
Insight into the Effects of Norfloxacin on Bacterial Community and Antibiotic Resistance Genes during Chicken Manure Composting


Abstract
1. Introduction
2. Materials and Methods
2.1. Composting Raw Materials
2.2. Experimental Design
2.3. Physicochemical Parameters Analysis and Germination Index
2.4. Extraction and Analysis of NOR
2.5. DNA Extraction and High-Throughput Sequencing Analysis
2.6. ARGs Analysis
2.7. Statistical Analysis
3. Results and Discussion
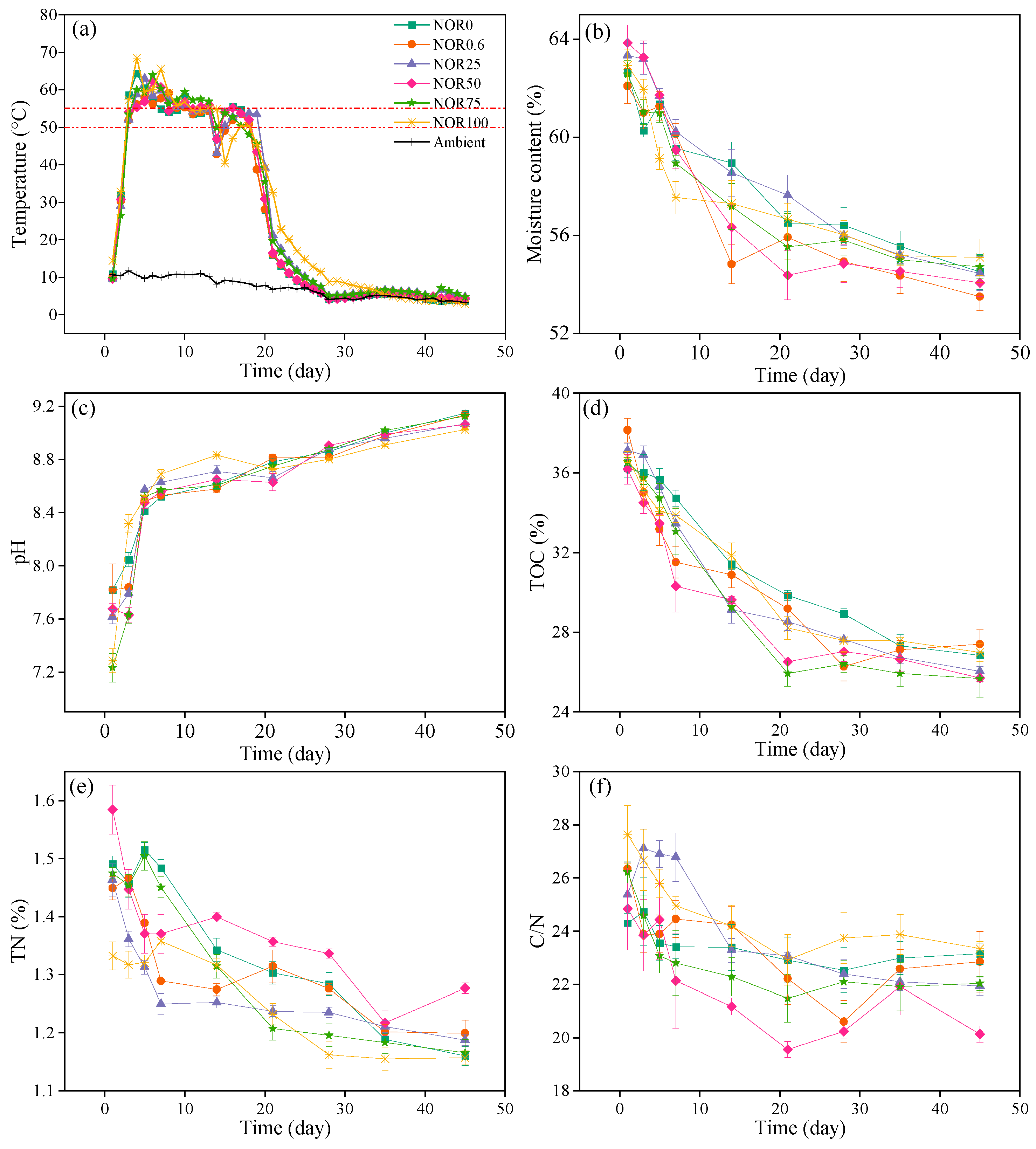
3.1. Variations in Physicochemical Properties of Composting
3.2. Change in Germination Index during Composting
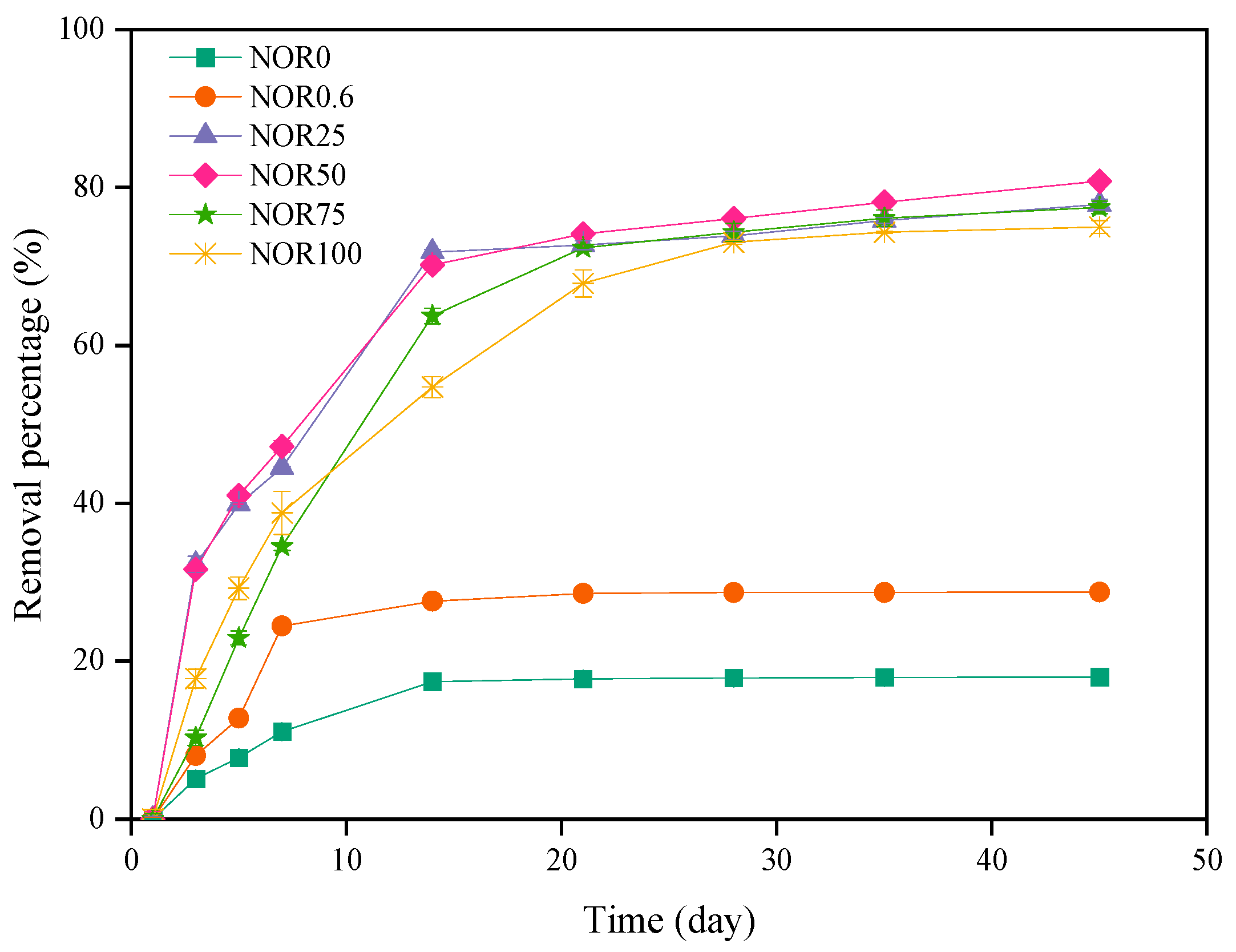
3.3. Fate of Norfloxacin during Composting
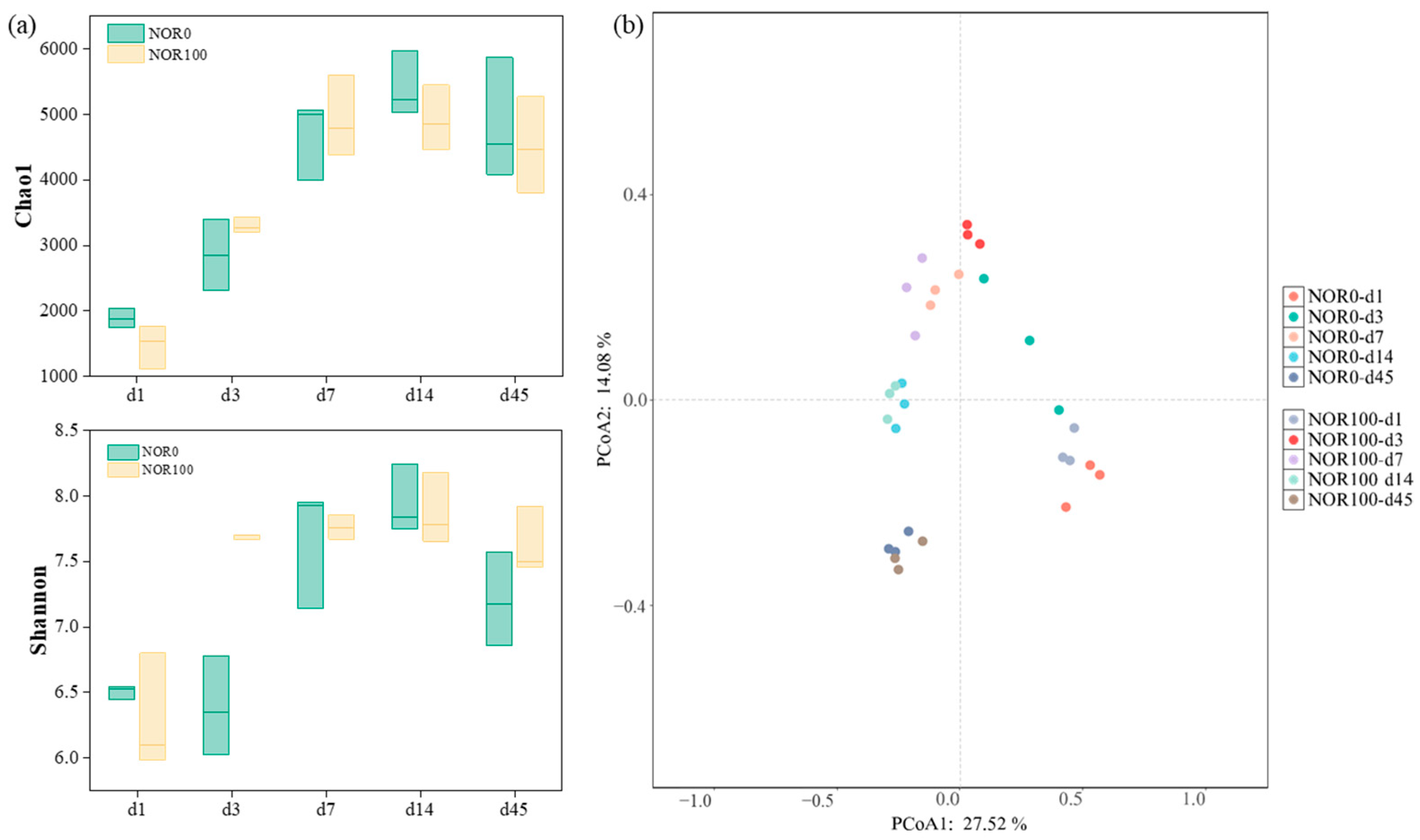
3.4. Succession of Bacterial Community during Composting
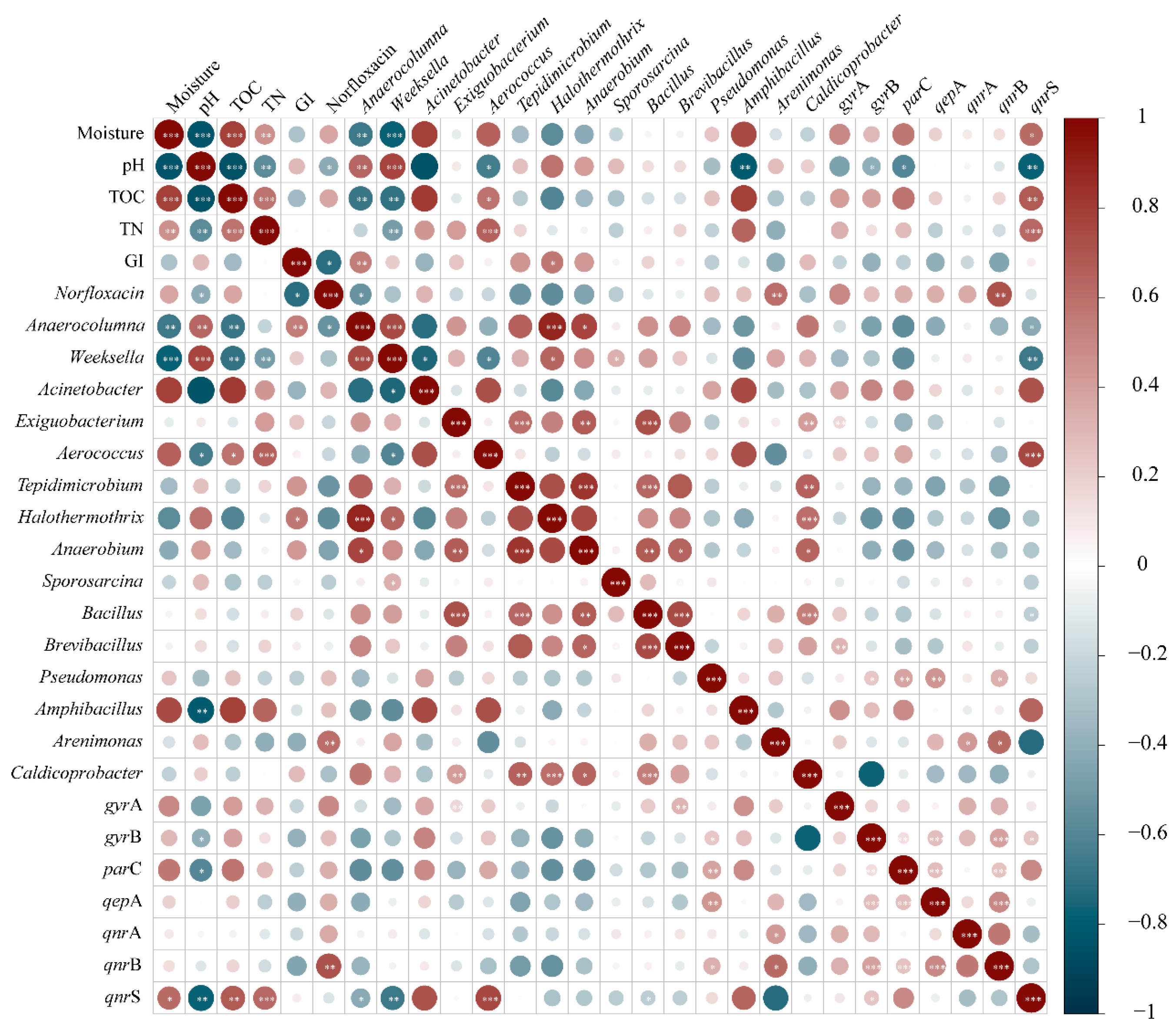
3.5. Behavior of Targeted Fluoroquinolone Resistance Genes and the Relationship with Composting Environmental Factors and Bacterial Communities
4. Conclusions and Future Research Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Namrata, R.; Sruthi, A.A.; Chandrasekaran, N.; Amitava, M.; Krishnan, K. A comprehensive update on antibiotics as an emerging water pollutant and their removal using nano-structured photocatalysts. J. Environ. Chem. Eng. 2021, 9, 104796. [Google Scholar]
- Shi, J.Y.; Dong, Y.B.; Shi, Y.Y.; Yin, T.T.; He, W.; An, T.Y.; Tang, Y.L.; Hou, X.W.; Chong, S.J.; Chen, D.N.; et al. Groundwater antibiotics and microplastics in a drinking-water source area, northern China: Occurrence, spatial distribution, risk assessment, and correlation. Environ. Res. 2022, 210, 112855. [Google Scholar] [CrossRef] [PubMed]
- Jurado, A.; Margareto, A.; Pujades, E.; Vázquez-Suñé, E.; Diaz-Cruz, M.S. Fate and risk assessment of sulfonamides and metabolites in urban groundwater. Environ. Pollut. 2020, 267, 115480. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, L.; Yen, H.; Zhao, F.K.; Wang, X.M.; Zhou, T.H.; Feng, Q.Y.; Chen, L.D. Occurrence, spatial distribution and ecological risks of antibiotics in soil in urban agglomeration. J. Environ. Sci. 2023, 125, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Nkoh, J.N.; Shang, C.J.; Okeke, E.S.; Ejeromedoghene, O.; Oderinde, O.; Etafo, N.O.; Mgbechidinma, C.L.; Bakare, O.C.; Meugang, E.F. Antibiotics soil-solution chemistry: A review of environmental behavior and uptake and transformation by plants. J. Environ. Manage 2024, 354, 120312. [Google Scholar] [CrossRef] [PubMed]
- Sunidhi, B.; Subhankar, C. Fluoroquinolone antibiotics: Occurrence, mode of action, resistance, environmental detection, and remediation—A comprehensive review. Environ. Pollut. 2022, 315, 120440. [Google Scholar]
- Gbylik-Sikorska, M.; Posyniak, A.; Sniegocki, T.; Zmudzki, J. Liquid chromatography–tandem mass spectrometry multiclass method for the determination of antibiotics residues in water samples from water supply systems in food-producing animal farms. Chemosphere 2015, 119, 8–15. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Ying, G.G.; Pan, C.G.; Liu, Y.S.; Zhao, J.L. Comprehensive evaluation of antibiotics emission and fate in the river basins of China: Source analysis, multimedia modeling, and linkage to bacterial resistance. Environ. Sci. Technol. 2015, 49, 6772–6782. [Google Scholar] [CrossRef]
- Wang, R.; Feng, F.; Chai, Y.; Meng, X.S.; Sui, Q.W.; Chen, M.X.; Wei, Y.S.; Qi, K.M. Screening and quantitation of residual antibiotics in two different swine wastewater treatment systems during warm and cold seasons. Sci. Total Environ. 2019, 660, 1542–1554. [Google Scholar] [CrossRef]
- Xiong, W.G.; Sun, Y.X.; Ding, X.Y.; Wang, M.Z.; Zeng, Z.L. Selective pressure of antibiotics on ARGs and bacterial c ommunities in manure-polluted fresh water-sediment microcosms. Front. Microbiol. 2015, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.J.; Qian, X.; Gu, J.; Wang, X.J.; Gao, H. Effects of oxytetracycline on the abundance and community structure of nitrogen-fixing bacteria during cattle manure composting. Bioresour. Technol. 2016, 216, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.Y.; Liu, B.T.; Imtiaz, A.; Yang, J.; Zhang, B. Composting effect and antibiotic removal under a new temperature control strategy. Waste Manag. 2022, 153, 89–98. [Google Scholar] [CrossRef]
- Gaballah, M.S.; Guo, J.; Sun, H.; Aboagye, D.; Sobhi, M.; Muhmood, A.; Dong, R. A review targeting veterinary antibiotics removal from livestock manure management systems and future outlook. Bioresour. Technol. 2021, 333, 125069. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, X. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar. Bioresour. Technol. 2014, 171, 274–284. [Google Scholar] [CrossRef]
- Boonsaner, M.; Hawker, D.W. Transfer of oxytetracycline from swine manure to three different aquatic plants: Implications for human exposure. Chemosphere 2015, 122, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, S.; Chen, Z.; Wen, Q.; Wang, Y. Maturity and security assessment of pilot-scale aerobic co-composting of penicillin fermentation dregs (PFDs) with sewage sludge. Bioresour. Technol. 2016, 204, 185–191. [Google Scholar] [CrossRef]
- Liu, B.; Li, Y.X.; Zhang, X.L.; Feng, C.H.; Gao, M.; Shen, Q. Effects of composting process on the dissipation of extractable sulfonamides in swine manure. Bioresour. Technol. 2015, 175, 284–290. [Google Scholar] [CrossRef]
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef]
- Kim, K.R.; Owens, G.; Ok, Y.S.; Park, W.; Lee, D.B.; Kwon, S.I. Decline in extractable antibiotics in manure-based composts during composting. Waste Manage. 2012, 32, 110–116. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, G.Z.; Liu, Y.W.; Cheng, D.M.; Fan, S.H.; Zhao, Q.S.; Xue, J.M.; Zhang, S.Q.; Li, Z.J. The impacts of oxytetracycline on humification during manure composting can be alleviated by initial moisture contents as illustrated by NMR. J Integr. Agr. 2020, 19, 2–13. [Google Scholar] [CrossRef]
- Liao, H.P.; Zhao, Q.; Cui, P.; Chen, Z.; Yu, Z.; Stefan, G.; Ville-Petri, F.; Zhou, S.G. Efficient reduction of antibiotic residues and associated resistance genes in tylosin antibiotic fermentation waste using hyperthermophilic composting. Environ. Int. 2019, 133 Pt B, 105203. [Google Scholar] [CrossRef]
- Tong, Z.Y.; Liu, F.W.; Sun, B.; Tian, Y.; Zhang, J.Z.; Duan, J.Z.; Bi, W.L.; Qin, J.M.; Xu, S.Z. Effect of biochars with different particle sizes on fates of antibiotics and antibiotic resistance genes during composting of swine manure. Bioresour. Technol. 2023, 370, 128542. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.H.; Wang, X.X.; Feng, Y.; Ren, Y.F.; He, J.Y.; Li, Z.J. Dynamics of physicochemical properties, microbial composition, and antibiotic and antibiotic resistance genes during chicken manure composting with strain T4. J. Soils Sediments 2024, 24, 1750–1763. [Google Scholar] [CrossRef]
- Ezzariai, A.; Hafidi, M.; Khadra, A.; Aemig, Q.; El Felsa, L.; Barret, M.; Merlina, G.; Patureau, D.; Pinelli, E. Human and veterinary antibiotics during composting of sludge or manure: Global perspectives on persistence, degradation, and resistance genes. J. Hazard. Mater. 2018, 359, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Ezzariai, A.; Barret, M.; Merlina, G.; Pinelli, E.; Hafidi, M. Evaluation of the antibiotics effects on the physical and chemical parameters during the co-composting of sewage sludge with palm wastes in a bioreactor. Waste Manage. 2017, 68, 388–397. [Google Scholar] [CrossRef]
- Li, C.N.; Li, H.Y.; Yao, T.; Su, M.; Ran, F.; Han, B.; Li, J.H.; Lan, X.J.; Zhang, Y.C.; Yang, X.M.; et al. Microbial inoculation influences bacterial community succession and physicochemical characteristics during pig manure composting with corn straw. Bioresour. Technol. 2019, 289, 121653. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.M.; Feng, Y.; Liu, Y.W.; Xue, J.M.; Li, Z.J. Dynamics of oxytetracycline, sulfamerazine, and ciprofloxacin and related antibiotic resistance genes during swine manure composting. J. Environ. Manage. 2019, 230, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wei, C.J.; Zhang, W.J.; Liu, Y.W.; Li, Z.J.; Hu, H.Y.; Xue, J.M.; Davis, M. A simple and economic method for simultaneous determination of 11 antibiotics in manure by solid-phase extraction and high-performance liquid chromatography. J. Soils Sediments 2016, 16, 2242–2251. [Google Scholar] [CrossRef]
- Liu, Y.W.; Feng, Y.; Cheng, D.M.; Xue, J.M.; Wakelin, S.A.; Li, Z.J. Dynamics of bacterial composition and the fate of antibiotic resistance genes and mobile genetic elements during the co-composting with gentamicin fermentation residue and lovastatin fermentation residue. Bioresour. Technol. 2018, 261, 249–256. [Google Scholar] [CrossRef]
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.; Fierer, N.; Gonzalez Pena, A.; Goodrich, J.; Gordon, J.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.J.; Sun, L.F.; Zhang, Y.F.; Zhang, X.L.; Qiao, J.J. Conversion of spent mushroom substrate to biofertilizer using a stress-tolerant phosphatesolubilizing Pichia farinose FL7. Bioresour. Technol. 2012, 111, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Wu, Y.Q.; Wen, Q.X.; Bao, H.Y.; Fu, Q.Q. Insight into the effects of sulfamethoxazole and norfloxacin on nitrogen transformation functional genes during swine manure composting. Bioresour. Technol. 2020, 297, 122463. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.F.; Ying, G.G.; Liu, S.; Zhou, L.J.; Zhao, J.L.; Tao, R.; Peng, P.A. Biological degradation and microbial function effect of norfloxacin in a soil under different conditions. J. Environ. Sci. Health B 2012, 47, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Li, X.; Cui, P.; Lin, J.; Zeng, J.; Lin, H.; Zhou, S. Nitrification plays a key role in N2O emission in electric-field assisted aerobic composting. Bioresour. Technol. 2020, 297, 122470. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Ma, B.R.; She, Z.L.; Guo, L.; Zhao, Y.G.; Jin, C.J.; Gao, M.C. The presence of norfloxacin inhibited the volatilization of NH4+-N in the form of NH3 and promote its conversion to NO3−-N. Environ. Technol. Inno. 2020, 18, 100726. [Google Scholar] [CrossRef]
- Qu, Y.; Qu, J.; Yan, W.; Yue, T.; Zhang, Q.; Yi, W.; Liu, X.; Sun, Y. Influence of biochar on physico-chemical, microbial community and maturity during biogas residue aerobic composting process. Fermentation 2022, 8, 623. [Google Scholar] [CrossRef]
- Shi, H.L.; Wang, X.C.; Li, Q.; Jiang, S.Q. Effects of elevated tetracycline concentrations on aerobic composting of human feces: Composting behavior and microbial community succession. Indian J. Microbiol. 2018, 58, 423–432. [Google Scholar] [CrossRef]
- Xu, X.Y.; Ma, W.J.; Zhou, K.X.; An, B.Y.; Huo, M.X.; Lin, X.D.; Wang, L.; Wang, H.Y.; Liu, Z.L.; Cheng, G.Y.; et al. Effects of composting on the fate of doxycycline, microbial community, and antibiotic resistance genes in swine manure and broiler manure. Sci. Total Environ. 2022, 832, 155039. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Y.; Ye, C.; He, X.; Zhang, S. Fate of antibiotics and antibiotic resistance genes during aerobic co-composting of food waste with sewage sludge. Sci. Total Environ. 2021, 784, 146950. [Google Scholar] [CrossRef]
- Zhang, M.; He, L.Y.; Liu, Y.S.; Zhao, J.L.; Liu, W.R.; Zhang, J.N.; Chen, J.; He, L.K.; Zhang, Q.Q.; Ying, G.G. Fate of veterinary antibiotics during animal manure composting. Sci. Total Environ. 2019, 650, 1363–1370. [Google Scholar] [CrossRef] [PubMed]
- Ezugworie, F.N.; Igbokwe, V.C.; Onwosi, C.O. Proliferation of antibiotic-resistant microorganisms and associated genes during composting: An overview of the potential impacts on public health, management and future. Sci. Total Environ. 2021, 784, 147191. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Feng, Y.; Cheng, D.M.; Xue, J.M.; Wakelin, S.A.; Hu, H.Y.; Li, Z.J. Gentamicin degradation and changes in fungal diversity and physicochemical properties during composting of gentamicin production residue. Bioresour. Technol. 2017, 244, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Bourdat-Deschamps, M.; Ferhi, S.; Bernet, N.; Feder, F.; Crouzet, O.; Patureau, D.; Montenach, D.; Moussard, G.D.; Mercier, V.; Benoit, P. Fate and impacts of pharmaceuticals and personal care products after repeated applications of organic waste products in long-term field experiments. Sci. Total Environ. 2017, 607, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, M.; Nandal, M.; Khosla, B. Microbes as vital additives for solid waste composting. Heliyon 2020, 6, e03343. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Peng, Y.; Ye, C.; Zhang, S. Effects of antibiotics on hydrolase activity and structure of microbial community during aerobic co-composting of food waste with sewage sludge. Bioresour. Technol. 2021, 321, 124506. [Google Scholar] [CrossRef]
- Zhao, C.X.; Xin, L.Q.; Xu, X.K.; Qin, Y.; Wu, W.X. Dynamics of antibiotics and antibiotic resistance genes in four types of kitchen waste composting processes. J. Hazard. Mater. 2022, 424 Pt C, 127526. [Google Scholar] [CrossRef]
- Wei, Y.; Liang, Z.; Zhang, Y. Evolution of physicochemical properties and bacterial community in aerobic composting of swine manure based on a patent compost tray. Bioresour. Technol. 2022, 343, 126136. [Google Scholar] [CrossRef]
- Li, X.; Wang, P.; Chu, S.; Xu, Y.; Su, Y.; Wu, D.; Xie, B. Short-term biodrying achieves compost maturity and significantly reduces antibiotic resistance genes during semi-continuous food waste composting inoculated with mature compost. J. Hazard. Mater. 2022, 427, 127915. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.; Lin, C.; Hoang, H.G.; Nguyen, M.T.; Kaewlaoyoong, A.; Cheruiyot, N.K.; Bui, X.T.; Vu, C.T. Biodegradation of dioxin-contaminated soil via composting: Identification and phylogenetic relationship of bacterial communities. Environ. Technol. Innovation 2020, 19, 101023. [Google Scholar] [CrossRef]
- Qian, X.; Sun, W.; Gu, J.; Wang, X.; Zhang, Y.; Duan, M.; Li, H.; Zhang, R. Reducing antibiotic resistance genes, integrons, and pathogens in dairy manure by continuous thermophilic composting. Bioresour. Technol. 2016, 220, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.C.; Qin, H.Y.; Zhang, H.; Luo, Y.H.; Ru, Y.N.; Li, J.R.; Kim, W.S.; Wang, L.S.; Yu, X.N.; Guo, W.H. Variations in antibiotic resistance genes and removal mechanisms induced by C/N ratio of substrate during composting. Sci. Total Environ. 2021, 798, 149288. [Google Scholar] [CrossRef]
- Wen, X.; Chen, M.J.; Ma, B.H.; Xu, J.J.; Zhu, T.; Zou, Y.D.; Liao, X.D.; Wang, Y.; Anja, W.; Wu, Y.B. Removal of antibiotic resistance genes during swine manure composting is strongly impaired by high levels of doxycycline residues. Waste Manag. 2024, 177, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Koyama, S.; Murase, T.; Ozaki, H. Research Note: Longitudinal monitoring of chicken houses in a commercial layer farm for antimicrobial resistance in Escherichia coli with special reference to plasmid-mediated quinolone resistance. Poultry Sci. 2020, 99, 1150–1155. [Google Scholar] [CrossRef]
- Cui, M.Q.; Zhang, P.; Li, J.Y.; Sun, C.T.; Song, L.; Zhang, C.P.; Zhao, Q.; Wu, C.M. Prevalence and characterization of fluoroquinolone resistant Salmonella isolated from an integrated broiler chicken supply chain. Front. Microbiol. 2019, 10, 1865. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | MC/% | pH | TOC/% | TN/% | C/N |
|---|---|---|---|---|---|
| Chicken manure | 46.72 | 8.34 | 28.64 | 3.28 | 8.73 |
| Wheat straw | 8.77 | 7.56 | 44.63 | 1.13 | 39.50 |
| Sawdust | 8.50 | 6.20 | 49.27 | 0.19 | 259.31 |
| Treatment | Fitted Equation | R a | k b (day−1) | t½ c (day) |
|---|---|---|---|---|
| NOR0 | C = C0 e−0.0032t − 0.0908 | 0.8041 | 0.0032 | 216.56 |
| NOR0.6 | C = C0 e−0.0049t − 0.1745 | 0.7357 | 0.0049 | 141.43 |
| NOR25 | C = C0 e−0.0267t − 0.5136 | 0.8862 | 0.0267 | 25.96 |
| NOR50 | C = C0 e−0.0300t − 0.4952 | 0.9257 | 0.0300 | 23.10 |
| NOR75 | C = C0 e−0.0339t − 0.2520 | 0.9113 | 0.0339 | 20.44 |
| NOR100 | C = C0 e−0.0299t − 0.2876 | 0.9396 | 0.0299 | 23.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Y.; Shi, H.; Fei, Y.; Zhao, Q.; Li, Z. Insight into the Effects of Norfloxacin on Bacterial Community and Antibiotic Resistance Genes during Chicken Manure Composting. Fermentation 2024, 10, 366. https://doi.org/10.3390/fermentation10070366
Feng Y, Shi H, Fei Y, Zhao Q, Li Z. Insight into the Effects of Norfloxacin on Bacterial Community and Antibiotic Resistance Genes during Chicken Manure Composting. Fermentation. 2024; 10(7):366. https://doi.org/10.3390/fermentation10070366
Chicago/Turabian StyleFeng, Yao, Huading Shi, Yang Fei, Quansheng Zhao, and Zhaojun Li. 2024. "Insight into the Effects of Norfloxacin on Bacterial Community and Antibiotic Resistance Genes during Chicken Manure Composting" Fermentation 10, no. 7: 366. https://doi.org/10.3390/fermentation10070366
APA StyleFeng, Y., Shi, H., Fei, Y., Zhao, Q., & Li, Z. (2024). Insight into the Effects of Norfloxacin on Bacterial Community and Antibiotic Resistance Genes during Chicken Manure Composting. Fermentation, 10(7), 366. https://doi.org/10.3390/fermentation10070366