1 | 0 | 1 | 11,764.82 | 12,422.53 | −657.71 | 12,809.30 | −386.77 |
| 19 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | 1 | −1 | −1 | 0 | 11,050.89 | 11,473.96 | −423.07 | 10,541.29 | 932.67 |
| 20 | 1 | −1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | 0 | 9062.09 | 9336.50 | −274.41 | 8506.97 | 829.53 |
| 21 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10,299.97 | 10,405.23 | −105.26 | 9512.85 | 892.38 |
| 22 | 2 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 15,103.49 | 14,872.12 | 231.37 | 15,113.26 | −241.14 |
| 23 | 2 | 0 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 6696.33 | 5938.34 | 757.99 | 6549.77 | −611.43 |
| 24 | 2 | 1 | 0 | −1 | −1 | 1 | −1 | 1 | 1 | 1 | −1 | 16,520.35 | 15,266.91 | 1253.44 | 15,368.87 | −101.96 |
| 25 | 2 | −1 | 0 | 1 | 1 | −1 | 1 | −1 | −1 | −1 | 1 | 6164.38 | 5543.55 | 620.83 | 5588.06 | −44.51 |
| 26 | 2 | 1 | −1 | 0 | −1 | 1 | 1 | −1 | 1 | −1 | 1 | 13,485.65 | 12,707.83 | 777.82 | 13,498.79 | −790.96 |
| 27 | 2 | −1 | 1 | 0 | 1 | −1 | −1 | 1 | −1 | 1 | −1 | 8490.15 | 8102.63 | 387.52 | 8039.90 | 62.73 |
| 28 * | 2 | 1 | −1 | −1 | 0 | −1 | 1 | 1 | −1 | 1 | 1 | 7109.29 | 8103.29 | −994.00 | 7250.51 | 852.78 |
| 29 * | 2 | −1 | 1 | 1 | 0 | 1 | −1 | −1 | 1 | −1 | −1 | 12,380.76 | 12,707.16 | −326.40 | 11,727.83 | 979.33 |
| 30 | 2 | 1 | 1 | 1 | −1 | 0 | −1 | −1 | −1 | 1 | 1 | 7388.26 | 7469.19 | −80.93 | 7743.02 | −273.83 |
| 31 | 2 | −1 | −1 | −1 | 1 | 0 | 1 | 1 | 1 | −1 | −1 | 13,928.61 | 13,341.27 | 587.34 | 13,442.04 | −100.77 |
| 32 | 2 | 1 | −1 | 1 | 1 | −1 | 0 | −1 | 1 | 1 | −1 | 11,148.89 | 12,689.00 | −1540.11 | 12,096.10 | 592.90 |
| 33 | 2 | −1 | 1 | −1 | −1 | 1 | 0 | 1 | −1 | −1 | 1 | 6891.31 | 8121.46 | −1230.15 | 8016.38 | 105.08 |
| 34 | 2 | 1 | 1 | −1 | 1 | −1 | −1 | 0 | 1 | −1 | 1 | 8666.13 | 7968.64 | 697.49 | 8555.07 | −586.43 |
| 35 | 2 | −1 | −1 | 1 | −1 | 1 | 1 | 0 | −1 | 1 | −1 | 13975.6 | 12,841.82 | 1133.78 | 13,376.95 | −535.13 |
| 36 | 2 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | 0 | −1 | −1 | 11,091.89 | 11,056.00 | 35.89 | 11,216.67 | −160.67 |
| 37 | 2 | −1 | −1 | −1 | 1 | 1 | −1 | −1 | 0 | 1 | 1 | 10,932.91 | 9754.46 | 1178.45 | 9718.07 | 36.39 |
| 38 * | 2 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | 0 | −1 | 8892.11 | 8387.93 | 504.18 | 8094.15 | 293.78 |
| 39 | 2 | −1 | −1 | 1 | −1 | −1 | −1 | 1 | 1 | 0 | 1 | 11,809.82 | 12,422.53 | −612.71 | 12,809.30 | −386.77 |
| 40 * | 2 | 1 | −1 | 1 | 1 | 1 | −1 | 1 | −1 | −1 | 0 | 11,435.86 | 11,473.96 | −38.10 | 10,541.29 | 932.67 |
| 41 | 2 | −1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | 0 | 8200.18 | 9336.50 | −1136.32 | 8506.97 | 829.53 |
| 42 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9014.10 | 10,405.23 | −1391.13 | 9512.85 | 892.38 |
| 43 * | 3 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 16,001.40 | 14,872.12 | 1129.28 | 15,113.26 | −241.14 |
| 44 * | 3 | 0 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 5902.41 | 5938.34 | −35.93 | 6549.77 | −611.43 |
| 45 * | 3 | 1 | 0 | −1 | −1 | 1 | −1 | 1 | 1 | 1 | −1 | 15,071.49 | 15,266.91 | −195.42 | 15,368.87 | −101.96 |
| 46 | 3 | −1 | 0 | 1 | 1 | −1 | 1 | −1 | −1 | −1 | 1 | 5543.45 | 5543.55 | −0.10 | 5588.06 | −44.51 |
| 47 | 3 | 1 | −1 | 0 | −1 | 1 | 1 | −1 | 1 | −1 | 1 | 12,544.75 | 12,707.83 | −163.08 | 13,498.79 | −790.96 |
| 48 | 3 | −1 | 1 | 0 | 1 | −1 | −1 | 1 | −1 | 1 | −1 | 9813.02 | 8102.63 | 1710.39 | 8039.90 | 62.73 |
| 49 * | 3 | 1 | −1 | −1 | 0 | −1 | 1 | 1 | −1 | 1 | 1 | 6990.30 | 8103.29 | −1112.99 | 7250.51 | 852.78 |
| 50 | 3 | −1 | 1 | 1 | 0 | 1 | −1 | −1 | 1 | −1 | −1 | 12,952.70 | 12,707.16 | 245.54 | 11,727.83 | 979.33 |
| 51 | 3 | 1 | 1 | 1 | −1 | 0 | −1 | −1 | −1 | 1 | 1 | 8349.17 | 7469.19 | 879.98 | 7743.02 | −273.83 |
| 52 * | 3 | −1 | −1 | −1 | 1 | 0 | 1 | 1 | 1 | −1 | −1 | 12,324.77 | 13,341.27 | −1016.50 | 13,442.04 | −100.77 |
| 53 | 3 | 1 | −1 | 1 | 1 | −1 | 0 | −1 | 1 | 1 | −1 | 12,905.71 | 12,689.00 | 216.71 | 12,096.10 | 592.90 |
| 54 * | 3 | −1 | 1 | −1 | −1 | 1 | 0 | 1 | −1 | −1 | 1 | 8493.15 | 8121.46 | 371.69 | 8016.38 | 105.08 |
| 55 | 3 | 1 | 1 | −1 | 1 | −1 | −1 | 0 | 1 | −1 | 1 | 7426.26 | 7968.64 | −542.38 | 8555.07 | −586.43 |
| 56 * | 3 | −1 | −1 | 1 | −1 | 1 | 1 | 0 | −1 | 1 | −1 | 14,094.59 | 12,841.82 | 1252.77 | 13,376.95 | −535.13 |
| 57 | 3 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | 0 | −1 | −1 | 11,211.88 | 11,056 | 155.88 | 11,216.67 | −160.67 |
| 58 * | 3 | −1 | −1 | −1 | 1 | 1 | −1 | −1 | 0 | 1 | 1 | 9646.04 | 9754.46 | −108.42 | 9718.07 | 36.39 |
| 59 * | 3 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | 0 | −1 | 9112.09 | 8387.93 | 724.16 | 8094.15 | 293.78 |
| 60 | 3 | −1 | −1 | 1 | −1 | −1 | −1 | 1 | 1 | 0 | 1 | 13,563.64 | 12,422.53 | 1141.11 | 12,809.30 | −386.77 |
| 61 | 3 | 1 | −1 | 1 | 1 | 1 | −1 | 1 | −1 | −1 | 0 | 10,374.96 | 11,473.96 | −1099.00 | 10,541.29 | 932.67 |
| 62 | 3 | −1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | 0 | 9187.08 | 9336.50 | −149.42 | 8506.97 | 829.53 |
| 63 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8203.18 | 10,405.23 | −2202.05 | 9512.85 | 892.38 |
| Source | Coefficient | Freedom Degree | Sum of Squares | Mean of Squares | F Ratio | Prob > F * | VIF |
|---|---|---|---|---|---|---|---|
| Model | 10,405 | 10 | 477,587,902 | 47,758,790 | 52.87 | 0.0000 | - |
| X1 | 164 | 1 | 1,451,766 | 1,451,766 | 1.61 | 0.2110 | 1.00 |
| X2 | −625 | 1 | 21,097,031 | 21,097,031 | 23.35 | 0.0000 | 1.00 |
| X3 | 825 | 1 | 36,786,002 | 36,786,002 | 40.72 | 0.0000 | 1.00 |
| X4 | −168 | 1 | 1,527,152 | 1,527,152 | 1.69 | 0.1990 | 1.00 |
| X5 | 1387 | 1 | 103,943,196 | 103,943,196 | 115.07 | 0.0000 | 1.00 |
| X6 | 283 | 1 | 4,312,066 | 4,312,066 | 4.77 | 0.0330 | 1.00 |
| X7 | 1013 | 1 | 55,365,589 | 55,365,589 | 61.29 | 0.0000 | 1.00 |
| X8 | 1963 | 1 | 208,032,461 | 208,032,461 | 230.29 | 0.0000 | 1.00 |
| X9 | 532 | 1 | 15,288,751 | 15,288,751 | 16.92 | 0.0000 | 1.00 |
| X10 | −743 | 1 | 29,783,888 | 29,783,888 | 32.97 | 0.0000 | 1.00 |
| Error | - | 52 | 46,973,828 | 903,343 | - | - | - |
| Lack-of-Fit | - | 10 | 15,095,995 | 1,509,599 | 1.99 | 0.0590 | - |
| Pure error | - | 42 | 31,877,833 | 758,996 | - | - | - |
| Total | - | 62 | 524,561,730 | - | - | - | - |
| The goodness-of-fit statistics | |||||||
| Coefficient of determination (R2) | 0.9105 | ||||||
| Adjusted-R2 | 0.8932 | ||||||
| Predicted-R2 | 0.8720 | ||||||
| Measure | Training | Validation |
|---|---|---|
| R2 | 0.9138 | 0.9433 |
| Root average square error | 816.35 | 720.38 |
| Mean absolute deviation | 673.67 | 661.48 |
| −Log-likelihood | 341.20 | 167.97 |
| Sum frequency | 42 | 21 |
| Test Point | Investigated Parameter | Amino Acids (µg/g RCBB) | Desirability | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| X2 | X3 | X5 | X6 | X7 | X8 | X9 | X10 | Predicted | Experimental | ||
| Optimal | 2.42 | 0.7 | 0.7 | 0.015 | 6.3 | 11 | 3,000,000 | 25 | 18,582.52 | 18,298.14 ± 97.08 | 0.9977 |
| One | 1.50 | 0.5 | 0.6 | 0.005 | 5.0 | 7 | 1,500,000 | 30 | 9583.93 | 9805.02 ± 103.96 | |
| Two | 2.00 | 0.4 | 0.3 | 0.010 | 5.5 | 8 | 2,500,000 | 35 | 5873.05 | 5905.71 ± 59.70 | |
| Three | 1.00 | 0.6 | 0.4 | 0.005 | 7.0 | 10 | 1,000,000 | 30 | 10,216.68 | 10,999.12 ± 150.00 | |
| Four | 3.00 | 0.5 | 0.5 | 0.010 | 6.0 | 9 | 2,000,000 | 25 | 9760.88 | 9700.25 ± 11.03 | |
| Peak | RT | Name | Formula | Molecular Weight | Area Sum % |
|---|---|---|---|---|---|
| 1 | 7.45 | Palmitic acid | C16H32O2 | 256 | 1.88 |
| 2 | 796 | Citraconic anhydride | C5H4O3 | 112 | 11.99 |
| 3 | 12.33 | Undecanal | C11H22O | 170 | 1.24 |
| 4 | 12.68 | 2-methylmalonic acid (MMA) | C6H12O2 | 116 | 3.91 |
| 5 | 12.92 | Maltol | C6H6O3 | 126 | 3.91 |
| 6 | 13.27 | 1-Butanol, 3-methyl, formate | C6H12O2 | 116 | 12.06 |
| 7 | 13.86 | 2,3-dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one (DDPM) | C6H8O4 | 144 | 8.73 |
| 8 | 15.56 | Cyclopenta[cd]pentalene | C10H6 | 126 | 2.10 |
| 9 | 18.79 | 2-methoxy vinylphenol | C9H10O2 | 150 | 3.25 |
| 10 | 20.44 | 4-O-alpha-D-Glucopyranosyl-D-glucose | C12H22O11 | 342 | 1.03 |
| 11 | 22.49 | 2,2,3,3,4,4 Hexadeutero octadecanal | C18H30D6O | 274 | 4.04 |
| 12 | 23.09 | Mome inositol | C7H14O6 | 194 | 7.68 |
| 13 | 23.51 | alpha-D-glucopyranose-4-O-alpha-d-glactopyranosyl | C12H22O11 | 342 | 2.61 |
| 14 | 24.29 | Generyl isovalerate | C15H26O2 | 238 | 1.51 |
| 15 | 24.84 | Hexopyranosyl-(1->3)hex-2-ulofuranosyl hexopyranoside | C18H32O16 | 504 | 2.28 |
| 16 | 25.12 | 1,3-Cyclohexanedione | C11H16O2 | 180 | 1.25 |
| 17 | 25.79 | D-Mannose | C6H12O6 | 180 | 1.31 |
| 18 | 26.00 | alpha-D-Glucopyranoside, O-à-D-glucopyranosyl-(1.fwdarw.3)-alpha-D-fructofuranosyl | C18H32O16 | 504 | 1.05 |
| 19 | 26.70 | 4-C-Methyl-myo-inositol | C7H14O6 | 194 | 4.06 |
| 20 | 28.18 | Octadecanoic acid 9,10-dichloro-,methyl ester | C19H36Cl2O2 | 366 | 1.07 |
| 21 | 30.98 | 6,8-Nonadien-2-one, 8-methyl-5-(1-methylethyl)-, (E)- | C13H22O | 194 | 2.06 |
| 22 | 34.65 | Spiro [4.5]decan-7-one, 1,8-dimethyl-8,9-epoxy-4-isopropyl- | C15H24O2 | 236 | 1.35 |
| 23 | 56.08 | Stigmast-5-en-3-ol, (3.beta.,24S) | C29H50O | 414 | 2.18 |
| Sample | Concentration (mg/mL) | Remaining DPPH (%) | Scavenging Activity (%) | IC50 (mg/mL) |
|---|---|---|---|---|
| Fungal filtrate | 1.988 | 55.01 ± 1.55 | 44.99 ± 1.80 | 2.277 ± 0.129 |
| 0.994 | 69.23 ± 1.73 | 30.77 ± 1.70 | ||
| 0.497 | 80.17 ± 2.05 | 19.83 ± 2.08 | ||
| 0.248 | 90.49 ± 1.67 | 8.83 ± 2.05 | ||
| Ascorbic acid | 0.062 | 15.27 ± 1.08 | 84.73 ± 1.05 | 0.022 ± 0.250 |
| 0.031 | 39.08 ± 1.44 | 60.92 ± 1.45 | ||
| 0.016 | 61.07 ± 1.19 | 38.93 ± 1.28 | ||
| 0.008 | 74.81 ± 1.05 | 25.19 ± 1.08 |
| Cell Line Type | The Fungal Filtrate (µg/mL) | Doxorubicin (Control; µg/mL) |
|---|---|---|
| Normal skin fibroblast | >200 | 100 ± 1.1 |
| Caucasian breast adenocarcinoma | 61.40 ± 1.7 | 4.17 ± 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alrdahe, S.S.; Moussa, Z.; Alanazi, Y.F.; Alrdahi, H.; Saber, W.I.A.; Darwish, D.B.E. Optimization of Biodegradation of Common Bean Biomass for Fermentation Using Trichoderma asperellum WNZ-21 and Artificial Neural Networks. Fermentation 2024, 10, 354. https://doi.org/10.3390/fermentation10070354
Alrdahe SS, Moussa Z, Alanazi YF, Alrdahi H, Saber WIA, Darwish DBE. Optimization of Biodegradation of Common Bean Biomass for Fermentation Using Trichoderma asperellum WNZ-21 and Artificial Neural Networks. Fermentation. 2024; 10(7):354. https://doi.org/10.3390/fermentation10070354
Chicago/Turabian StyleAlrdahe, Salma Saleh, Zeiad Moussa, Yasmene F. Alanazi, Haifa Alrdahi, WesamEldin I. A. Saber, and Doaa Bahaa Eldin Darwish. 2024. "Optimization of Biodegradation of Common Bean Biomass for Fermentation Using Trichoderma asperellum WNZ-21 and Artificial Neural Networks" Fermentation 10, no. 7: 354. https://doi.org/10.3390/fermentation10070354