

Correlation between the Characteristic Flavour and Microbial Community of Xuanwei Ham after Ripening
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Instruments and Equipment
2.3. Experimental Methods
2.3.1. Sample Processing
2.3.2. Free Amino Acid Content Determination
2.3.3. Taste Active Value (TAV) Calculation
2.3.4. Volatile Flavour Content Determination
2.3.5. Electronic Nose Odour Fingerprint Extraction
2.4. Experimental Procedures of Metagenomic Sequencing
2.5. Data Processing
3. Results
3.1. Free Amino Acid Content and TAV Value of Differently Aged Xuanwei Hams
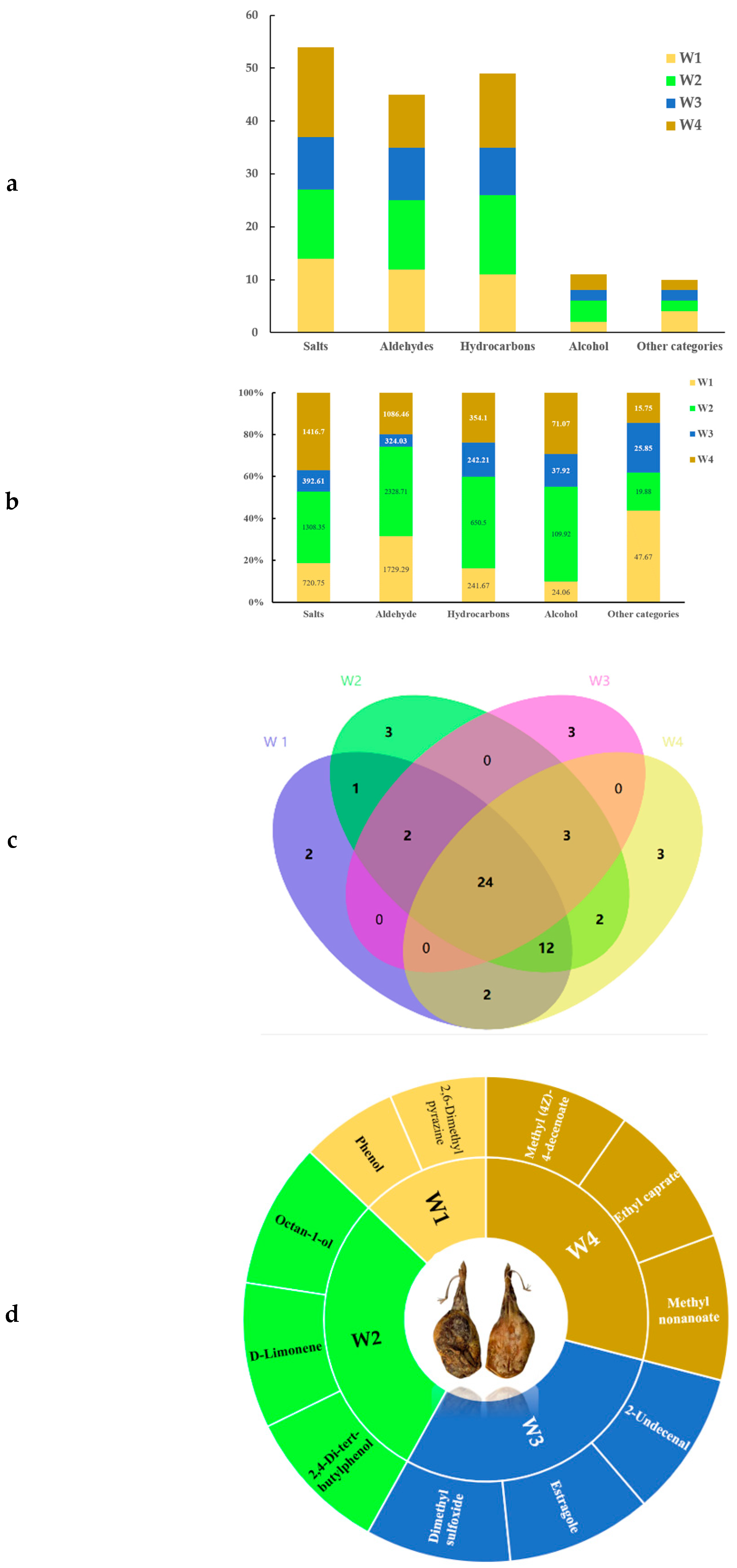
3.2. VOCs in Xuanwei Ham of Different Vintages
3.3. Electronic Nose Analysis of Xuanwei Ham in Different Years
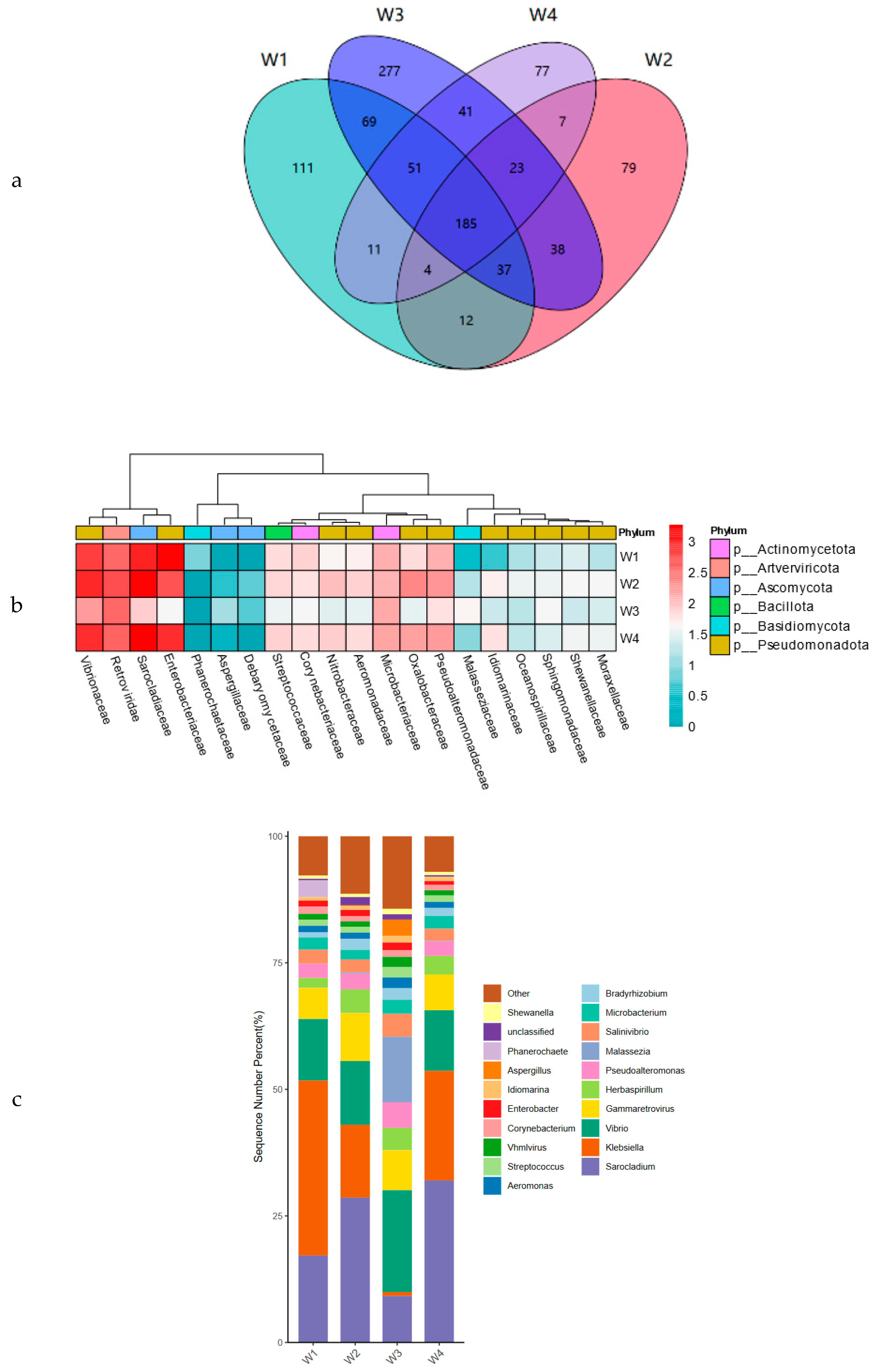
3.4. Microbial Community Analysis
3.4.1. Macrogenomic Data Overview
3.4.2. Bacterial Microbiological Composition of Xuanwei Ham
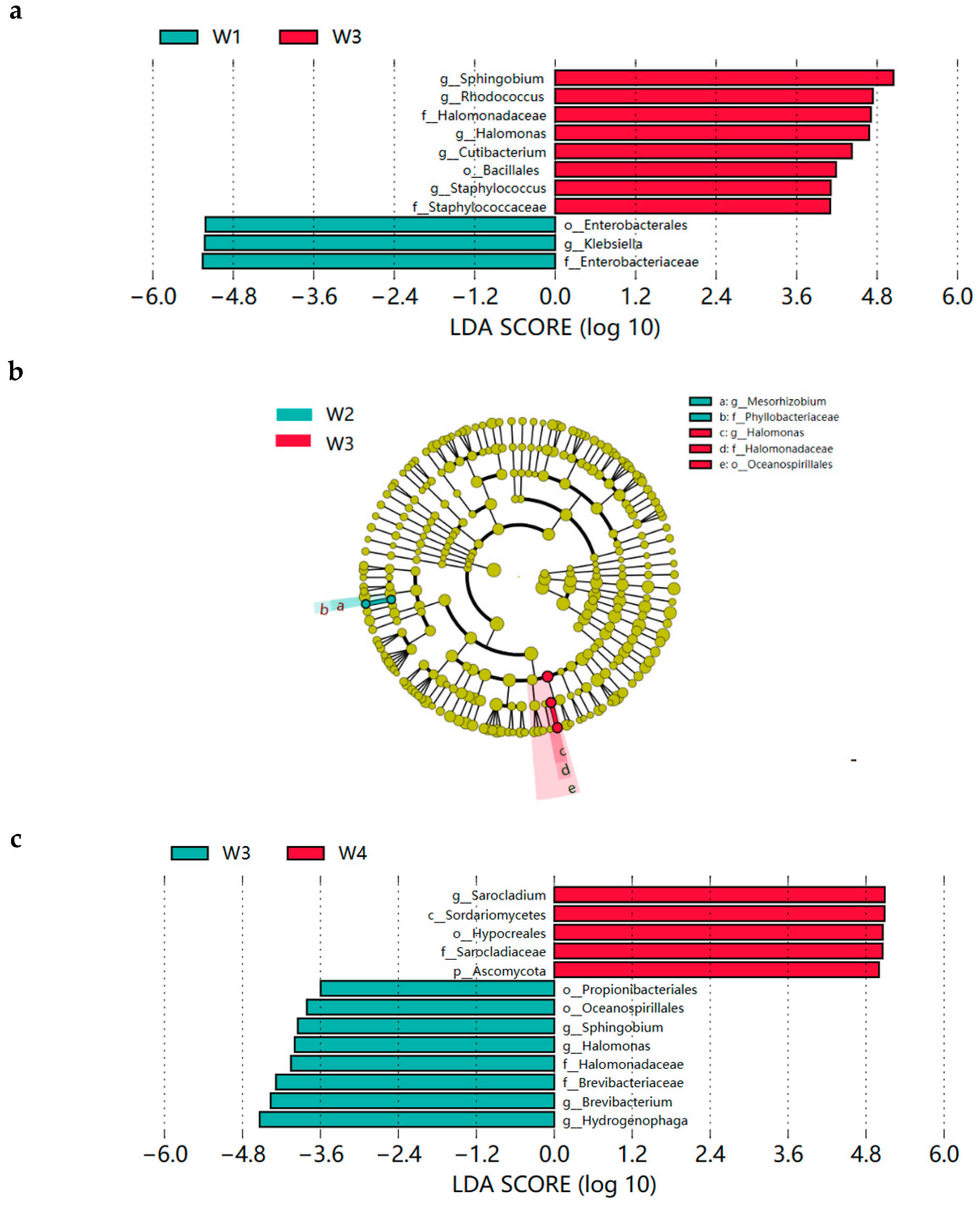
3.4.3. Xuanwei Ham LEfSe Analysis
3.4.4. Correlation Analysis of Phylum and Genus Microbiota with Free Amino Acids, VOCs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, Y. Modernisation of traditional Chinese flavour meat products. Meat Res. 1998, 2, 3–6. [Google Scholar]
- Wu, W.; Zhou, Y.; Wang, G.; Zhu, R.; Ge, C.; Liao, G. Changes in the physicochemical properties and volatile flavor compounds of dry-cured Chinese Laowo ham during processing. J. Food Process. Preserv. 2020, 44, e14593. [Google Scholar] [CrossRef]
- Huang, A.X.; Ge, C.R.; Huang, Q.C. The study of ingredients and processing techniques of Xuanwei style ham. J. Food Process. Preserv. 2010, 34, 136–148. [Google Scholar] [CrossRef]
- Yi, Y.; Wang, X.; Xiang, J.; Zang, L.; Chen, C.; Chen, D.; Wang, X.; Cao, J. Effects of curing agent and rinsing process on the flavour of Xuanwei ham. Food Ferment. Ind. 2024, 50, 85–92. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Pan, D.; Geng, F.; Zhou, C.; Cao, J. Insight into the relationship between microorganism communities and flavor quality of Chinese dry-cured boneless ham with different quality grades. Food Biosci. 2022, 50, 102174. [Google Scholar] [CrossRef]
- Wang, H.; Yin, X.; Zhang, L.; Wang, X.; Zhang, J.; Wen, R.; Cao, J. Insight into the Relationship between the Causes of Off-Odour and Microorganism Communities in Xuanwei Ham. Foods 2024, 13, 776. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, Y.; Lu, S.; Wang, J.; Fu, H.; Gu, B.; Lyu, B.; Wang, Q. Changes in proteolysis, protein oxidation, flavor, color and texture of dry-cured mutton ham during storage. LWT 2021, 149, 111860. [Google Scholar] [CrossRef]
- Zhu, Y.; Guo, Y.; Yang, F.; Zhou, C.; Tang, C.; Zhou, G. Combined application of high-throughput sequencing and UHPLC-Q/TOF-MS-based metabolomics in the evaluation of microorganisms and metabolites of dry-cured ham of different origins. Int. J. Food Microbiol. 2021, 359, 109422. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.R.; Wang, G.Y.; Sun, Y.H.; Ge, C.R.; Liao, G.Z. Changes in physicochemical parameters, free fatty acid profile and water-soluble compounds of Yunnan dry-cured beef during processing. J. Food Process. Preserv. 2020, 44, e14380. [Google Scholar] [CrossRef]
- Virgili, R.; Saccani, G.; Gabba, L.; Tanzi, E.; Soresi Bordini, C. Changes of free amino acids and biogenic amines during extended ageing of Italian dry-cured ham. Lwt–Food Sci. Technol. 2007, 40, 871–878. [Google Scholar] [CrossRef]
- Li, C.; Zheng, Z.; Wang, G.; Chen, G.; Zhou, N.; Zhong, Y.; Yang, Y.; Wu, H.; Yang, C.; Liao, G. Revealing the intrinsic relationship between microbial communities and physicochemical properties during ripening of Xuanwei ham. Food Res. Int. 2024, 186, 114377. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Pei, H.; Liu, R.; Chen, L.; Gao, X.; Gu, Y.; Hou, Q.; Yin, Y.; Yu, H.; Wu, M.; et al. Effects of Lactobacillus plantarum NJAU-01 from Jinhua ham on the quality of dry-cured fermented sausage. LWT 2019, 101, 513–518. [Google Scholar] [CrossRef]
- Wang, Y.; Li, F.; Chen, J.; Sun, Z.; Wang, F.; Wang, C.; Fu, L. High-throughput sequencing-based characterization of the predominant microbial community associated with characteristic flavor formation in Jinhua Ham. Food Microbiol. 2021, 94, 103643. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, Y.; Wu, Y.; Li, C.; Li, L.; Zhao, Y.; Hu, X.; Wei, Y.; Huang, H. Comparison of the microbial community and flavor compounds in fermented mandarin fish (Siniperca chuatsi): Three typical types of Chinese fermented mandarin fish products. Food Res. Int. 2021, 144, 110365. [Google Scholar] [CrossRef] [PubMed]
- Zhong, A.; Chen, W.; Duan, Y.; Li, K.; Tang, X.; Tian, X.; Wu, Z.; Li, Z.; Wang, Y.; Wang, C. The potential correlation between microbial communities and flavors in traditional fermented sour meat. LWT 2021, 149, 111873. [Google Scholar] [CrossRef]
- Deng, J.; Xu, H.; Li, X.; Wu, Y.; Xu, B. Correlation of characteristic flavor and microbial community in Jinhua ham during the post-ripening stage. LWT 2022, 171, 114067. [Google Scholar] [CrossRef]
- Li, P.; Bao, Z.; Wang, Y.; Su, X.; Zhou, H.; Xu, B. Role of microbiota and its ecological succession on flavor formation in traditional dry-cured ham: A review. Crit. Rev. Food Sci. Nutr. 2023, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Duan, R.; Wang, Y.; He, Y.; Li, C.; Shen, X.; Li, Y. Effects of different drying temperatures on the profile and sources of flavor in semi-dried golden pompano (Trachinotus ovatus). Food Chem. 2023, 401, 134112. [Google Scholar] [CrossRef]
- Zhu, N.; Wang, S.; Zhao, B.; Zhang, S.; Zang, M.; Wu, Q.; Li, S.; Qiao, X. Label-free proteomic strategy to identify proteins associated with quality properties in sauced beef processing. Food Biosci. 2021, 42, 101163. [Google Scholar] [CrossRef]
- Xu, X.; You, M.; Song, H.; Gong, L.; Pan, W. Investigation of umami and kokumi taste-active components in bovine bone marrow extract produced during enzymatic hydrolysis and Maillard reaction. Int. J. Food Sci. Technol. 2018, 53, 2465–2481. [Google Scholar] [CrossRef]
- Cao, C.; Feng, M.; Sun, J.; Xu, X.; Zhou, G. Effects of functional fermentation agents on oxidative stability and volatile flavour substances of fermented sausages. Food Sci. 2019, 40, 106–113. [Google Scholar] [CrossRef]
- Marcel, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Eur. Mol. Biol. Netw. 2011, 17, 1. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Breitwieser, F.P.; Thielen, P.; Salzberg, S.L. Bracken: Estimating species abundance in metagenomics data. PeerJ Comput. Sci. 2017, 3, e104. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef]
- Brum, J.R.; Ignacio-Espinoza, J.C.; Roux, S.; Doulcier, G.; Acinas, S.G.; Alberti, A.; Sullivan, M.B.; Roux, S.; Doulcier, G.; Acinas, S.G.; et al. Patterns and ecological drivers of ocean viral communities. Science 2015, 348, 1261498. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, G.; Jiang, S.; Liu, Y.X. Wekemo Bioincloud: A user-friendly platform for meta-omics data analyses. iMeta 2024, 3, e175. [Google Scholar] [CrossRef]
- Li, Q. Identification of Characteristic Flavour Substances in Agaricus Bisporus Soup and Study on the Release of Flavour Substances during Simmering. Ph.D. Thesis, Jiangnan University, Wuxi, China, 2011. [Google Scholar]
- Liu, B.; Wang, X.; Zhao, W.; Li, H.; Li, H.; Zhang, J.; Shi, Q. Physicochemical and flavour properties of Norden ham from different sources and years. Meat Res. 2021, 35, 1–8. [Google Scholar] [CrossRef]
- Ur-Rehman, S.; Fox, P.F. Effect of added α-ketoglutaric acid, pyruvic acid or pyridoxal phosphate on proteolyis and quality of Cheddar cheese. Food Chem. 2002, 76, 21–26. [Google Scholar] [CrossRef]
- Zhao, J. Study on the Melad Reaction Involving Free Amino Acids during the Flavour Formation of Jinhua Ham. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2013. [Google Scholar]
- Michikawa, K.; Konosu, S. Sensory identification of effective components for masking bitterness of arginine in synthetic extract of scallop. In Proceedings of the Olfaction and Taste XI: Proceedings of the 11th International Symposium on Olfaction and Taste and of the 27th Japanese Symposium on Taste and Smell, Joint Meeting Held at Kosei-Nenkin Kaikan, Sapporo, Japan, 12–16 July 1993; Springer: Tokyo, Japan, 1994; p. 278. [Google Scholar] [CrossRef]
- Sforza, S.; Pigazzani, A.; Motti, M.; Porta, C.; Virgili, R.; Galaverna, G.; Dossena, A.; Marchelli, R. Oligopeptides and free amino acids in Parma hams of known cathepsin B activity. Food Chem. 2001, 75, 267–273. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, G.; Ma, Y.; Suo, H.; Wang, L. Effects of lactic acid bacteria complex on growth performance, slaughter performance, meat quality and volatile flavour composition of black Tibetan sheep. China Feed 2023, 21, 73–80. [Google Scholar] [CrossRef]
- Chen, Y. Analysis of Microflora of Low-Salt and High-Moisture Sichuan Bacon and the Effect of Pasteurisation Treatment on Its Storage Quality. Master’s Thesis, Sichuan Agricultural University, Yaan, China, 2022. [Google Scholar] [CrossRef]
- Reyes-Díaz, R.; González-Córdova, A.F.; del Carmen Estrada-Montoya, M.; Méndez-Romero, J.I.; Mazorra-Manzano, M.A.; Soto-Valdez, H.; Vallejo-Cordoba, B. Volatile and sensory evaluation of Mexican Fresco cheese as affected by specific wild Lactococcus lactis strains. J. Dairy Sci. 2020, 103, 242–253. [Google Scholar] [CrossRef]
- Papadopoulou, O.S.; Panagou, E.Z.; Mohareb, F.R.; Nychas, G.J.E. Sensory and microbiological quality assessment of beef fillets using a portable electronic nose in tandem with support vector machine analysis. Food Res. Int. 2013, 50, 241–249. [Google Scholar] [CrossRef]
- Rattray NJ, W.; Hamrang, Z.; Trivedi, D.K.; Goodacre, R.; Fowler, S.J. Taking your breath away: Metabolomics breathes life in to personalized medicine. Trends Biotechnol. 2014, 32, 538–548. [Google Scholar] [CrossRef]
- Lin, J.; Song, C.; Zhang, Y.; Zhang, Y.; Quan, W.; Shu, X. Comparative analysis of microbial diversity of five dry-cured hams from Yunnan. China Food Addit. 2023, 34, 233–241. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, L.; Jiang, Y.; Zhao, P. GC-MS analysis of aroma components of white-flowered papaya fruit. South. China Fruit Tree 2017, 46, 117–120. [Google Scholar] [CrossRef]
- Lin, J. Correlation between Microbial Community Structure and Flavour Substances in the Ripening Process of Traditional Mongolian Cheese. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2023. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FAA | Taste Contribution | Thresholds/ (mg/100 g) | W1 | W2 | W3 | W4 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Content/ (mg/100 g) | TAV | Content/ (mg/100 g) | TAV | Content/ (mg/100 g) | TAV | Content/ (mg/100 g) | TAV | |||
| Asp | Fresh/Sweet (+) | 100 | 213.40 ± 4.41 a | 2.13 | 167.61 ± 1.87 c | 1.68 | 66.32 ± 0.47 d | 0.66 | 180.21 ± 0.45 b | 1.80 |
| Glu | Fresh (+) | 30 | 434.65 ± 3.65 b | 14.49 | 370.64 ± 2.31 c | 12.35 | 157.67 ± 4.62 d | 5.26 | 452.99 ± 3.53 a | 15.10 |
| Ser | Sweet (+) | 150 | 191.68 ± 1.90 b | 1.28 | 177.65 ± 3.61 c | 1.18 | 67.05 ± 1.09 d | 0.45 | 216.27 ± 2.64 a | 1.44 |
| Gly | Sweet (+) | 130 | 189.87 ± 0.86 b | 1.46 | 178.94 ± 3.27 c | 1.38 | 54.37 ± 1.37 d | 0.42 | 197.76 ± 2.20 a | 1.52 |
| Thr | Sweet (+) | 260 | 194.56 ± 1.96 b | 0.75 | 171.33 ± 3.06 c | 0.66 | 54.56 ± 0.53 d | 0.21 | 210.65 ± 3.77 a | 0.81 |
| Ala | Sweet (+) | 60 | 138.75 ± 2.23 b | 2.31 | 164.80 ± 0.87 a | 2.75 | 121.12 ± 0.86 c | 2.02 | 63.82 ± 0.35 d | 1.06 |
| Sar | Sweet (\) | \ | 24.72 ± 0.72 a | \ | 19.25 ± 3.94 a | \ | 16.44 ± 0.51 a | \ | 17.81 ± 2.92 a | \ |
| Arg | Sweet/Bitter (+) | 50 | 248.02 ± 1.43 b | 4.96 | 212.58 ± 1.40 c | 4.25 | 86.94 ± 1.48 d | 1.74 | 279.66 ± 4.44 a | 5.59 |
| Val | Sweet/Bitter (+) | 40 | 208.82 ± 1.16 a | 5.22 | 192.27 ± 1.73 b | 4.81 | 73.73 ± 2.47 c | 1.84 | 213.88 ± 1.07 a | 5.35 |
| Pro | Sweet/Bitter (+) | 300 | 171.66 ± 2.03 b | 0.57 | 150.96 ± 2.14 c | 0.50 | 53.04 ± 2.10 d | 0.18 | 187.08 ± 2.81 a | 0.62 |
| Cys | Sweet/Bitter (−) | 250 | 7.58 ± 0.04 b | <0.1 | 7.76 ± 0.06 b | <0.1 | 1.75 ± 0.14 c | <0.1 | 8.90 ± 0.12 a | <0.1 |
| Lys | Sweet/Bitter (−) | 50 | 310.07 ± 3.12 b | 6.20 | 265.52 ± 5.72 c | 5.31 | 89.32 ± 3.11 d | 1.79 | 345.45 ± 11.37 a | 6.90 |
| His | Bitter (−) | 20 | 106.25 ± 3.48 a | 5.31 | 99.12 ± 3.77 a | 4.96 | 29.78 ± 0.12 b | 1.49 | 106.97 ± 4.26 a | 5.35 |
| Tyr | Bitter (−) | \ | 58.65 ± 0.30 a | \ | 57.89 ± 1.49 a | \ | 27.92 ± 0.32 c | \ | 53.75 ± 0.38 b | \ |
| Met | Bitter (\) | 30 | 107.77 ± 1.71 b | 3.59 | 87.59 ± 1.10 c | 2.92 | 29.49 ± 0.20 d | 0.98 | 121.76 ± 0.62 a | 4.06 |
| Nva | Bitter (\) | \ | 11.99 ± 0.43 c | \ | 17.77 ± 0.16 a | \ | 15.86 ± 0.22 b | \ | 10.56 ± 0.15 d | \ |
| Trp | Bitter (−) | \ | 30.15 ± 0.44 a | \ | 21.51 ± 0.57 b | \ | 5.85 ± 0.09 c | \ | 29.47 ± 0.46 a | \ |
| Phe | Bitter (−) | 90 | 164.60 ± 3.94 a | 1.83 | 137.37 ± 0.23 b | 1.53 | 44.93 ± 0.46 c | 0.50 | 153.09 ± 8.62 a | 1.70 |
| Ile | Bitter (−) | 90 | 183.26 ± 4.11 a | 2.04 | 153.86 ± 3.19 b | 1.71 | 52.21 ± 0.67 c | 0.58 | 190.82 ± 0.61 a | 2.12 |
| Leu | Bitter (−) | 190 | 414.28 ± 4.35 b | 2.18 | 354.51 ± 4.84 c | 1.87 | 135.12 ± 3.15 d | 0.71 | 437.66 ± 2.51 a | 2.30 |
| Hyp | Bitter (\) | \ | 159.88 ± 0.69 a | \ | 130.65 ± 3.02 b | \ | 90.81 ± 2.89 c | \ | 118.79 ± 9.26 b | \ |
| Cit | Bitter (\) | \ | 7.61 ± 0.08 c | \ | 12.02 ± 0.35 b | \ | 2.68 ± 0.04 d | \ | 13.82 ± 0.28 a | \ |
| Asn | Odourless | \ | 78.78 ± 0.19 b | \ | 56.90 ± 1.04 c | \ | 17.90 ± 0.34 d | \ | 90.40 ± 2.39 a | \ |
| Gln | Odourless | \ | 3.45 ± 0.03 c | \ | 4.77 ± 0.11 b | \ | 1.71 ± 0.04 d | \ | 7.53 ± 0.10 a | \ |
| Gaba | Odourless | \ | 568.32 ± 3.04 b | \ | 553.98 ± 5.21 c | \ | 199.20 ± 0.59 d | \ | 619.59 ± 4.10 a | \ |
| Total Amino Acid Content | 4228.76 ± 37.14 b | 3767.25 ± 51.26 c | 1495.77 ± 22.63 d | 4328.70 ± 41.37 a | ||||||
| Categories | PK | Library | CAS | RT/min | Content/(μg/100g) | |||
|---|---|---|---|---|---|---|---|---|
| W1 | W2 | W3 | W4 | |||||
| Salts | A1 | Trimethyl borate | 000121-43-7 | 6.18 | 118.26 ± 2.63 d | 239.82 ± 5.46 a | 179.07 ± 9.51 b | 152.94 ± 2.91 c |
| A2 | Methyl butyrate | 000623-42-7 | 8.11 | 11.86 ± 2.13 c | 21.96 ± 1.46 b | \ | 26.94 ± 3.03 a | |
| A3 | Methyl 2-methylbutyrate | 000868-57-5 | 8.71 | 43.47 ± 1.79 a | \ | \ | 38.71 ± 2.01 b | |
| A4 | Methyl isovalerate | 000556-24-1 | 9.03 | 41.64 ± 1.72 b | 53.93 ± 2.68 a | \ | 33.49 ± 3.12 c | |
| A5 | Methyl caproate | 000106-70-7 | 14.13 | 89.44 ± 1.86 b | 213.30 ± 2.49 a | 10.29 ± 1.22 c | 93.04 ± 2.35b | |
| A6 | Methyl octanoate | 000111-11-5 | 20.30 | 70.34 ± 2.04 c | 234.75 ± 3.22 b | \ | 364.52 ± 3.69a | |
| A7 | Methyl n-caprate | 000110-42-9 | 25.74 | 25.50 ± 2.00 c | 51.34 ± 2.19 b | 7.57 ± 2.07 d | 261.30 ± 1.85a | |
| A8 | Dodecanoic acid, methylester | 000111-82-0 | 30.56 | 4.36 ± 1.83 c | 12.07 ± 1.34 b | 5.41 ± 1.94 c | 18.25 ± 2.15a | |
| A9 | Methyl myrist | 000124-10-7 | 34.88 | 27.54 ± 0.99 b | 39.53 ± 2.25 a | 18.08 ± 2.71 c | 37.64 ± 1.67a | |
| A10 | Methyl hexadecanoate | 000112-39-0 | 38.85 | 85.20 ± 1.93 b | 141.80 ± 1.49 a | 63.71 ± 1.50 d | 75.93 ± 3.28c | |
| A11 | 1,6-Hexanediol diacrylate | 013048-33-4 | 38.92 | 62.45 ± 2.52 b | 91.83 ± 1.44 a | 13.55 ± 2.00 d | 27.63 ± 1.25c | |
| A12 | Octadecanoic acid, methyl ester | 000112-61-8 | 42.51 | 22.16 ± 1.53 b | 31.57 ± 2.13 a | 17.43 ± 1.81 c | 21.82 ± 1.53b | |
| A13 | Methyl oleate | 000112-62-9 | 42.86 | 55.45 ± 1.96 c | 84.05 ± 2.43 a | 57.44 ± 2.15 c | 67.26 ± 0.84b | |
| A14 | Methyl linoleate | 000112-63-0 | 43.66 | 63.07 ± 2.61 b | 83.39 ± 2.71 a | 20.04 ± 3.44 d | 44.55 ± 1.99c | |
| A15 | Methyln-nonanoate | 001731-84-6 | 23.16 | \ | \ | \ | 13.53 ± 1.81 | |
| A16 | Ethyl caprate | 000110-38-3 | 25.78 | \ | \ | \ | 6.61 ± 1.92 | |
| A17 | 4-Decenoic acid, methylester, (4Z)- | 007367-83-1 | 27.05 | \ | \ | \ | 132.56 ± 2.37 | |
| Subtotal | 720.75 ± 5.59 c | 1308.35 ± 2.02 b | 392.61 ± 9.97 d | 1416.70 ± 0.64 a | ||||
| Aldehyde | B1 | Isovaleraldehyde | 000590-86-3 | 6.46 | 32.01 ± 1.77 c | 114.82 ± 1.50 a | 9.40 ± 1.07 d | 43.44 ± 1.79 b |
| B2 | Hexanal | 000066-25-1 | 11.05 | 10.31 ± 1.95 b | 15.59 ± 2.14 a | 4.44 ± 1.12 c | \ | |
| B3 | 1-Nonana | 000124-19-6 | 20.50 | 59.30 ± 1.73 b | 147.36 ± 2.16 a | 11.70 ± 1.35 c | 56.38 ± 1.00 b | |
| B4 | Phenylmethana | 000100-52-7 | 24.21 | 62.51 ± 2.16 c | 157.20 ± 2.41 a | 15.51 ± 1.27 d | 74.66 ± 0.74 b | |
| B5 | Phenylacetaldehyde | 000122-78-1 | 27.02 | 76.54 ± 0.54 b | 122.30 ± 1.75 a | 28.93 ± 1.70 c | \ | |
| B6 | trans,trans-2,4-Decadien-1-al | 025152-84-5 | 30.97 | 8.45 ± 2.18 b | 18.97 ± 1.66 a | \ | \ | |
| B7 | Tetradecanal | 000124-25-4 | 33.20 | 18.92 ± 1.57 b | 29.09 ± 1.47 a | \ | 16.63 ± 2.04 b | |
| B8 | Pentadecanal | 002765-11-9 | 35.34 | 26.31 ± 2.19 b | 44.21 ± 2.00 a | \ | 24.52 ± 2.16 b | |
| B9 | Hexadecanal | 000629-80-1 | 37.41 | 1029.03 ± 2.34 b | 1334.18 ± 2.37 a | 169.45 ± 1.84 d | 638.27 ± 1.84 c | |
| B10 | Heptadecanal | 1000376-70-0 | 39.35 | 68.20 ± 2.04 b | 72.42 ± 1.82 a | 14.37 ± 1.25 d | 46.29 ± 2.06 c | |
| B11 | Octadecanal | 000638-66-4 | 41.23 | 195.35 ± 2.03 a | 143.28 ± 1.77 b | 25.42 ± 2.09 d | 98.52 ± 2.35 c | |
| B12 | 13-Octadecenal, (13Z)- | 058594-45-9 | 41.66 | 142.35 ± 1.82 a | 112.64 ± 1.96 b | 17.48 ± 1.92 d | 75.19 ± 2.05 c | |
| B13 | 2-Methylbutyraldehyde | 000096-17-3 | 6.38 | \ | 16.66 ± 1.50 a | \ | 12.56 ± 2.07 b | |
| B14 | 2-Undecenal | 002463-77-6 | 29.61 | \ | \ | 27.33 ± 1.87 | \ | |
| Subtotal | 1729.29 ± 10.07 b 2328.71 ± 3.92 a | 324.03 ± 10.79 d | 1086.46 ± 4.30 c | |||||
| Hydrocarbons | C1 | Valencene | 004630-07-3 | 28.07 | 8.35 ± 2.25 ab | 5.54 ± 2.07 b | \ | 9.27 ± 2.05a |
| C2 | trans-Caryophyllene | 000087-44-5 | 28.90 | 21.90 ± 1.47 c | 39.36 ± 2.15 a | 7.39 ± 1.08 d | 29.25 ± 2.01b | |
| C3 | alpha-himachalene | 003853-83-6 | 28.98 | 27.63 ± 1.90 c | 54.13 ± 1.00 a | 11.94 ± 1.48 d | 35.92 ± 2.54b | |
| C4 | delta-Cadinene | 000483-76-1 | 29.60 | 53.59 ± 1.80 c | 124.63 ± 0.98 a | \ | 56.91 ± 1.74b | |
| C5 | germacrene d | 023986-74-5 | 29.70 | 13.50 ± 1.96 c | 25.53 ± 1.97 a | 6.41 ± 0.80 d | 17.25 ± 2.21 b | |
| C6 | α-curcumene | 000644-30-4 | 29.93 | 22.45 ± 2.03 c | 48.73 ± 2.53 a | 11.54 ± 1.00 d | 32.57 ± 0.99 b | |
| C7 | Cuparene | 016982-00-6 | 31.13 | 14.32 ± 1.02 b | 25.86 ± 2.02 a | 6.51 ± 0.92 c | 25.51 ± 1.79 a | |
| C8 | Calamenene | 000483-77-2 | 31.29 | 12.44 ± 2.07 c | 30.58 ± 2.13 a | \ | 21.73 ± 1.33 b | |
| C9 | D-Limonene | 005989-27-5 | 14.37 | \ | 22.39 ± 2.04 | \ | \ | |
| C10 | Pentadecane | 000629-62-9 | 23.57 | 20.56 ± 0.96 b | 34.51 ± 1.04 a | 7.84 ± 1.50 c | 18.65 ± 2.35 b | |
| C11 | n-Hexadecane | 000544-76-3 | 26.07 | 32.38 ± 2.12 b | 49.43 ± 1.94 a | \ | 19.26 ± 1.02 c | |
| C12 | n-Heptadecane | 000629-78-7 | 28.41 | 14.57 ± 1.01 b | 23.59 ± 2.01 a | \ | 9.32 ± 1.56 c | |
| C13 | Hexane | 000110-54-3 | 3.69 | \ | 45.68 ± 1.79 b | 88.89 ± 1.66 a | 23.53 ± 1.87 c | |
| C14 | Heptane | 000142-82-5 | 4.00 | \ | 41.91 ± 1.53 a | 37.26 ± 0.86 b | 16.35 ± 1.05 c | |
| C15 | Cyclohexane | 000110-82-7 | 4.09 | \ | 78.6 ± 2.04 a | 64.43 ± 2.06 b | 38.59 ± 2.08 c | |
| Subtotal | 241.67 ± 2.80 c | 650.50 ± 3.49 a | 242.21 ± 2.13 c | 354.10 ± 9.46 b | ||||
| Alcohol | D1 | Mushroom alcohol | 003391-86-4 | 21.96 | 7.58 ± 1.79 b | 28.03 ± 1.48 a | \ | 10.05 ± 1.86 b |
| D2 | Dodecyl alcohol | 000112-53-8 | 33.92 | 16.48 ± 0.89 b | 37.23 ± 1.86 a | 4.42 ± 0.99 c | \ | |
| D3 | 1-Octanol | 000111-87-5 | 24.76 | \ | 30.31 ± 1.03 | \ | \ | |
| D4 | 2-Phenylethanol | 000060-12-8 | 32.90 | \ | 14.35 ± 0.85 a | \ | 7.39 ± 2.18 b | |
| D5 | Dodecyl alcohol | 000112-53-8 | 33.92 | \ | \ | 33.51 ± 1.88 a | 9.30 ± 1.20 b | |
| Subtotal | 24.06 ± 2.64 d | 109.92 ± 0.38 a | 37.92 ± 0.89 c | 32.74 ± 3.57 b | ||||
| Else | E1 | Butylated hydroxytoluene | 000128-37-0 | 32.85 | 8.10 ± 1.68 b | 11.47 ± 0.96 a | \ | 5.56 ± 1.93 b |
| E2 | Phenol | 000108-95-2 | 34.73 | 12.37 ± 2.03 | \ | \ | \ | |
| E3 | 2,4-Di-tert-butylphenol | 000096-76-4 | 40.28 | \ | 8.41 ± 2.02 | \ | \ | |
| E4 | Methyl tridecyl ketone | 002345-28-0 | 35.18 | 7.69 ± 1.52 b | \ | \ | 10.19 ± 1.40 a | |
| E5 | 2,6-Dimethyl pyrazine | 000108-50-9 | 18.94 | 19.52 ± 1.95 | \ | \ | \ | |
| E6 | Estragole | 000140-67-0 | 31.29 | \ | \ | 10.45 ± 0.84 | \ | |
| E7 | Dimethyl sulfoxide | 000067-68-5 | 26.07 | \ | \ | 15.40 ± 2.07 | \ | |
| Subtotal | 47.67 ± 7.08 a | 19.88 ± 1.07 bc | 25.85 ± 1.34 b | 15.75 ± 0.57 c | ||||
| Aggregate | 59 | 2763.44 ± 3.88 c | 4417.35 ± 3.60 a | 1022.62 ± 20.76 d 2944.08 ± 9.71 b | ||||
| Transducers | Responsive Substance | Category Substances |
|---|---|---|
| S1 | Alkanes, fumes | Propane, natural gas, fumes |
| S2 | Alcohols, aldehydes, short-chain alkanes | Alcohol, fumes, isobutane, formaldehyde |
| S3 | Ozone (O3) | \ |
| S4 | Sulphide | Hydrogen sulphide |
| S5 | Organic amine | Ammonia, methylamine, ethanolamine |
| S6 | Organic gases, benzophenones, alcohols, and aldehydes; aromatic compounds | Toluene, acetone, ethanol, hydrogen, and other organic vapours |
| S7 | Short-chain burnt hydrocarbons | Methane, natural gas, biogas |
| S8 | Aromatic compounds, alcohols, and aldehydes | Toluene, formaldehyde, benzene, alcohol, acetone |
| S9 | Hydrogen-containing gas | Hydrogen (gas) |
| S10 | Flammable gases | Methane CH4 |
| W1 | W2 | W3 | W4 | |
|---|---|---|---|---|
| chao1 | 262.72 | 232.83 | 405.41 | 253.079 |
| observed_features | 231 | 200.33 | 342 | 214.33 |
| shannon_entropy | 3.71 | 4.09 | 4.61 | 3.69 |
| Simpson | 0.78 | 0.84 | 0.89 | 0.79 |
| Raw Base (GB) | 6.64 | 6.09 | 7.19 | 6.33 |
| Clean Reads | 197,130 | 170,440 | 255,527 | 189,746 |
| Clean Q20 (%) | 99.42 | 99.36 | 99.29 | 99.40 |
| Clean Q30 (%) | 97.91 | 97.78 | 97.46 | 97.86 |
| Clean GC (%) | 42.33 | 42.67 | 43.00 | 41.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Li, S.; Wen, Y.; Yang, J.; Wang, P.; Wang, H.; Cui, Y.; Wu, W.; Li, L.; Liu, Z. Correlation between the Characteristic Flavour and Microbial Community of Xuanwei Ham after Ripening. Fermentation 2024, 10, 392. https://doi.org/10.3390/fermentation10080392
Li G, Li S, Wen Y, Yang J, Wang P, Wang H, Cui Y, Wu W, Li L, Liu Z. Correlation between the Characteristic Flavour and Microbial Community of Xuanwei Ham after Ripening. Fermentation. 2024; 10(8):392. https://doi.org/10.3390/fermentation10080392
Chicago/Turabian StyleLi, Guipeng, Simin Li, Yiling Wen, Jing Yang, Ping Wang, Huaiyao Wang, Yawen Cui, Wenliang Wu, Liang Li, and Zhendong Liu. 2024. "Correlation between the Characteristic Flavour and Microbial Community of Xuanwei Ham after Ripening" Fermentation 10, no. 8: 392. https://doi.org/10.3390/fermentation10080392