
Harnessing Fermentation by Bacillus and Lactic Acid Bacteria for Enhanced Texture, Flavor, and Nutritional Value in Plant-Based Matrices

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Base Preparation and Fermentation
2.3. Analysis of Volatile Organic Compounds
2.4. Raffinose-Family-Type Alpha-Galactooligosaccharide Analysis
2.5. Carotenoid Production
2.6. Statistical Analysis
3. Results and Discussion
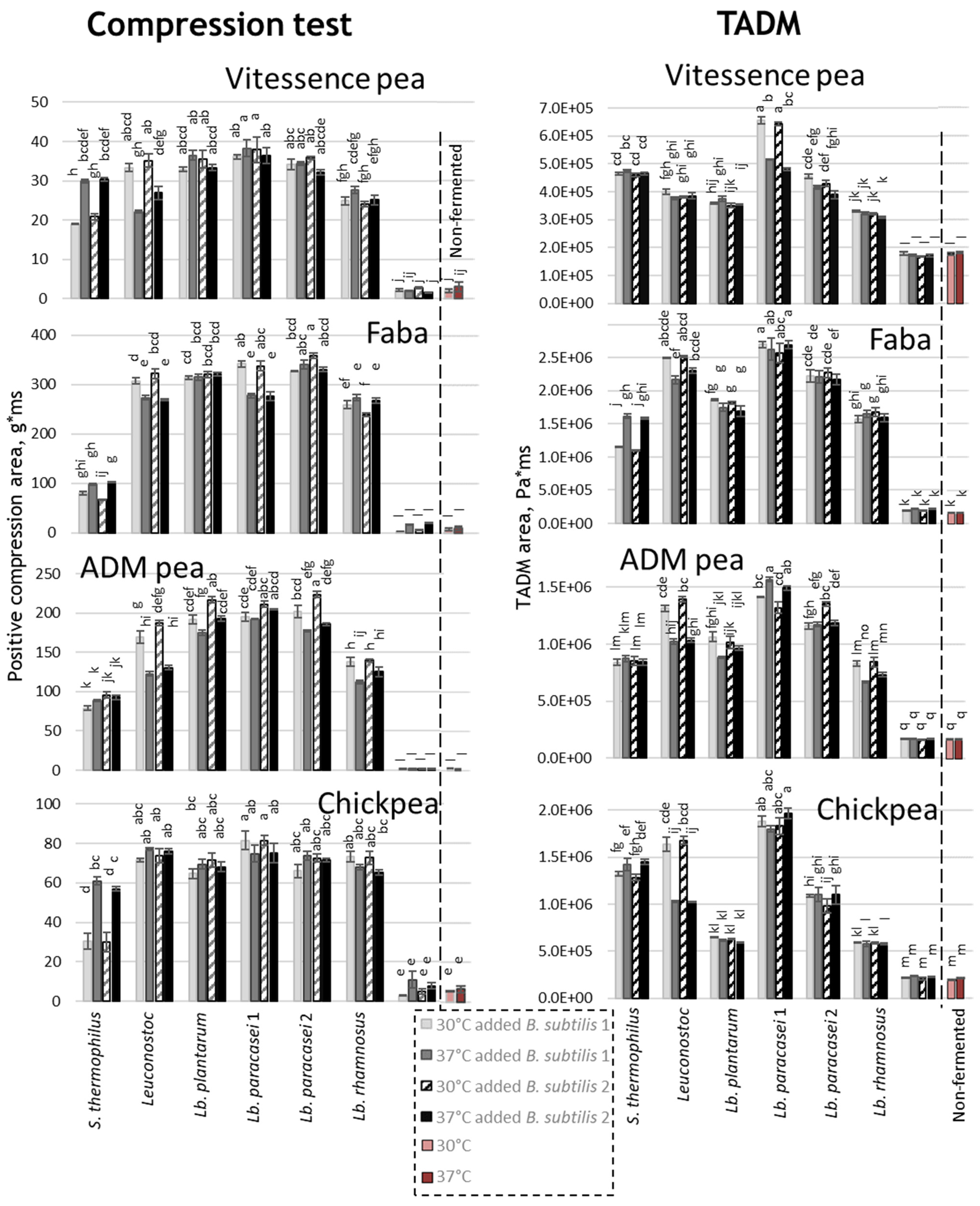
3.1. Texture Formation
3.2. Reduction of Off-Flavors
3.3. Formation of Desirable Dairy Note Compounds
3.4. Ethanol and Ester Formation
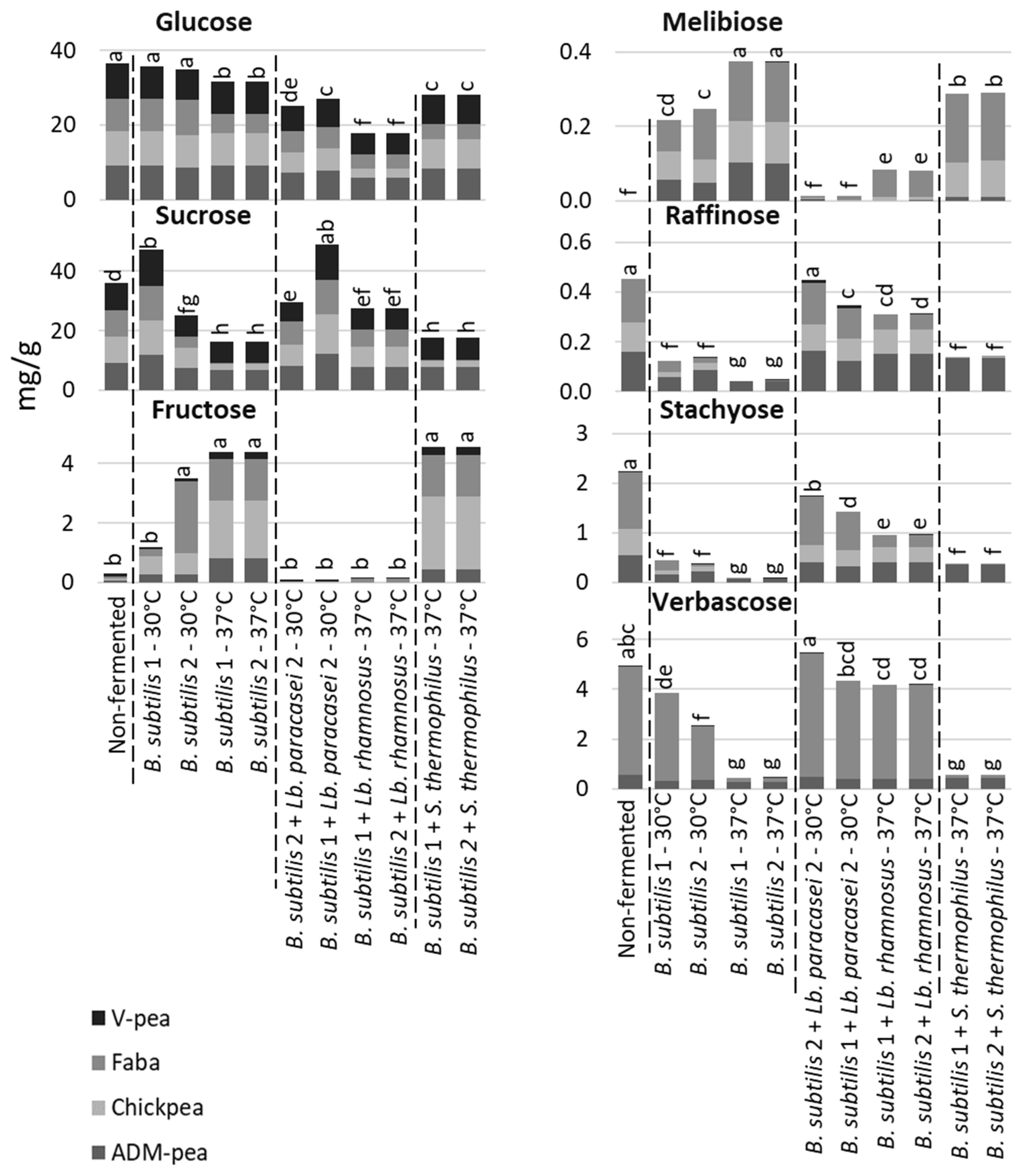
3.5. Degradation of Raffinose Family Oligosaccharides
3.6. Pigment Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McClements, D.J. Meat Less: The Next Food Revolution; Springer: Berlin/Heidelberg, Germany, 2023; Volume 30. [Google Scholar]
- Nolden, A.A.; Forde, C.G. The Nutritional Quality of Plant-Based Foods. Sustainability 2023, 15, 3324. [Google Scholar] [CrossRef]
- Poulsen, V.K.; Moghadam, E.G.; Kračun, S.K.; Svendsen, B.A.; Nielsen, W.M.; Oregaard, G.; Krarup, A. Versatile Lactococcus lactis strains improve texture in both fermented milk and soybean matrices. FEMS Microbiol. Lett. 2022, 369, fnac117. [Google Scholar] [CrossRef] [PubMed]
- Tangyu, M.; Fritz, M.; Tan, J.P.; Ye, L.; Bolten, C.J.; Bogicevic, B.; Wittmann, C. Flavour by design: Food-grade lactic acid bacteria improve the volatile aroma spectrum of oat milk, sunflower seed milk, pea milk, and faba milk towards improved flavour and sensory perception. Microb. Cell Fact. 2023, 22, 133. [Google Scholar] [CrossRef] [PubMed]
- Masiá, C.; Fernández-Varela, R.; Poulsen, V.K.; Jensen, P.E.; Sørensen, K.I. Composition of bacterial blends for fermentation-induced pea protein emulsion gels using multi-property screening of lactic acid bacteria. Food Biosci. 2023, 56, 103333. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed]
- Beck, F.; Pedersen, N.R.; Nielsen, D.S. Fermented Rapeseed and Soybean Alone and in Combination with Macro Algae Inhibit Human and Pig Pathogenic Bacteria In Vitro. Microorganisms 2024, 12, 891. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.R.; Dobson, R.C.J.; Morris, V.K.; Moggré, G.J. Fermentation of plant-based dairy alternatives by lactic acid bacteria. Microb. Biotechnol. 2022, 15, 1404–1421. [Google Scholar] [CrossRef] [PubMed]
- Engels, W.; Siu, J.; van Schalkwijk, S.; Wesselink, W.; Jacobs, S.; Bachmann, H. Metabolic Conversions by Lactic Acid Bacteria during Plant Protein Fermentations. Foods 2022, 11, 1005. [Google Scholar] [CrossRef]
- Heravi, K.M.; Watzlawick, H.; Altenbuchner, J. The melredca operon encodes a utilization system for the raffinose family of oligosaccharides in bacillus subtilis. J. Bacteriol. 2019, 201, e00109-19. [Google Scholar] [CrossRef]
- Elango, D.; Rajendran, K.; Van der Laan, L.; Sebastiar, S.; Raigne, J.; Thaiparambil, N.A.; El Haddad, N.; Raja, B.; Wang, W.; Ferela, A.; et al. Raffinose Family Oligosaccharides: Friend or Foe for Human and Plant Health? Front. Plant Sci. 2022, 13, 829118. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, S.; Bain, J.R.; Muehlbauer, M.J.; Provenza, F.D.; Kronberg, S.L.; Pieper, C.F.; Huffman, K.M. A metabolomics comparison of plant-based meat and grass-fed meat indicates large nutritional differences despite comparable Nutrition Facts panels. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Ahmad, M.I.; Farooq, S.; Alhamoud, Y.; Li, C.; Zhang, H. Soy Leghemoglobin: A review of its structure, production, safety aspects, and food applications. Trends Food Sci. Technol. 2023, 141, 104199. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Kauser-Ul-alam, M.; Toba, Y.; Hioki, S.; Hayakawa, T.; Kumura, H.; Wakamatsu, J.I. Lactococcus lactis subsp. cremoris Produces Zinc Protoporphyrin IX Both Aerobically and Anaerobically and Improves the Bright Red Color of Fermented Meat Products. Foods 2020, 9, 1583. [Google Scholar] [CrossRef]
- Solopova, A.; van Tilburg, A.Y.; Foito, A.; Allwood, J.W.; Stewart, D.; Kulakauskas, S.; Kuipers, O.P. Engineering Lactococcus lactis for the production of unusual anthocyanins using tea as substrate. Metab. Eng. 2019, 54, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Parajuli, P.; Koffas, M.A.G.; Sohng, J.K. Microbial production of natural and non-natural flavonoids: Pathway engineering, directed evolution and systems/synthetic biology. Biotechnol. Adv. 2016, 34, 634–662. [Google Scholar] [CrossRef]
- Zha, J.; Wu, X.; Koffas, M.A. Making brilliant colors by microorganisms. Curr. Opin. Biotechnol. 2020, 61, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, X.; Zeng, W.; Xu, S.; Li, D.; Yu, S.; Zhou, J. De novo biosynthesis of carminic acid in Saccharomyces cerevisiae. Metab. Eng. 2023, 76, 50–62. [Google Scholar] [CrossRef]
- Frandsen, R.J.N.; Khorsand-Jamal, P.; Kongstad, K.T.; Nafisi, M.; Kannangara, R.M.; Staerk, D.; Okkels, F.T.; Binderup, K.; Madsen, B.; Møller, B.L. Heterologous production of the widely used natural food colorant carminic acid in Aspergillus nidulans. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Sandmann, G.; Pollmann, H.; Gassel, S.; Breitenbach, J. Xanthophyllomyces dendrorhous, a Versatile Platform for the Production of Carotenoids and Other Acetyl-CoA-Derived Compounds. Adv. Exp. Med. Biol. 2021, 1261, 137–151. [Google Scholar] [CrossRef]
- Buzzini, P.; Innocenti, M.; Turchetti, B.; Libkind, D.; Van Broock, M.; Mulinacci, N. Carotenoid profiles of yeasts belonging to the genera Rhodotorula, Rhodosporidium, Sporobolomyces, and Sporidiobolus. Can. J. Microbiol. 2007, 53, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Legein, M.; Wittouck, S.; Lebeer, S. Latilactobacillus fragifolii sp. nov., isolated from leaves of a strawberry plant (Fragaria x ananassa). Int. J. Syst. Evol. Microbiol. 2022, 72, 005193. [Google Scholar] [CrossRef]
- Kuzina, V.; Cerdá-Olmedo, E. Ubiquinone and carotene production in the Mucorales Blakeslea and Phycomyces. Appl. Microbiol. Biotechnol. 2007, 76, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Cerdá-Olmedo, E. Phycomyces and the biology of light and color. FEMS Microbiol. Rev. 2001, 25, 503–512. [Google Scholar] [CrossRef]
- Ruiz-Hidalgo, M.J.; López-Matas, M.A.; Velayos, A.; Fraser, P.D.; Bramley, P.M.; Eslava, A.P. Carotenoid Mutants of Mucor circinelloides. Bot. Acta 1995, 108, 396–400. [Google Scholar] [CrossRef]
- Coleman, B.; Van Poucke, C.; De Witte, B.; Ruttens, A.; Moerdijk-Poortvliet, T.C.W.; Latsos, C.; De Reu, K.; Blommaert, L.; Duquenne, B.; Timmermans, K.; et al. Potential of microalgae as flavoring agents for plant-based seafood alternatives. Future Foods 2022, 5, 100139. [Google Scholar] [CrossRef]
- Kazir, M.; Livney, Y.D. Plant-based seafood analogs. Molecules 2021, 26, 1559. [Google Scholar] [CrossRef]
- García-Vaquero, M.; Brunton, N.; Lafarga, T. Microalgae as a source of pigments for food applications. In Cultured Microalgae for the Food Industry: Current and Potential Applications; Academic Press: New York, NY, USA, 2021. [Google Scholar] [CrossRef]
- Balasubramaniam, V.; Chelyn, L.J.; Vimala, S.; Fairulnizal, M.N.M.; Brownlee, I.A.; Amin, I. Carotenoid composition and antioxidant potential of Eucheuma denticulatum, Sargassum polycystum and Caulerpa lentillifera. Heliyon 2020, 6, e04654. [Google Scholar] [CrossRef]
- Rocchi, R.; Zwinkels, J.; Kooijman, M.; Garre, A.; Smid, E.J. Development of novel natto using legumes produced in Europe. Heliyon 2024, 10, e26849. [Google Scholar] [CrossRef]
- Bjerre, K.; Cantor, M.D.; Janzen, T.; Derkx, P. Fermented Milk Inoculated with both Lactic Acid Bacteria (LAB) and Bacillus. U.S. Patent US11684073B2, 27 June 2017. [Google Scholar]
- Poulsen, V.K.; Derkx, P.; Oregaard, G. High-throughput screening for texturing Lactococcus strains. FEMS Microbiol. Lett. 2019, 366, fnz001. [Google Scholar] [CrossRef]
- Lara-Abia, S.; Lobo-Rodrigo, G.; Welti-Chanes, J.; Cano, M.P. Carotenoid and carotenoid ester profile and their deposition in plastids in fruits of new papaya (Carica papaya L.) varieties from the canary islands. Foods 2021, 10, 434. [Google Scholar] [CrossRef]
- Saffarionpour, S. Off-Flavors in Pulses and Grain Legumes and Processing Approaches for Controlling Flavor-Plant Protein Interaction: Application Prospects in Plant-Based Alternative Foods. Food Bioprocess Technol. 2023, 17, 1141–1182. [Google Scholar] [CrossRef]
- Fischer, E.; Cachon, R.; Cayot, N. Impact of Ageing on Pea Protein Volatile Compounds and Correlation with Odor. Molecules 2022, 27, 852. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tuccillo, F.; Lampi, A.M.; Knaapila, A.; Pulkkinen, M.; Kariluoto, S.; Coda, R.; Edelmann, M.; Jouppila, K.; Sandell, M.; et al. Flavor challenges in extruded plant-based meat alternatives: A review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2898–2929. [Google Scholar] [CrossRef] [PubMed]
- El Youssef, C.; Bonnarme, P.; Fraud, S.; Péron, A.C.; Helinck, S.; Landaud, S. Sensory improvement of a pea protein-based product using microbial co-cultures of lactic acid bacteria and yeasts. Foods 2020, 9, 349. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Lan, Y.; Rao, J.; Chen, B. HS-SPME-GC-MS/olfactometry combined with chemometrics to assess the impact of germination on flavor attributes of chickpea, lentil, and yellow pea flours. Food Chem. 2019, 280, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Macciola, V.; Candela, G.; De Leonardis, A. Rapid gas-chromatographic method for the determination of diacetyl in milk, fermented milk and butter. Food Control 2008, 19, 873–878. [Google Scholar] [CrossRef]
- Zhao, M.; Li, H.; Zhang, D.; Li, J.; Wen, R.; Ma, H.; Zou, T.; Hou, Y.; Song, H. Variation of Aroma Components of Pasteurized Yogurt with Different Process Combination before and after Aging by DHS/GC-O-MS. Molecules 2023, 28, 1975. [Google Scholar] [CrossRef]
- Pedersen, T.B.; Ristagno, D.; McSweeney, P.L.H.; Vogensen, F.K.; Ardö, Y. Potential impact on cheese flavour of heterofermentative bacteria from starter cultures. Int. Dairy J. 2013, 33, 112–119. [Google Scholar] [CrossRef]
- Rajendran, S.; Silcock, P.; Bremer, P. Flavour Volatiles of Fermented Vegetable and Fruit Substrates: A Review. Molecules 2023, 28, 3236. [Google Scholar] [CrossRef]
- Boulay, M.; Al Haddad, M.; Rul, F. Streptococcus thermophilus growth in soya milk: Sucrose consumption, nitrogen metabolism, soya protein hydrolysis and role of the cell-wall protease PrtS. Int. J. Food Microbiol. 2020, 335, 108903. [Google Scholar] [CrossRef]
- Donkor, O.N.; Henriksson, A.; Vasiljevic, T.; Shah, N.P. α-Galactosidase and proteolytic activities of selected probiotic and dairy cultures in fermented soymilk. Food Chem. 2007, 104, 10–20. [Google Scholar] [CrossRef]
- Wang, Y.C.; Yu, R.C.; Yang, H.Y.; Chou, C.C. Sugar and acid contents in soymilk fermented with lactic acid bacteria alone or simultaneously with bifidobacterial. Food Microbiol. 2003, 20, 333–338. [Google Scholar] [CrossRef]
- Garro, M.S.; De Valdez, G.F.; Oliver, G.; De Giori, G.S. Growth characteristics and fermentation products of Streptococcus salivarius subsp. thermophilus, Lactobacillus casei and L. fermentum in soymilk. Eur. Food Res. Technol. 1998, 206, 72–75. [Google Scholar] [CrossRef]
- Kim, M.; Jung, D.H.; Seo, D.H.; Chung, W.H.; Seo, M.J. Genome analysis of Lactobacillus plantarum subsp. plantarum KCCP11226 reveals a well-conserved C30 carotenoid biosynthetic pathway. 3 Biotech 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Tanger, C.; Engel, J.; Kulozik, U. Influence of extraction conditions on the conformational alteration of pea protein extracted from pea flour. Food Hydrocoll. 2020, 107, 105949. [Google Scholar] [CrossRef]
- Emkani, M.; Oliete, B.; Saurel, R. Pea protein extraction assisted by lactic fermentation: Impact on protein profile and thermal properties. Foods 2021, 10, 549. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Varela, R.; Hansen, A.H.; Svendsen, B.A.; Moghadam, E.G.; Bas, A.; Kračun, S.K.; Harlé, O.; Poulsen, V.K. Harnessing Fermentation by Bacillus and Lactic Acid Bacteria for Enhanced Texture, Flavor, and Nutritional Value in Plant-Based Matrices. Fermentation 2024, 10, 411. https://doi.org/10.3390/fermentation10080411
Fernández-Varela R, Hansen AH, Svendsen BA, Moghadam EG, Bas A, Kračun SK, Harlé O, Poulsen VK. Harnessing Fermentation by Bacillus and Lactic Acid Bacteria for Enhanced Texture, Flavor, and Nutritional Value in Plant-Based Matrices. Fermentation. 2024; 10(8):411. https://doi.org/10.3390/fermentation10080411
Chicago/Turabian StyleFernández-Varela, Raquel, Anders Holmgaard Hansen, Birgit Albrecht Svendsen, Elahe Ghanei Moghadam, Arzu Bas, Stjepan Krešimir Kračun, Olivier Harlé, and Vera Kuzina Poulsen. 2024. "Harnessing Fermentation by Bacillus and Lactic Acid Bacteria for Enhanced Texture, Flavor, and Nutritional Value in Plant-Based Matrices" Fermentation 10, no. 8: 411. https://doi.org/10.3390/fermentation10080411
APA StyleFernández-Varela, R., Hansen, A. H., Svendsen, B. A., Moghadam, E. G., Bas, A., Kračun, S. K., Harlé, O., & Poulsen, V. K. (2024). Harnessing Fermentation by Bacillus and Lactic Acid Bacteria for Enhanced Texture, Flavor, and Nutritional Value in Plant-Based Matrices. Fermentation, 10(8), 411. https://doi.org/10.3390/fermentation10080411