
Enhancement of Active Substances in Astragali Radix Broth with Lactic Acid Bacteria Fermentation and the Promotion Role of Chlorella Growth Factor

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains and Reagents
2.2. Preparation of Fermentation Medium and CGF Extract
- (1)
- Fermentation medium: Astragali Radix was crushed to powder and sieved through an 80-mesh sieve. A total of 2 g of dried Astragali Radix powder was taken and mixed in 20 mL of distilled water and then sterilized at 121 °C for 25 min and cooled for later use.
- (2)
- Fermentation lactic acid bacteria biomass: a total of 1 mL was taken from each of the aforementioned six preserved lactic acid bacteria biomass (with a colony count of 2.0 × 106 CFU/mL for each) and were inoculated into 100 mL of NB medium (beef extract 0.3 g, peptone 1.0 g, sodium chloride 0.5 g, distilled water 100 mL, pH 7.2–7.5). Then, the solution was incubated at 37 °C for 24 h to obtain the fermentation lactic acid bacteria biomass. (The total colony count of lactic acid bacteria in the solution was counted as 2.3 × 107 CFU/mL).
- (3)
- Chlorella Growth Factor (CGF) extract: 50 mL fresh Chlorella sp. culture medium (counting 3.5 × 107/mL microalgal cells) was subjected to centrifugation at 4000 rpm for 10 min. The resulting precipitate was collected and washed three times with sterile water [19]. Subsequently, it was dissolved in another 50 mL of sterile water and heated in an autoclaved cooker at 121 °C for 15 min to obtain the CGF extract [20].
2.3. Fermentation of Astragali Radix and Determination of Active Compounds
2.3.1. Fermentation of Astragali Radix
2.3.2. Treatment of Fermentation Broth and Determination of Active Compounds
- (1)
- Determination of total polysaccharide content
- (2)
- Determination of total flavonoid content
- (3)
- Determination of total saponin content
2.4. Analysis of Microbial Community Structure
2.5. Non-Targeted Metabolomics Analysis
2.6. Data Processing
3. Results and Discussion
3.1. Effect of Fermentation Conditions on the Content of Active Substances in Astragali Radix Fermentation Broth
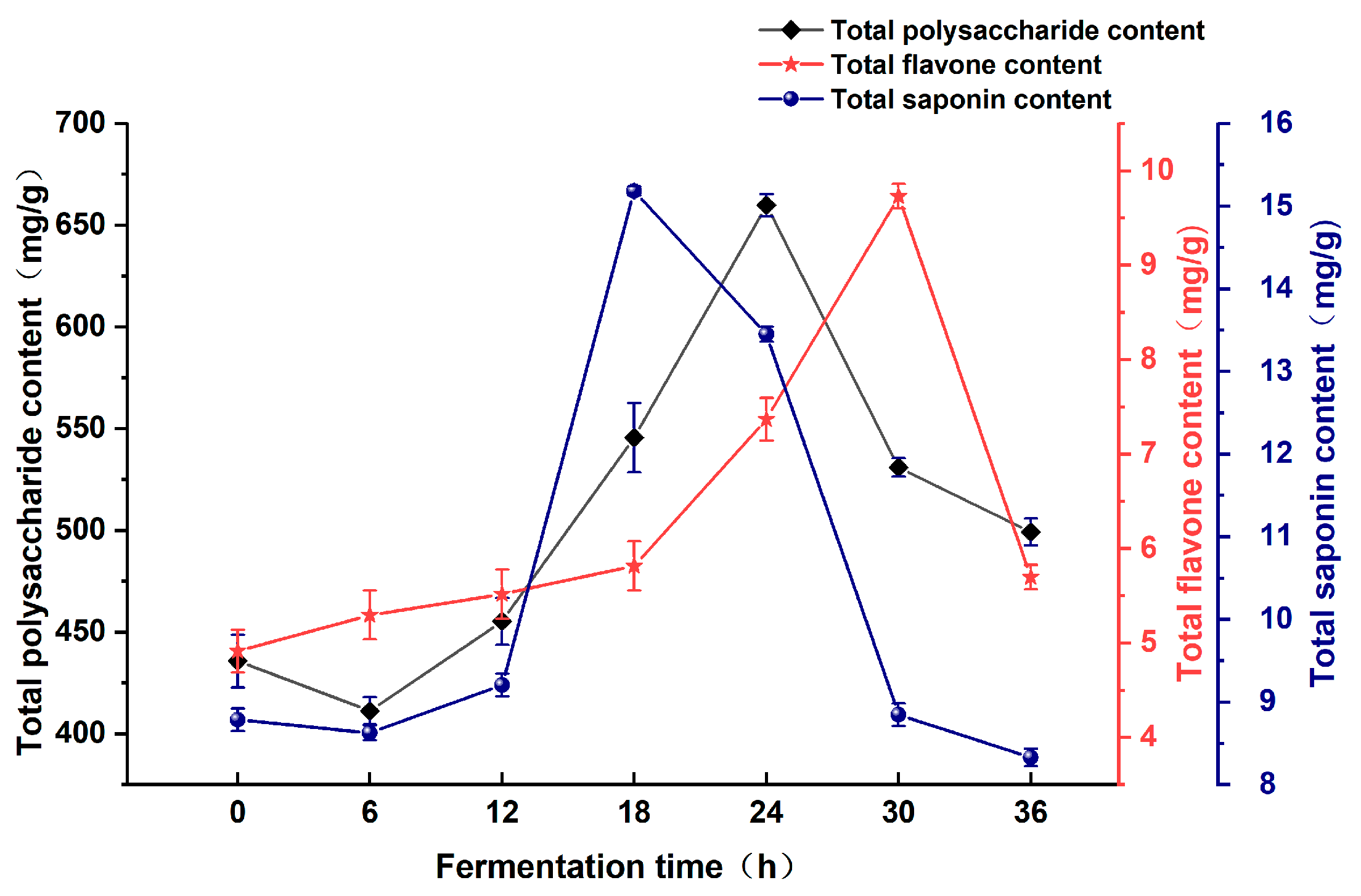
3.1.1. Effect of Fermentation Time
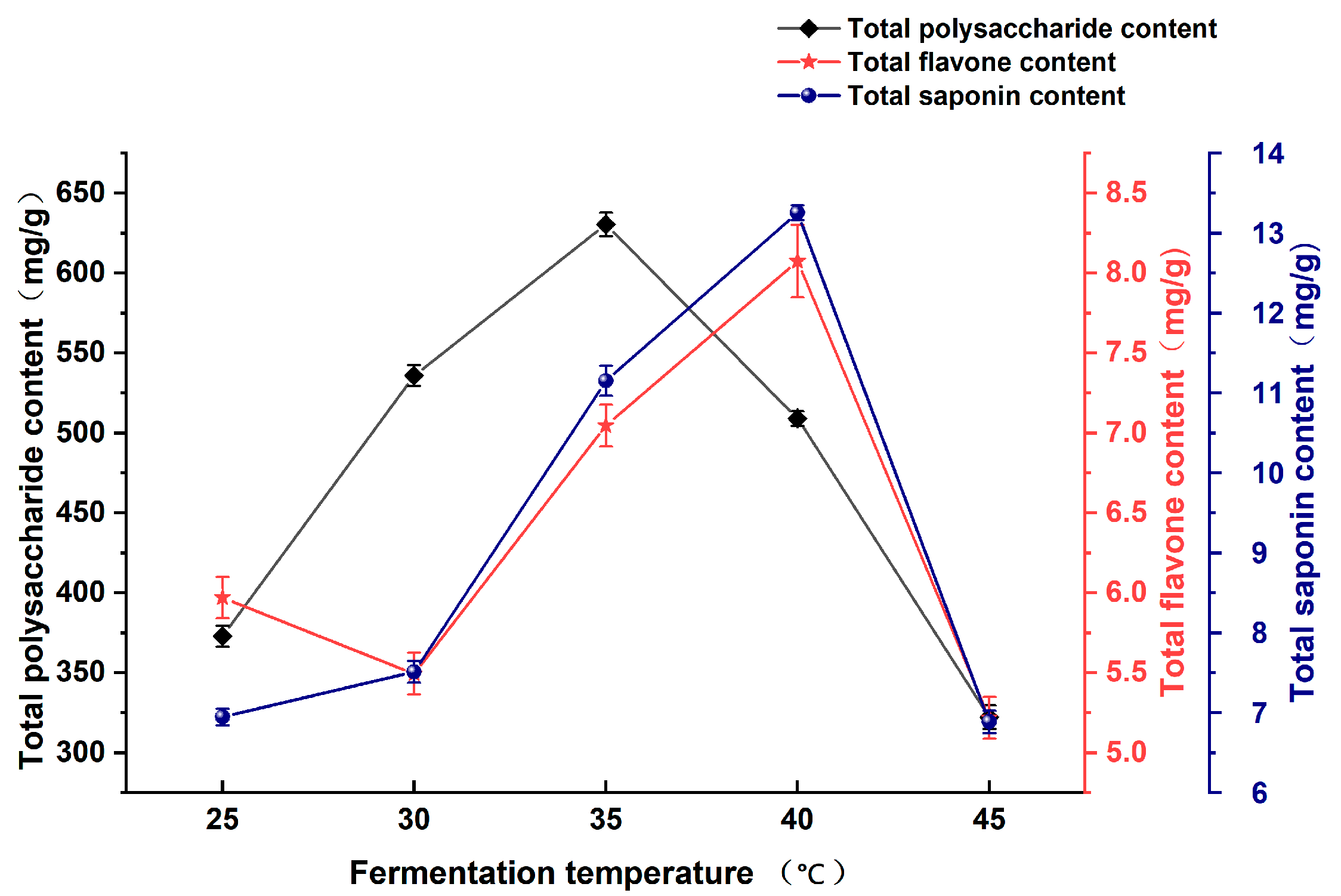
3.1.2. Effect of Fermentation Temperature
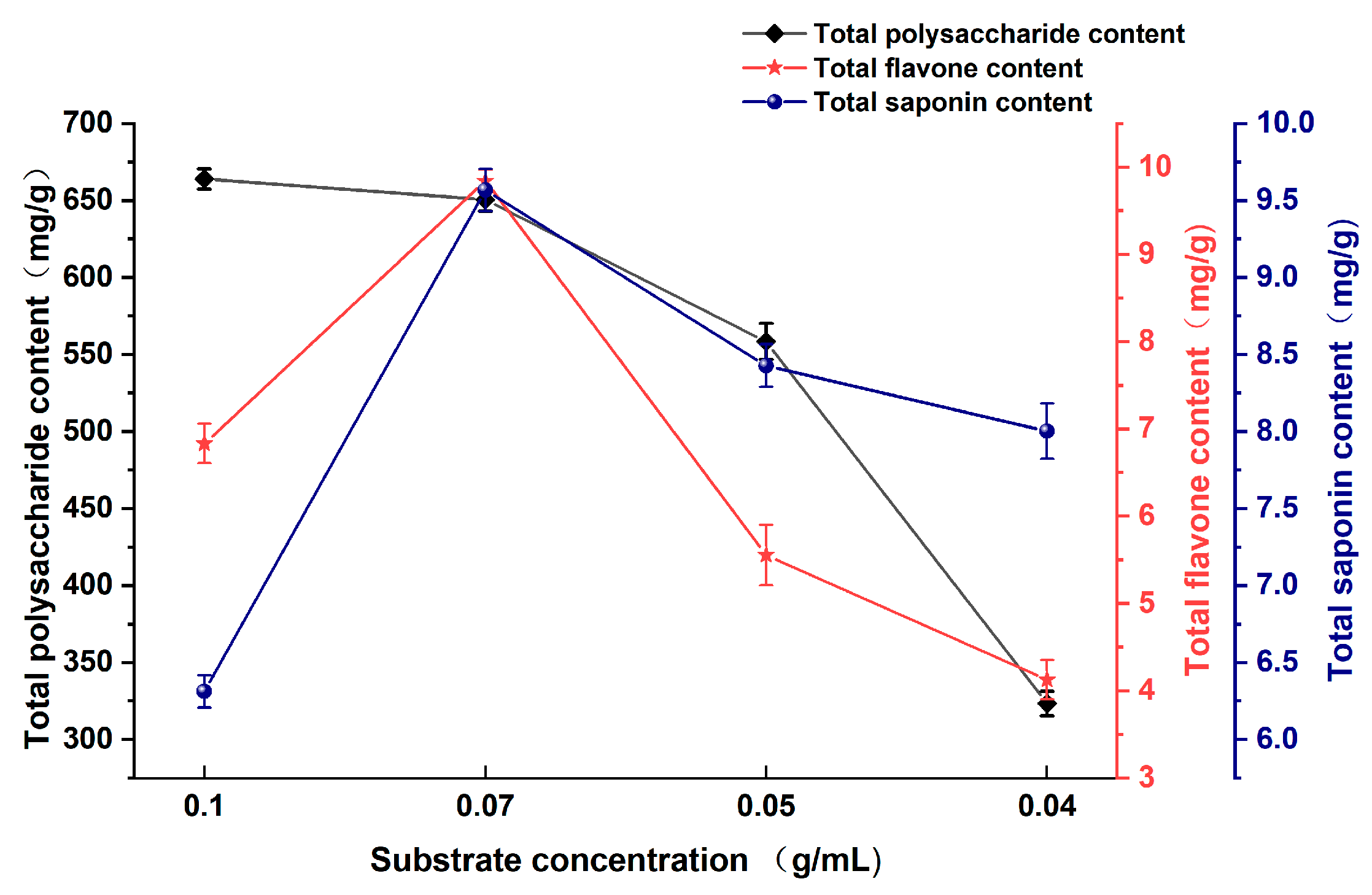
3.1.3. Effect of Substrate Concentration
3.1.4. Effect of pH
3.1.5. Effect of Bacteria Inoculum Dosage
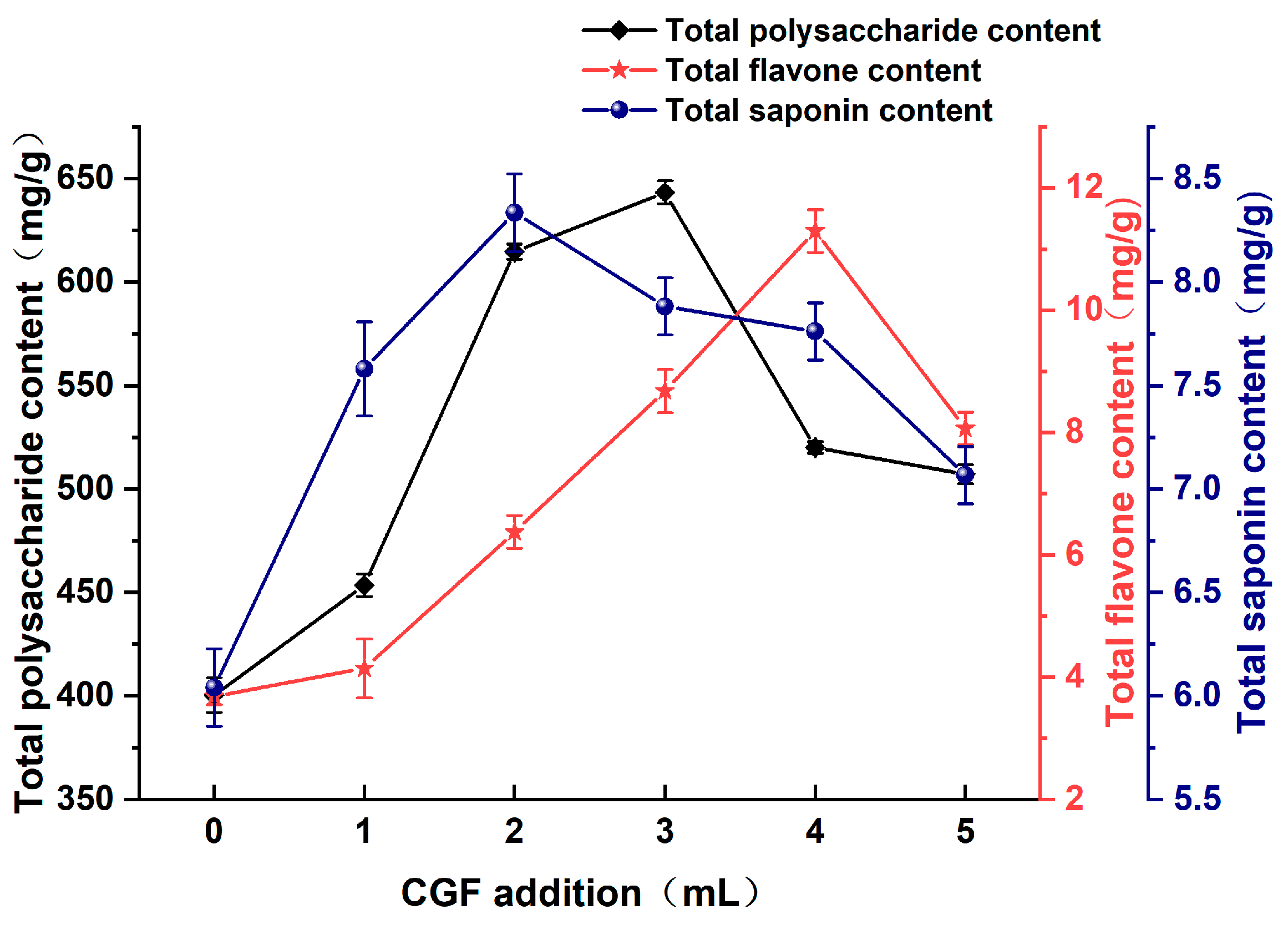
3.1.6. Effect of CGF Addition
3.1.7. Optimum Fermentation Conditions and Effects
3.2. Changes in Microbiological Composition before and after Fermentation
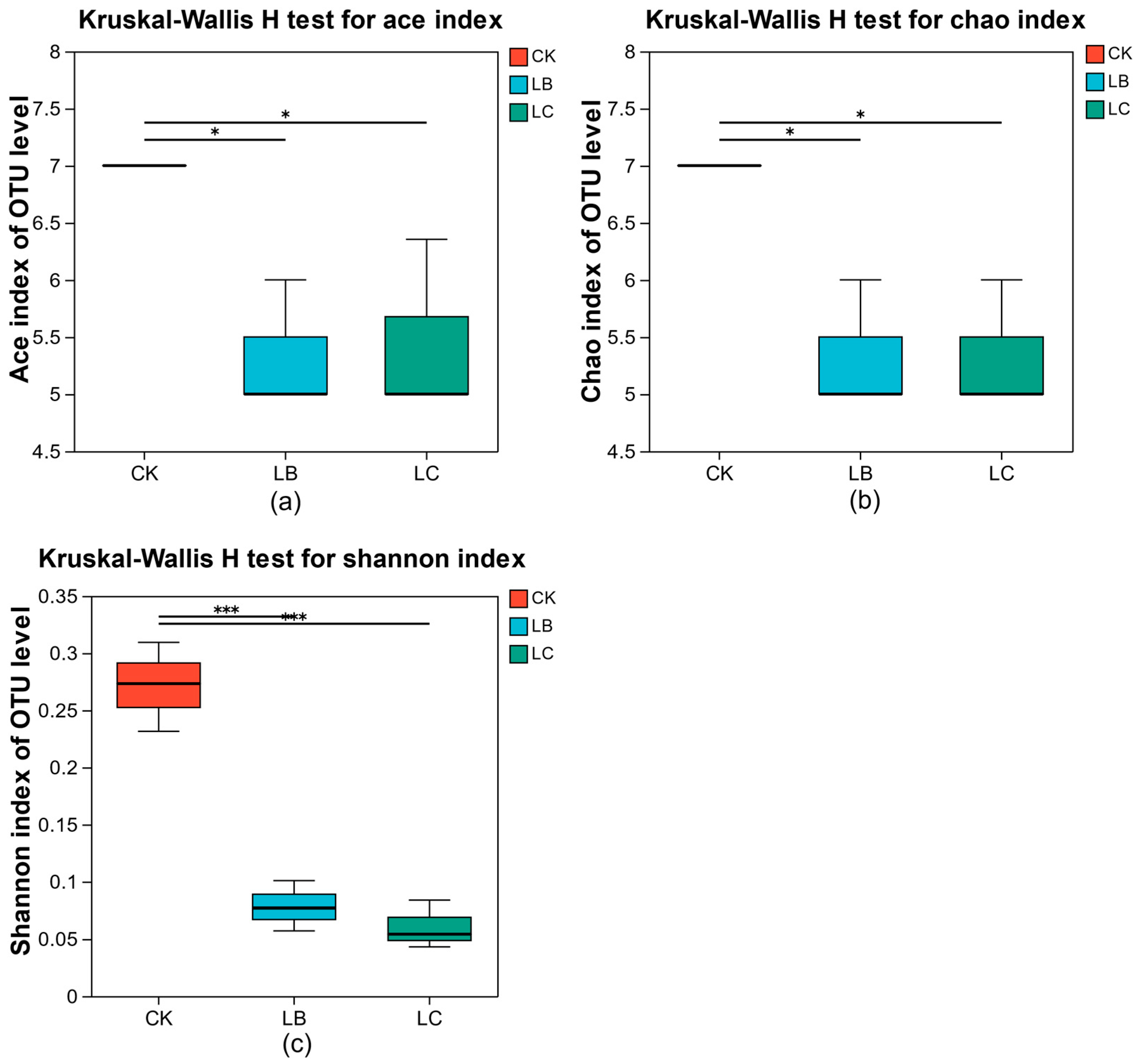
3.2.1. Changes in Colony Abundance and Diversity
3.2.2. Changes in the Structural Composition of the Microflora
3.3. Metabolomic Analysis of Fermentation Products
3.3.1. PCA Analysis of Intergroup Metabolites
3.3.2. Differential Metabolite Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ren, C.; Zhao, X.; Liu, K.; Wang, L.; Chen, Q.; Jiang, H.; Gao, X.; Lv, X.; Zhi, X.; Wu, X.; et al. Research progress of natural medicine Astragalus mongholicus Bunge in treatment of myocardial fibrosis. J. Ethnopharmacol. 2023, 305, 116128. [Google Scholar] [PubMed]
- Kalaycı, B.; Özek, N.Ş.; Aysin, F.; Özbek, H.; Kazaz, C.; Önal, M.; Güvenalp, Z.J.S.P.J. Evaluation of cytotoxic and apoptotic effects of the extracts and phenolic compounds of Astragalus globosus Vahl and Astragalus breviflorus DC. Saudi Pharm. J. 2023, 31, 101682. [Google Scholar] [PubMed]
- Qiao, Y.; Liu, C.; Guo, Y.; Zhang, W.; Guo, W.; Oleksandr, K.; Wang, Z. Polysaccharides derived from Astragalus membranaceus and Glycyrrhiza uralensis improve growth performance of broilers by enhancing intestinal health and modulating gut microbiota. Poult. Sci. 2022, 101, 101905. [Google Scholar] [PubMed]
- Zhong, R.; Yu, M.; Liu, H.; Sun, H.; Cao, Y.; Zhou, D. Effects of dietary Astragalus polysaccharide and Astragalus membranaceus root supplementation on growth performance, rumen fermentation, immune responses, and antioxidant status of lambs. Anim. Feed. Sci. Technol. 2012, 174, 60–67. [Google Scholar]
- Chen, Y.; Liu, C.; Yang, F.; Chen, H.; Yang, C.; Fan, Z.; Xiao, Y.; Xiao, D. UPLC–QQQ–MS/MS-based widely targeted metabolomic analysis, antioxidant and α-glucosidase inhibitory activities of mulberry leaves processed by solid-state fermentation. LWT 2023, 188, 115351. [Google Scholar]
- Wu, H.; Liu, H.-N.; Ma, A.-M.; Zhou, J.-Z.; Xia, X.-D. Synergetic effects of Lactobacillus plantarum and Rhizopus oryzae on physicochemical, nutritional and antioxidant properties of whole-grain oats (Avena sativa L.) during solid-state fermentation. LWT 2022, 154, 112687. [Google Scholar]
- Roasa, J.; De Villa, R.; Mine, Y.; Tsao, R. Phenolics of cereal, pulse and oilseed processing by-products and potential effects of solid-state fermentation on their bioaccessibility, bioavailability and health benefits: A review. Trends Food Sci. Technol. 2021, 116, 954–974. [Google Scholar]
- Long, M.; Pei, X.; Lu, Z.; Xu, D.; Zheng, N.; Li, Y.; Ge, H.; Cao, W.; Osire, T.; Xia, X. Effective degradation of anthraquinones in Folium Sennae with Monascus fermentation for toxicity reduce and efficacy enhancement. Heliyon 2023, 9, e18735. [Google Scholar] [PubMed]
- Qi, J.; Huang, H.; Wang, J.; Liu, N.; Chen, X.; Jiang, T.; Xu, H.; Lei, H. Insights into the improvement of bioactive phytochemicals, antioxidant activities and flavor profiles in Chinese wolfberry juice by select lactic acid bacteria. Food Biosci. 2021, 43, 101264. [Google Scholar]
- Weon, J.B.; Yun, B.-R.; Lee, J.; Eom, M.R.; Ko, H.-J.; Lee, H.Y.; Park, D.-S.; Chung, H.-C.; Chung, J.Y.; Ma, C.J. Neuroprotective effect of steamed and fermented Codonopsis lanceolata. Biomol. Ther. 2014, 22, 246. [Google Scholar]
- Yim, N.H.; Gu, M.J.; Park, H.R.; Hwang, Y.H.; Ma, J.Y. Enhancement of neuroprotective activity of Sagunja-tang by fermentation with lactobacillus strains. BMC Complement. Altern. Med. 2018, 18, 312. [Google Scholar]
- Ben Abdesslem, S.; Ben Moussa, O.; Boulares, M.; Elbaz, M.; Chouaibi, M.; Ayachi, S.; Hassouna, M. Evaluation of the effect of fennel (Foeniculum vulgare Mill) essential oil addition on the quality parameters and shelf-life prediction of yoghurt. Int. J. Dairy Technol. 2020, 73, 403–410. [Google Scholar]
- Xu, Z.; Li, M.; Wang, Y.; Feng, M.; Gan, Z.; Leng, X.; Li, X. Affecting mechanism of Chlorella sorokiniana meal replacing fish meal on growth and immunity of Litopenaeus vannamei based on transcriptome analysis. Aquac. Rep. 2023, 31, 101645. [Google Scholar]
- Luo, J.-W.; Xiao, S.; Wang, B.; Cai, Y.-X.; Wang, J.-H. In vitro fermentation of pineapple–whey protein fermentation product on human intestinal microbiota derived from fecal microbiota transplant donors. LWT 2024, 191, 115637. [Google Scholar]
- Song, X.; Wang, J.; Wang, Y.; Feng, Y.; Cui, Q.; Lu, Y. Artificial creation of Chlorella pyrenoidosa mutants for economic sustainable food production. Bioresour. Technol. 2018, 268, 340–345. [Google Scholar] [PubMed]
- Barghchi, H.; Dehnavi, Z.; Nattagh-Eshtivani, E.; Alwaily, E.R.; Almulla, A.F.; Kareem, A.K.; Barati, M.; Ranjbar, G.; Mohammadzadeh, A.; Rahimi, P.J.B.; et al. The effects of Chlorella vulgaris on cardiovascular risk factors: A comprehensive review on putative molecular mechanisms. Biomed. Pharmacother. 2023, 162, 114624. [Google Scholar]
- Geada, P.; Francisco, D.; Pereira, F.; Maciel, F.; Madureira, L.; Barros, A.; Silva, J.L.; Vicente, A.A.; Teixeira, J.A. Multivariable optimization process of heterotrophic growth of Chlorella vulgaris. Food Bioprod. Process. 2023, 138, 1–13. [Google Scholar]
- Zhou, J.; Wang, M.; Bäuerl, C.; Cortés-Macías, E.; Calvo-Lerma, J.; Collado, M.C.; Barba, F.J. The impact of liquid-pressurized extracts of Spirulina, Chlorella and Phaedactylum tricornutum on in vitro antioxidant, antiinflammatory and bacterial growth effects and gut microbiota modulation. Food Chem. 2023, 401, 134083. [Google Scholar]
- Solano, R.; Patino-Ruiz, D.; Herrera, A. Preparation of modified paints with nano-structured additives and its potential applications. Nanomater. Nanotechnol. 2020, 10, 1847980420909188. [Google Scholar]
- Peng, K.; Chen, X.; Wei, D.; Zhao, L.; Chen, B.; Mo, W.; Zheng, C.; Sun, Y. Inclusion of Chlorella water extract in Oreochromis niloticus fingerling diets: Effects on growth performance, body composition, digestive enzyme activity, antioxidant and immune capacity, intestine and hepatic histomorphology and sodium nitrite stress resistance. Aquac. Rep. 2020, 18, 100547. [Google Scholar]
- Liu, B.; Lu, H.; Shu, Q.; Chen, Q.; Wang, J. The influence of different pretreatment methods of highland barley by solid-state fermentation with Agaricus sinodeliciosus var. Chaidam ZJU-TP-08 on its nutrient content, functional properties and physicochemical characteristics. J. Fungi 2022, 8, 940. [Google Scholar] [CrossRef]
- Wang, S.; Lu, L.; Song, T.; Xu, X.; Yu, J.; Liu, T.J.H. Optimization of Cordyceps sinensis fermentation Marsdenia tenacissima process and the differences of metabolites before and after fermentation. Heliyon 2022, 8, e12586. [Google Scholar] [PubMed]
- Niazi, A.; Mehrgan, M.S.; Islami, H.R.J.A. Optimizing turmeric and green tea fermentation with Lactobacillus brevis to enhance growth performance, digestive enzymes, and immunity in rainbow trout. Aquaculture 2023, 577, 739962. [Google Scholar]
- Wang, M. Evaluation of Biological Properties of Extracts Obtained from Marine Biomass Assisted by Pulsed Electric Fields (PEF) and Pressurized Liquid Extraction (PLE). Ph.D. Thesis, Universitat de València, Valencia, Spain, 2022. [Google Scholar]
- Chen, Z.; Zhou, P.; Zhao, Z.; Li, B. Enhanced production of L-serine in Escherichia coli by fermentation process optimization and transcriptome sequencing. Biochem. Eng. J. 2023, 200, 109109. [Google Scholar]
- Yang, Y.; Na Wu, Y.; Ce, L.G.E.; Ge, X.G.B.R.; Shuang, Q.; Zhang, F.M. Analysis of microbial community and its correlation with flavor compounds during Congee fermentation. Food Biosci. 2023, 51, 102261. [Google Scholar]
- Guo, Q.; Zabed, H.; Zhang, H.; Wang, X.; Yun, J.; Zhang, G.; Yang, M.; Sun, W.; Qi, X. Optimization of fermentation medium for a newly isolated yeast strain (Zygosaccharomyces rouxii JM-C46) and evaluation of factors affecting biosynthesis of D-arabitol. LWT 2019, 99, 319–327. [Google Scholar]
- Zhou, S.; Ding, N.; Han, R.; Deng, Y. Metabolic engineering and fermentation optimization strategies for producing organic acids of the tricarboxylic acid cycle by microbial cell factories. Bioresour. Technol. 2023, 379, 128986. [Google Scholar]
- Bitew, D.; Alemu, M.; Tesfaye, A.; Andualem, B.J.R.; Reviews, S.E. Ethanologenic yeasts from Ethiopian fermented beverages and optimization of fermentation conditions. Renew. Sustain. Energy Rev. 2024, 190, 114090. [Google Scholar]
- Ni, K.; Wang, X.; Lu, Y.; Guo, L.; Li, X.; Yang, F. Exploring the silage quality of alfalfa ensiled with the residues of astragalus and hawthorn. Bioresour. Technol. 2020, 297, 122249. [Google Scholar]
- Zheng, Z.; Wei, L.; Zhu, M.; Qian, Z.; Liu, J.; Zhang, L.; Xu, Y. Effect of lactic acid bacteria co-fermentation on antioxidant activity and metabolomic profiles of a juice made from wolfberry and longan. Food Res. Int. 2023, 174, 113547. [Google Scholar]
- Guo, M.; Kong, Q.; Wang, W.; Yu, H. Biotransformation of amygdalin by lactic acid bacteria fermentation. Process Biochem. 2023, 132, 221–227. [Google Scholar]
- Shi, H.; Zhao, Y.; Wang, W.; Zhou, Y.; Liang, Y.; Wu, R.; Wu, J. The potential of lactic acid bacteria in fermented herbs-derived food products. Food Biosci. 2024, 61, 104714. [Google Scholar]
- Akpabli-Tsigbe, N.D.K.; Ma, Y.; Ekumah, J.-N.; Osabutey, J.; Hu, J.; Xu, M.; Johnson, N.A.N.J.L. Novel solid-state fermentation extraction of 5-O-caffeoylquinic acid from heilong48 soybean using Lactobacillus helviticus: Parametric screening and optimization. LWT 2021, 149, 111809. [Google Scholar]
- Kang, H.K.; Salim, H.M.; Akter, N.; Kim, D.W.; Kim, J.H.; Bang, H.T.; Kim, M.J.; Na, J.C.; Hwangbo, J.; Choi, H.C.; et al. Effect of various forms of dietary Chlorella supplementation on growth performance, immune characteristics, and intestinal microflora population of broiler chickens. J. Appl. Poult. Res. 2013, 22, 100–108. [Google Scholar]
- Kaga, Y.; Kuda, T.; Taniguchi, M.; Yamaguchi, Y.; Takenaka, H.; Takahashi, H.; Kimura, B.J.L. The effects of fermentation with lactic acid bacteria on the antioxidant and anti-glycation properties of edible cyanobacteria and microalgae. LWT 2021, 135, 110029. [Google Scholar]
- Zhang, R.; Song, X.; Liu, W.; Gao, X. Mixed fermentation of Chlorella pyrenoidosa and Bacillus velezensis SW-37 by optimization. LWT 2023, 175, 114448. [Google Scholar]
- Wu, T.; Deng, C.; Luo, S.; Liu, C.; Hu, X. Effect of rice bran on properties of yogurt: Comparison between addition of bran before fermentation and after fermentation. Food Hydrocoll. 2023, 135, 108122. [Google Scholar]
- Jiang, Y.; Wu, J.; Tian, L.; Liu, Y.; Zhao, F.; He, Z.; Mao, Y.; Jia, J.; Guan, T. The therapeutic effects of fermented milk with lactic acid bacteria from traditional Daqu on hypertensive mice. J. Dairy Sci. 2024, 107, 742–758. [Google Scholar]
- Charlier, C.; Cretenet, M.; Even, S.; Le Loir, Y. Interactions between Staphylococcus aureus and lactic acid bacteria: An old story with new perspectives. Int. J. Food Microbiol. 2009, 131, 30–39. [Google Scholar]
- Pau, S.; Tan, L.C.; Arriaga, S.; Lens, P.N. Lactic acid fermentation of food waste in a semicontinuous SBR system: Influence of the influent composition and hydraulic retention time. Environ. Technol. 2024, 45, 2993–3003. [Google Scholar]
- Zhao, W.; Zhang, Z.; Gao, Y.; Liu, X.; Du, C.; Ma, F.; Wang, S.; Shi, W.; Yang, Y.; Deng, R.; et al. Fungal dynamic changes in naturally fermented ‘Kyoho’grape juice. Arch. Microbiol. 2022, 204, 556. [Google Scholar]
- Zhao, X.; Tang, F.; Cai, W.; Peng, B.; Zhang, P.; Shan, C. Effect of fermentation by lactic acid bacteria on the phenolic composition, antioxidant activity, and flavor substances of jujube–wolfberry composite juice. LWT 2023, 184, 114884. [Google Scholar]
- Huang, W.; Zhang, C.; Gu, Z.; Li, C.; Fang, Z.; Zeng, Z.; Zhang, Z.; Hu, B.; Chen, H.; Wu, W.; et al. Effect of microbial fermentation on the sensory characteristics and chemical compositions of Chinese sweet tea (Lithocarpus litseifolius (Hance) Chun). Food Biosci. 2022, 46, 101567. [Google Scholar]
- Wang, X.; Ma, Y.; Xu, Q.; Shikov, A.N.; Pozharitskaya, O.N.; Flisyuk, E.V.; Liu, M.; Li, H.; Vargas-Murga, L.; Duez, P.J.P. Flavonoids and saponins: What have we got or missed? Phytomedicine 2023, 109, 154580. [Google Scholar] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Liu, W.; Ge, Q.; Xu, T.; Wu, X.; Zhong, R. Enhancement of Active Substances in Astragali Radix Broth with Lactic Acid Bacteria Fermentation and the Promotion Role of Chlorella Growth Factor. Fermentation 2024, 10, 455. https://doi.org/10.3390/fermentation10090455
Li X, Liu W, Ge Q, Xu T, Wu X, Zhong R. Enhancement of Active Substances in Astragali Radix Broth with Lactic Acid Bacteria Fermentation and the Promotion Role of Chlorella Growth Factor. Fermentation. 2024; 10(9):455. https://doi.org/10.3390/fermentation10090455
Chicago/Turabian StyleLi, Xiaomeng, Wei Liu, Qingyan Ge, Tongtong Xu, Xiang Wu, and Ruohui Zhong. 2024. "Enhancement of Active Substances in Astragali Radix Broth with Lactic Acid Bacteria Fermentation and the Promotion Role of Chlorella Growth Factor" Fermentation 10, no. 9: 455. https://doi.org/10.3390/fermentation10090455