
Solid-State Fermentation of Grain-Derived By-Products by Aspergillus kawachii and Rhizopus oryzae: Preparation and Evaluation of Anti-Allergic Activity



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Culture, Chemicals and Reagents
2.2. Solid-State Fermentation of Grain-Derived by Products by A. kawachii L1 and R. oryzae L1
2.3. Determination of Total Phenolics, Flavonoids and Amino Acids Contents
2.4. Ethics Statement and Conduct of Animal Experiments
2.5. Cultivation of Splenocytes and Quantification of Cytokines and Immunoglobulins
2.6. Statistical Analysis
3. Results
3.1. Appearance and Contents of Total Phenolics, Flavonoids and Amino Acids of the Solid-State Fermented Grain Product Extract
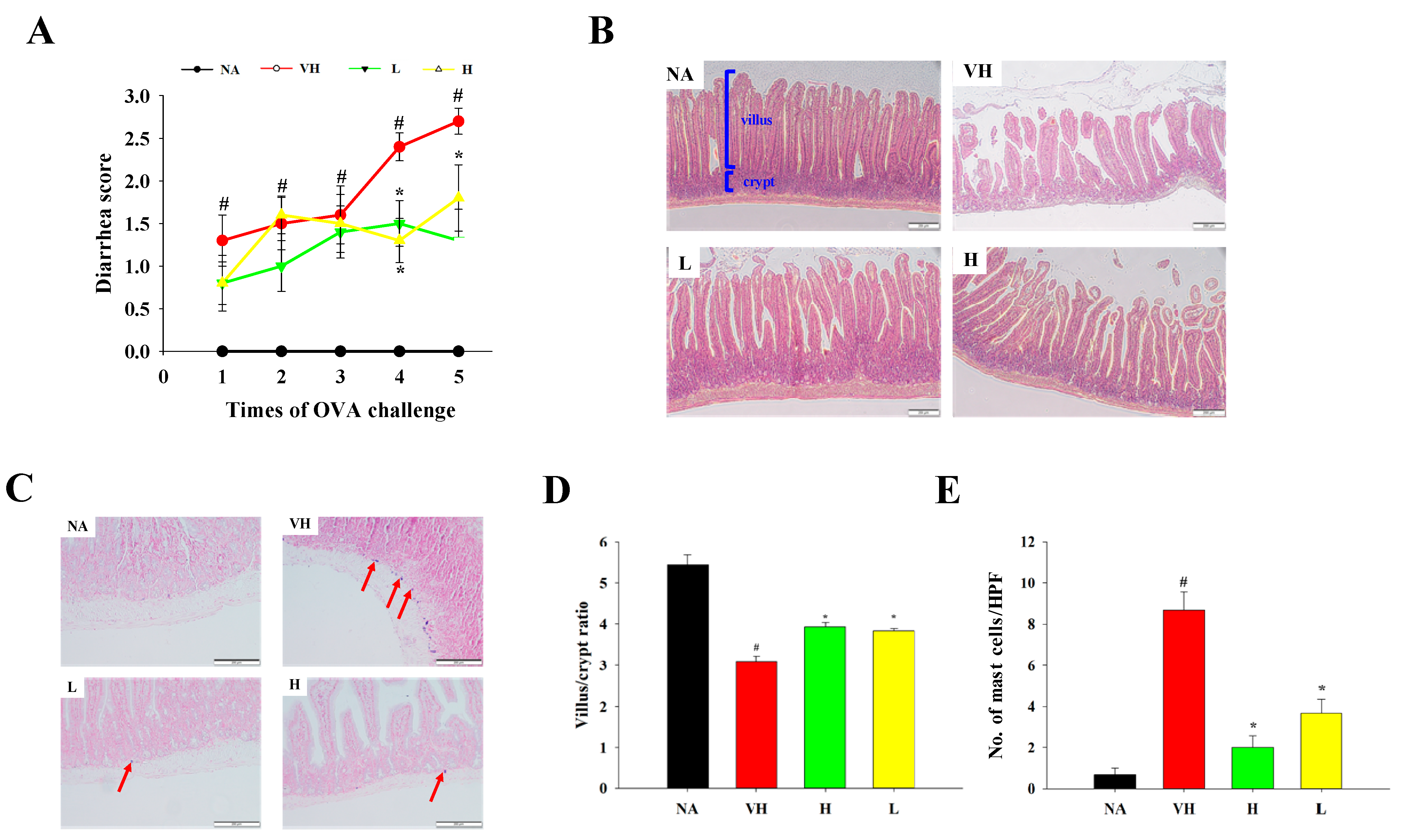
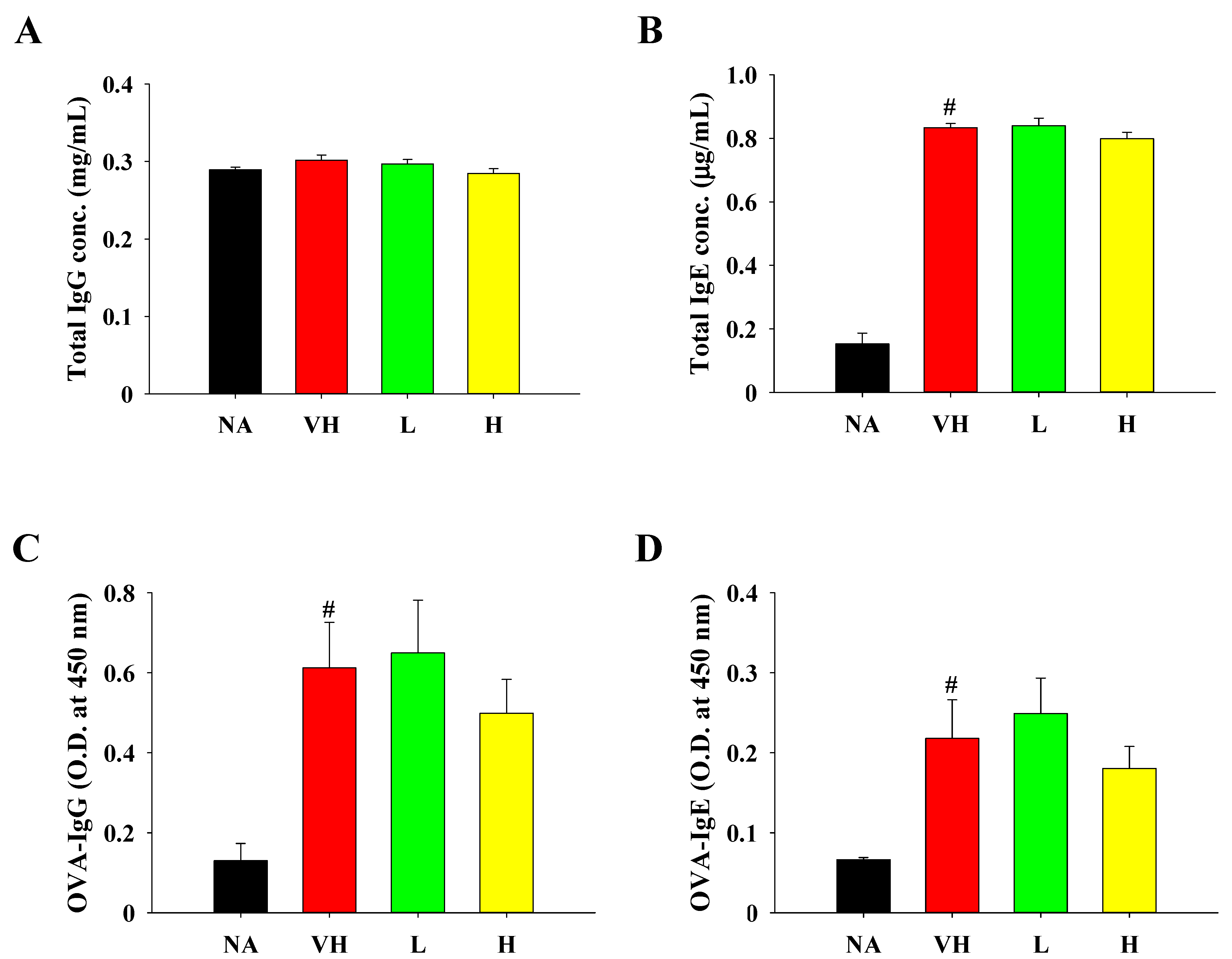
3.2. Effects of Solid-State Fermented Grain Product Extract on Attenuating Allergen-Induced Enteritis and Modulating Cytokine Production
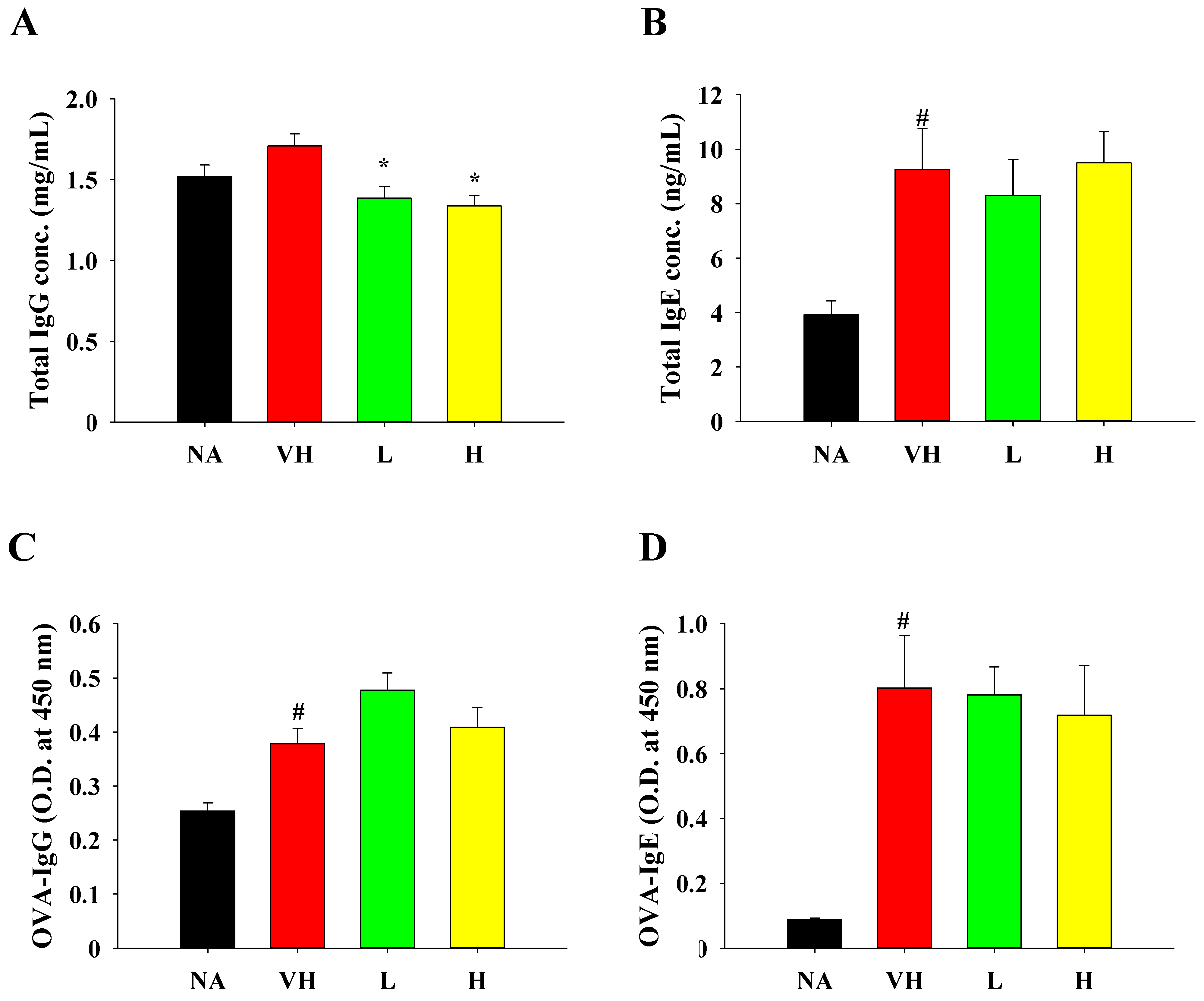
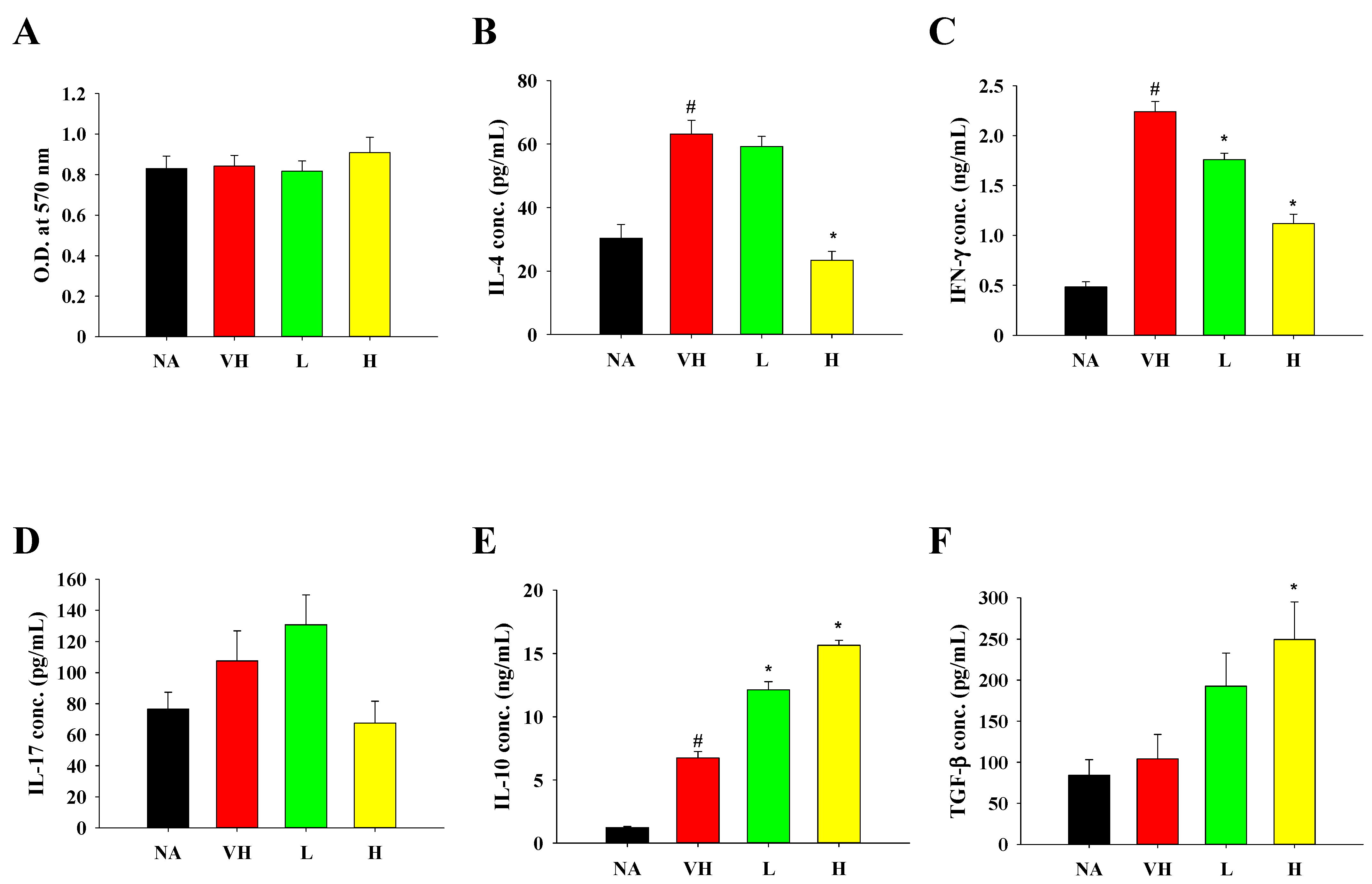
3.3. Effects of Solid-State Fermented Grain Product Extract on Ameliorating Allergen-Induced DTH and Modulating Cytokine Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fărcaș, A.C.; Socaci, S.A.; Nemeș, S.A.; Pop, O.L.; Coldea, T.E.; Fogarasi, M.; Biriș-Dorhoi, E.S. An update regarding the bioactive compound of cereal by-products: Health benefits and potential applications. Nutrients 2022, 14, 3470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, C.; Wang, W.; Wang, W.; Wei, D. Construction of enhanced transcriptional activators for improving cellulase production in Trichoderma reesei RUT C30. Bioresour. Bioprocess. 2018, 5, 40. [Google Scholar] [CrossRef]
- Bhargav, S.; Panda, B.P.; Ali, M.; Javed, S. Solid-state fermentation: An overview. Chem. Biochem. Eng. Q. 2008, 22, 49–70. [Google Scholar]
- Pandey, A.; Soccol, C.R.; Mitchell, D. New developments in solid state fermentation: I-bioprocesses and products. Process Biochem. 2000, 35, 1153–1169. [Google Scholar] [CrossRef]
- Abd Razak, D.L.; Abd Rashid, N.Y.; Jamaluddin, A.; Sharifudin, S.A.; Abd Kahar, A.; Long, K. Cosmeceutical potentials and bioactive compounds of rice bran fermented with single and mix culture of Aspergillus oryzae and Rhizopus oryzae. J. Saudi Soc. Agric. Sci. 2017, 16, 127–134. [Google Scholar] [CrossRef]
- Espitia-Hernández, P.; Ruelas-Chacón, X.; Chávez-González, M.L.; Ascacio-Valdés, J.A.; Flores-Naveda, A.; Sepúlveda-Torre, L. Solid-state fermentation of Sorghum by Aspergillus oryzae and Aspergillus niger: Effects on tannin content, phenolic profile, and antioxidant activity. Foods 2022, 11, 3121. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, Y.; Chen, B.; Wang, X.; Qiao, Y.; Chen, J. Combined treatment of rice bran by solid-state fermentation and extrusion: Effect of processing sequence and microbial strains. Food Chem. X 2024, 23, 101549. [Google Scholar] [CrossRef]
- Kajiwara, Y.; Takeshima, N.; Ohba, H.; Omori, T.; Shimoda, M.; Wada, H. Production of acid-stable α-amylase by Aspergillus kawachii during barley Shochu-Koji production. J. Ferment. Bioeng. 1997, 84, 224–227. [Google Scholar] [CrossRef]
- Kum, S.-J.; Yang, S.-O.; Lee, S.M.; Chang, P.-S.; Choi, Y.H.; Lee, J.J.; Hurh, B.S.; Kim, Y.-S. Effects of Aspergillus species inoculation and their enzymatic activities on the formation of volatile components in fermented soybean paste (doenjang). J. Agric. Food Chem. 2015, 63, 1401–1418. [Google Scholar] [CrossRef]
- Cho, H.-D.; Min, H.-J.; Won, Y.-S.; Ahn, H.-Y.; Cho, Y.-S.; Seo, K.-I. Solid state fermentation process with Aspergillus kawachii enhances the cancer-suppressive potential of silkworm larva in hepatocellular carcinoma cells. BMC Complement. Altern. Med. 2019, 19, 241. [Google Scholar] [CrossRef]
- Chen, T.-Y.; Chen, Y.-L.; Chiu, W.-C.; Yeh, C.-L.; Tung, Y.-T.; Shirakawa, H.; Liao, W.-T.; Yang, S.-C. Effects of the water extract of fermented rice bran on liver damage and intestinal injury in aged rats with high-fat diet feeding. Plants 2022, 11, 607. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.C.; da Silva Graça, C.; Chiattoni, L.M.; Massarolo, K.C.; Duarte, F.A.; Mellado, M.d.L.S.; de Souza Soares, L.A. Fermentation process in the availability of nutrients in rice bran. Res. Rev. J. Microbiol. Biotechnol. 2017, 6, 45–52. [Google Scholar]
- Abbas, A.K.; Murphy, K.M.; Sher, A. Functional diversity of helper T lymphocytes. Nature 1996, 383, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar] [PubMed]
- Crow, M.K.; Kirou, K.A. Interferon-α in systemic lupus erythematosus. Curr. Opin. Rheumatol. 2004, 16, 541–547. [Google Scholar] [CrossRef]
- Gordon, J.N.; Di Sabatino, A.; MacDonald, T.T. The pathophysiologic rationale for biological therapies in inflammatory bowel disease. Curr. Opin. Gastroenterol. 2005, 21, 431–437. [Google Scholar]
- Wu, C.; Yang, G.; Bermúdez-Humarán, L.G.; Pang, Q.; Zeng, Y.; Wang, J.; Gao, X. Immunomodulatory effects of IL-12 secreted by Lactococcus lactis on Th1/Th2 balance in ovalbumin (OVA)-induced asthma model mice. Int. Immunopharmacol. 2006, 6, 610–615. [Google Scholar] [CrossRef]
- Palomares, O.; Martin-Fontecha, M.; Lauener, R.; Traidl-Hoffmann, C.; Cavkaytar, O.; Akdis, M.; Akdis, C. Regulatory T cells and immune regulation of allergic diseases: Roles of IL-10 and TGF-β. Genes Immun. 2014, 15, 511–520. [Google Scholar] [CrossRef]
- Elghoudi, A.; Narchi, H. Food allergy in children—The current status and the way forward. World J. Clin. Pediatr. 2022, 11, 253. [Google Scholar] [CrossRef]
- Anvari, S.; Miller, J.; Yeh, C.-Y.; Davis, C.M. IgE-mediated food allergy. Clin. Rev. Allergy Immunol. 2019, 57, 244–260. [Google Scholar] [CrossRef]
- Kraneveld, A.; Buckley, T.; van Heuven-Nolsen, D.; Van Schaik, Y.; Koster, A.S.; Nijkamp, F. Delayed-type hypersensitivity-induced increase in vascular permeability in the mouse small intestine: Inhibition by depletion of sensory neuropeptides and NK1 receptor blockade. Br. J. Pharmacol. 1995, 114, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Chen, T.-Y.; Tsai, G.-J. Hypouricemic Effect of Submerged Culture of Ganoderma lucidum in Potassium Oxonate-Induced Hyperuricemic Rats. Metabolites 2022, 12, 553. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-Z.; Hu, C.-M.; Huang, C.-H.; Wey, S.-P.; Jan, T.-R. Cannabidiol attenuates delayed-type hypersensitivity reactions via suppressing T-cell and macrophage reactivity. Acta Pharmacol. Sin. 2010, 31, 1611–1617. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Pan, C.-L.; Tsai, G.-J.; Chang, C.-J.; Tsai, W.-C.; Lu, S.-Y. Anti-allergic diarrhea effect of diosgenin occurs via improving gut dysbiosis in a murine model of food allergy. Molecules 2021, 26, 2471. [Google Scholar] [CrossRef]
- Huang, C.-H.; Lin, Y.-C.; Jan, T.-R. Lactobacillus reuteri induces intestinal immune tolerance against food allergy in mice. J. Funct. Foods 2017, 31, 44–51. [Google Scholar] [CrossRef]
- Singh, A.; Holvoet, S.; Mercenier, A. Dietary polyphenols in the prevention and treatment of allergic diseases. Clin. Exp. Allergy 2011, 41, 1346–1359. [Google Scholar] [CrossRef]
- Chatchatee, P.; Nowak-Wegrzyn, A.; Lange, L.; Benjaponpitak, S.; Chong, K.W.; Sangsupawanich, P.; van Ampting, M.T.; Nijhuis, M.M.O.; Harthoorn, L.F.; Langford, J.E. Tolerance development in cow’s milk–allergic infants receiving amino acid–based formula: A randomized controlled trial. J. Allergy Clin. Immunol. 2022, 149, 650–658.e5. [Google Scholar] [CrossRef]
- Rakha, A.; Umar, N.; Rabail, R.; Butt, M.S.; Kieliszek, M.; Hassoun, A.; Aadil, R.M. Anti-inflammatory and anti-allergic potential of dietary flavonoids: A review. Biomed. Pharmacother. 2022, 156, 113945. [Google Scholar] [CrossRef]
- Ngoc, L.P.; Gold, D.R.; Tzianabos, A.O.; Weiss, S.T.; Celedon, J.C. Cytokines, allergy, and asthma. Curr. Opin. Allergy Clin. Immunol. 2005, 5, 161–166. [Google Scholar] [CrossRef]
- Pollard, K.M.; Cauvi, D.M.; Toomey, C.B.; Morris, K.V.; Kono, D.H. Interferon-γ and systemic autoimmunity. Discov. Med. 2013, 16, 123. [Google Scholar]
- Pène, J.R.M.; Chevalier, S.; Preisser, L.; Vénéreau, E.; Guilleux, M.-H.L.N.; Ghannam, S.; Molès, J.-P.; Danger, Y.; Ravon, E.; Lesaux, S. Chronically inflamed human tissues are infiltrated by highly differentiated Th17 lymphocytes. J. Immunol. 2008, 180, 7423–7430. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef]
- Gorelik, L.; Flavell, R.A. Transforming growth factor-β in T-cell biology. Nat. Rev. Immunol. 2002, 2, 46–53. [Google Scholar] [CrossRef]
- Park, H.-Y.; Lee, K.-W.; Choi, H.-D. Rice bran constituents: Immunomodulatory and therapeutic activities. Food Funct. 2017, 8, 935–943. [Google Scholar] [CrossRef]
- Yao, W.; Gong, Y.; Li, L.; Hu, X.; You, L. The effects of dietary fibers from rice bran and wheat bran on gut microbiota: An overview. Food Chem. X 2022, 13, 100252. [Google Scholar] [CrossRef] [PubMed]
- Hidvégi, M.; Rásó, E.; Farkas, R.T.; Lapis, K.; Szende, B. Effect of MSC on the immune response of mice. Immunopharmacology 1999, 41, 183–186. [Google Scholar] [CrossRef]
- Sukkar, S.G.; Rossi, E. Oxidative stress and nutritional prevention in autoimmune rheumatic diseases. Autoimmun. Rev. 2004, 3, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; Santos-Sánchez, G.; Pedroche, J.; Fernández-Pachón, M.-S.; Millán, F.; Millán-Linares, M.C.; Lardone, P.J.; Bejarano, I.; Guerrero, J.M. Immunomodulatory and antioxidant properties of wheat gluten protein hydrolysates in human peripheral blood mononuclear cells. Nutrients 2020, 12, 1673. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Shi, Y.; Han, F.; Sun, J.; Le, G. Study on Wheat Peptides on Immune Modulating and Antioxidant Effect. Nat. Prod. Res. Dev. 2009, 21, 473. [Google Scholar]
- Moore, J.; Cheng, Z.; Hao, J.; Guo, G.; Liu, J.-G.; Lin, C.; Yu, L. Effects of solid-state yeast treatment on the antioxidant properties and protein and fiber compositions of common hard wheat bran. J. Agric. Food Chem. 2007, 55, 10173–10182. [Google Scholar] [CrossRef]
- Bhanja, T.; Kumari, A.; Banerjee, R. Enrichment of phenolics and free radical scavenging property of wheat koji prepared with two filamentous fungi. Bioresour. Technol. 2009, 100, 2861–2866. [Google Scholar] [CrossRef] [PubMed]
- Đorđević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- Cai, S.; Wang, O.; Wu, W.; Zhu, S.; Zhou, F.; Ji, B.; Gao, F.; Zhang, D.; Liu, J.; Cheng, Q. Comparative study of the effects of solid-state fermentation with three filamentous fungi on the total phenolics content (TPC), flavonoids, and antioxidant activities of subfractions from oats (Avena sativa L.). J. Agric. Food Chem. 2012, 60, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Kimoto-Nira, H.; Moriya, N.; Nogata, Y.; Sekiyama, Y.; Toguchi, Y. Fermentation of Shiikuwasha (Citrus depressa Hayata) pomace by lactic acid bacteria to generate new functional materials. Int. J. Food Sci. Technol. 2019, 54, 688–695. [Google Scholar] [CrossRef]
- MacDonald, T.T.; Monteleone, G. Immunity, inflammation, and allergy in the gut. Science 2005, 307, 1920–1925. [Google Scholar] [CrossRef]
- Zuercher, A.; Holvoet, S.; Weiss, M.; Mercenier, A. Polyphenol-enriched apple extract attenuates food allergy in mice. Clin. Exp. Allergy 2010, 40, 942–950. [Google Scholar] [CrossRef]
- Li, Y.; Mattison, C.P. Polyphenol-rich pomegranate juice reduces IgE binding to cashew nut allergens. J. Sci. Food Agric. 2018, 98, 1632–1638. [Google Scholar] [CrossRef]
- Bansode, R.R.; Randolph, P.D.; Plundrich, N.J.; Lila, M.A.; Williams, L.L. Peanut protein-polyphenol aggregate complexation suppresses allergic sensitization to peanut by reducing peanut-specific IgE in C3H/HeJ mice. Food Chem. 2019, 299, 125025. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, T.; Hou, J.; Li, M. Ferulic acid alleviates atopic dermatitis-like symptoms in mice via its potent anti-inflammatory effect. Immunopharmacol. Immunotoxicol. 2020, 42, 156–164. [Google Scholar] [CrossRef]
- Sun, H.; Chen, Y.; Cheng, M.; Zhang, X.; Zheng, X.; Zhang, Z. The modulatory effect of polyphenols from green tea, oolong tea and black tea on human intestinal microbiota in vitro. J. Food Sci. Technol. 2018, 55, 399–407. [Google Scholar] [CrossRef]
- Turroni, F.; Milani, C.; Duranti, S.; Lugli, G.A.; Bernasconi, S.; Margolles, A.; Di Pierro, F.; Van Sinderen, D.; Ventura, M. The infant gut microbiome as a microbial organ influencing host well-being. Ital. J. Pediatr. 2020, 46, 16. [Google Scholar] [CrossRef]
- Magrone, T.; Jirillo, E. Influence of polyphenols on allergic immune reactions: Mechanisms of action. Proc. Nutr. Soc. 2012, 71, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Wei, D.; Bian, T.; Xie, P.; Zou, J.; Mu, H.; Zhang, B.; Zhou, X. Genistein attenuated allergic airway inflammation by modulating the transcription factors T-bet, GATA-3 and STAT-6 in a murine model of asthma. Pharmacology 2012, 89, 229–236. [Google Scholar] [CrossRef]
- Jang, T.Y.; Jung, A.-Y.; Kyung, T.-S.; Kim, D.-Y.; Hwang, J.-H.; Kim, Y.H. Anti-allergic effect of luteolin in mice with allergic asthma and rhinitis. Cent. Eur. J. Immunol. 2017, 42, 24–29. [Google Scholar] [CrossRef]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S. Anti-inflammatory effect of quercetin and galangin in LPS-stimulated RAW264. 7 macrophages and DNCB-induced atopic dermatitis animal models. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar] [PubMed]
- Vo, T.S.; Le, T.T.; Kim, S.Y.; Ngo, D.H. The role of myricetin from Rhodomyrtus tomentosa (Aiton) Hassk fruits on downregulation of FcɛRI-mediated mast cell activation. J. Food Biochem. 2020, 44, e13143. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Jeong, G.S. Therapeutic effect of kaempferol on atopic dermatitis by attenuation of T cell activity via interaction with multidrug resistance-associated protein 1. Br. J. Pharmacol. 2021, 178, 1772–1788. [Google Scholar] [CrossRef]
- Okumo, T.; Furuta, A.; Kimura, T.; Yusa, K.; Asano, K.; Sunagawa, M. Inhibition of angiogenic factor productions by quercetin in vitro and in vivo. Medicines 2021, 8, 22. [Google Scholar] [CrossRef]
- Yuan, A.; Zeng, J.; Zhou, H.; Liu, Q.; Rao, Z.; Gao, M.; Liu, R.; Zeng, N. Anti-type I allergic effects of Jing-Fang powder extracts via PI3K/Akt pathway in vitro and in vivo. Mol. Immunol. 2021, 135, 408–420. [Google Scholar] [CrossRef]
- Candy, D.C.; Van Ampting, M.T.; Oude Nijhuis, M.M.; Wopereis, H.; Butt, A.M.; Peroni, D.G.; Vandenplas, Y.; Fox, A.T.; Shah, N.; West, C.E. A synbiotic-containing amino-acid-based formula improves gut microbiota in non-IgE-mediated allergic infants. Pediatr. Res. 2018, 83, 677–686. [Google Scholar] [CrossRef]
- Nocerino, R.; Di Scala, C.; Coppola, S.; Giglio, V.; Carucci, L.; Cosenza, L.; Voto, L.; Iannicelli, A.M.; Luzzetti, A.; Berni Canani, R. Tolerability of a new amino acid-based formula for children with IgE-mediated cow’s milk allergy. Ital. J. Pediatr. 2021, 47, 151. [Google Scholar] [CrossRef]
- Ma, J.; Wang, C.; Wang, F.; Zhang, Y.; Liu, Y.; Zhang, J.; Gao, Z.; Zhang, Y.; Xie, H.; Wang, Y. Intestinal proline is a potential anti-allergy factor for allergy diagnosis and therapy. Front. Nutr. 2022, 9, 1036536. [Google Scholar] [CrossRef] [PubMed]
- Silwal, P.; Choi, S.; Shin, K.; Lee, C.Y.; Park, J.I.; Heo, J.-Y.; Lim, K.; Park, S.-K. Histidine Suppresses IgE-Mediated Allergic Responses. J. Korean Soc. Food Sci. Nutr. 2018, 47, 697–702. [Google Scholar] [CrossRef]
- Ariaee, N.; Sankian, M.; Varasteh, A.; Hosseinpour, M.; Jabbari, F. Determining the Effect of Amino Acids on the Allergenic Activity of Pollen Extracts. Rep. Biochem. Mol. Biol. 2020, 8, 394. [Google Scholar]
- Lin, S.-P.; Chou, Y.-C.; Chen, C.-E.; Lai, Y.-T.; Hsieh, C.-C.; Tsai, T.-Y.; Lin, Y.-H.; Santoso, S.P.; Lin, H.-W.; Cheng, K.-C. Enhancing Miso Production with Optimized Puffed-Rice Koji and its Biological Function Evaluation. Food Biosci. 2024, 60, 104463. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Yu, S.-H.; Kuo, H.-C.; Cheng, K.-W.; Hsu, C.-C.; Lin, Y.-P.; Khumsupan, D.; Lin, S.-P.; Angkawijaya, A.E.; Cheng, K.-C. Alleviation of PM2. 5-induced alveolar macrophage inflammation using extract of fermented Chenopodium formosanum Koidz sprouts via regulation of NF-κB pathway. J. Ethnopharmacol. 2024, 318, 116980. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-H.; Liao, Y.-M.; Tsai, G.-J. Solid-State Fermentation of Grain-Derived By-Products by Aspergillus kawachii and Rhizopus oryzae: Preparation and Evaluation of Anti-Allergic Activity. Fermentation 2024, 10, 457. https://doi.org/10.3390/fermentation10090457
Huang C-H, Liao Y-M, Tsai G-J. Solid-State Fermentation of Grain-Derived By-Products by Aspergillus kawachii and Rhizopus oryzae: Preparation and Evaluation of Anti-Allergic Activity. Fermentation. 2024; 10(9):457. https://doi.org/10.3390/fermentation10090457
Chicago/Turabian StyleHuang, Chung-Hsiung, Yu-Ming Liao, and Guo-Jane Tsai. 2024. "Solid-State Fermentation of Grain-Derived By-Products by Aspergillus kawachii and Rhizopus oryzae: Preparation and Evaluation of Anti-Allergic Activity" Fermentation 10, no. 9: 457. https://doi.org/10.3390/fermentation10090457