

Hemp Seed Protein Hydrolysate Enriched with γ-Aminobutyric Acid and Peptides by Microbial Bioconversion

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Starter Cultures
2.3. Production of Alkaline Protease and Hydrolysate
2.4. Production of GABA
2.5. Viable Bacterial Counts
2.6. pH and Titratable Acidity
2.7. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.8. Tyrosine Equivalent Peptide
2.9. Quantitative Analysis of Free Amino Acids
2.10. Qualitative Analysis of GABA
2.11. Quantitative Analysis of the Free Amino Acids
2.12. Statistical Analysis
3. Results and Discussions
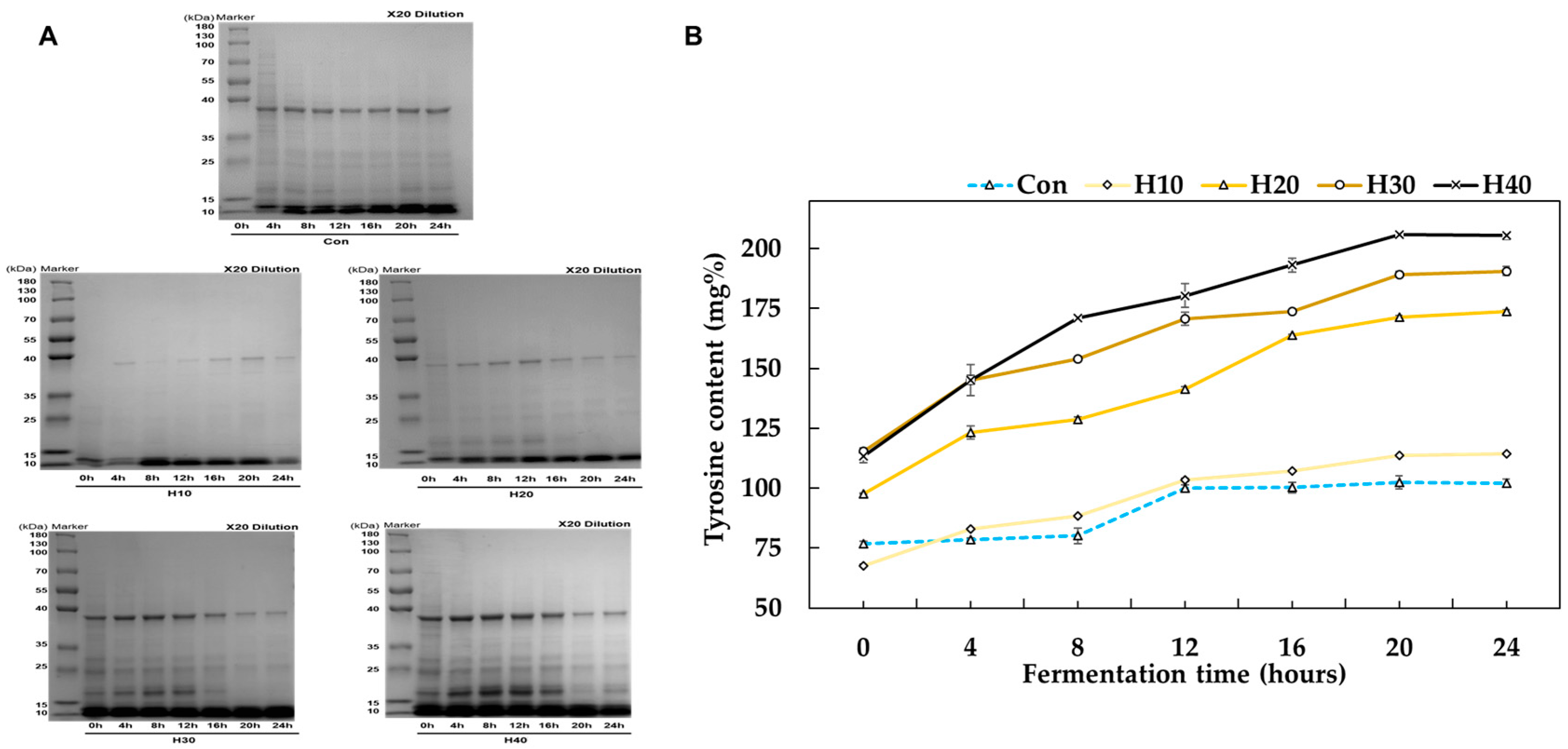
3.1. Preparation of Hemp Seed Protein (HSP) Hydrolysate
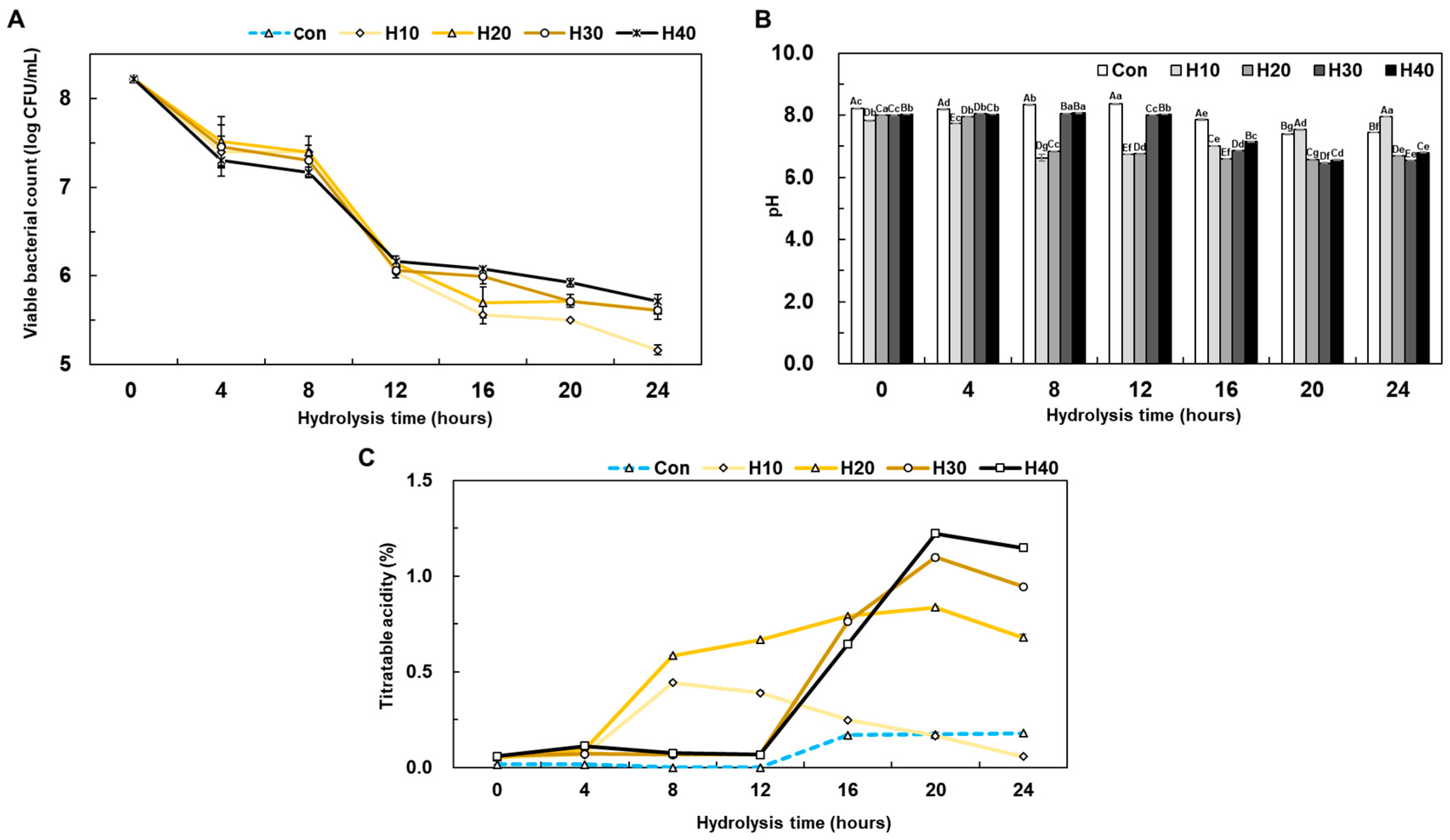
3.2. Physicochemical Properties of HSP Hydrolysate
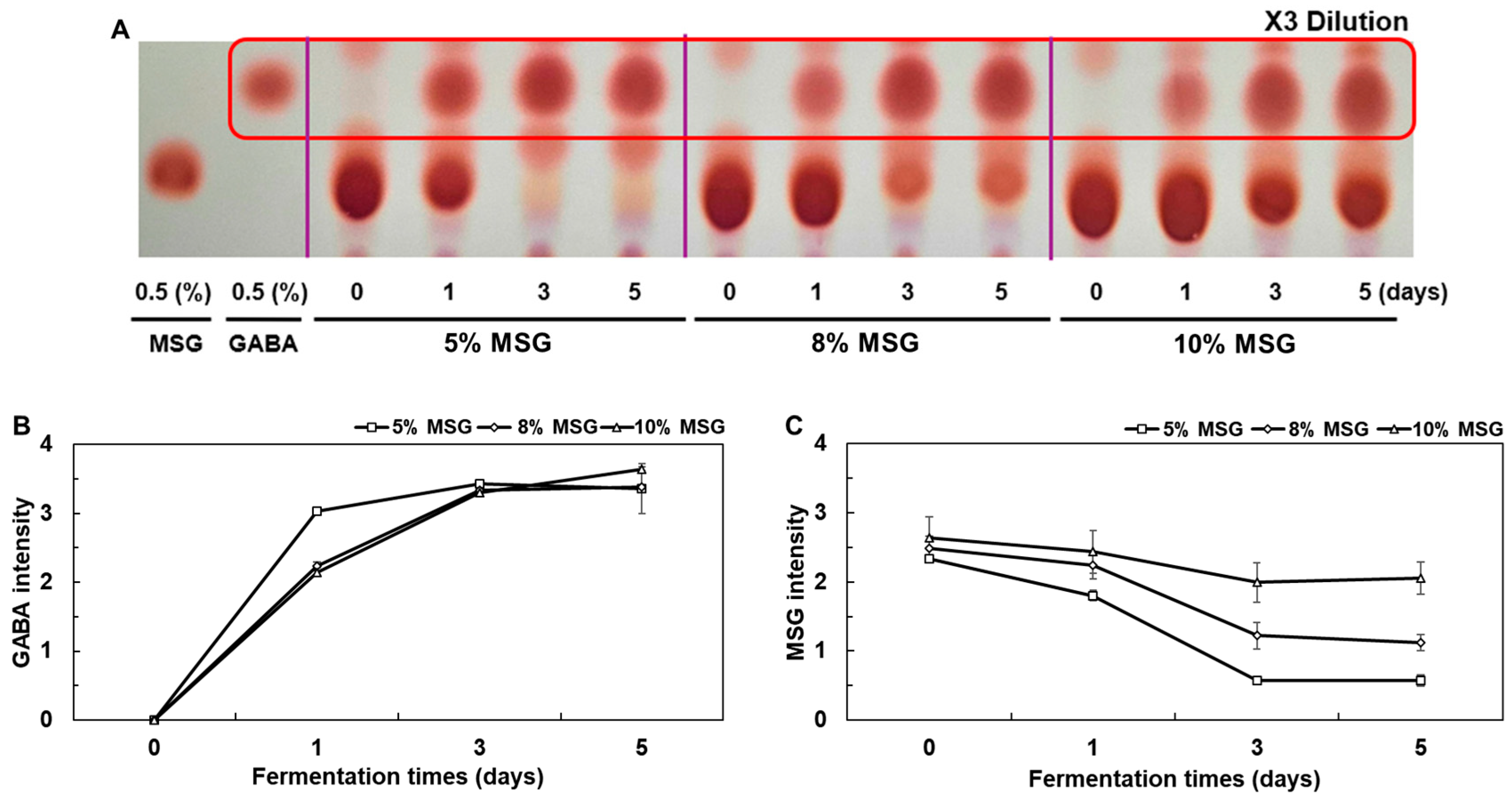
3.3. Analysis of Co-Fermented HSP Hydrolysate
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, Y.J.; Chung, S.; Hwang, J.T.; Shon, J.; Kim, E. A review of recent evidence of dietary protein intake and health. Nutr. Res. Pract. 2022, 16 (Suppl. S1), S37–S46. [Google Scholar] [CrossRef] [PubMed]
- Langyan, S.; Yadava, P.; Khan, F.N.; Dar, Z.A.; Singh, R.; Kumar, A. Sustaining protein nutrition through plant-based foods. Front. Nutr. 2022, 8, 772573. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Nawaz, M.A.; Buckow, R. Functional and food application of plant proteins—A review. Food Rev. Int. 2023, 39, 2428–2456. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, M.; Fan, H.; Wu, J. Emerging proteins as precursors of bioactive peptides/hydrolysates with health benefits. Curr. Opin. Food Sci. 2022, 48, 100914. [Google Scholar] [CrossRef]
- Liu, M.; Childs, M.; Loos, M.; Taylor, A.; Smart, L.B.; Abbaspourrad, A. The effects of germination on the composition and functional properties of hemp seed protein isolate. Food Hydrocoll. 2023, 134, 108085. [Google Scholar] [CrossRef]
- House, J.D.; Neufeld, J.; Leson, G. Evaluating the quality of protein from hemp seed (Cannabis sativa L.) products through the use of the protein digestibility-corrected amino acid score method. J. Agric. Food Chem. 2010, 58, 11801–11807. [Google Scholar] [CrossRef]
- Yano, H.; Fu, W. Hemp: A sustainable plant with high industrial value in food processing. Foods 2023, 12, 651. [Google Scholar] [CrossRef]
- Bhetwal, P.; Umar, M.; Anal, A.K. Enhanced functional characteristics and digestibility of blends of hemp protein hydrolysate and pea protein isolate. J. Food Meas. Charact. 2024, 18, 7112–7123. [Google Scholar] [CrossRef]
- Yoon, H.J.; Park, G.H.; Lee, Y.R.; Lee, J.M.; Ahn, H.L.; Lee, S.O. Enzymatic preparation and antioxidant activities of protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Sci. Preserv. 2023, 30, 434–445. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Rivero-Pino, F.; Villanueva, A.; Toscano-Sanchez, R.; Martin, M.E.; Millan, F.; Millan-Linares, M.C. Nutritional composition, ultrastructural characterization, and peptidome profile of antioxidant hemp protein hydrolysates. Food Biosci. 2023, 53, 102561. [Google Scholar] [CrossRef]
- Li, X.; He, Y.; Xie, Y.; Zhang, L.; Li, J.; Liu, H. Effects of fermentation with Kefir grains on nutrient composition, flavor volatiles, and product physical stability of a hemp seed (Cannabis sativa L.) beverage. Lebensm.-Wiss. Technol. 2023, 183, 114934. [Google Scholar] [CrossRef]
- Harwood, C.R.; Kikuchi, Y. The ins and outs of Bacillus proteases: Activities, functions and commercial significance. FEMS Microbiol. Rev. 2022, 46, fuab046. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, C.; Fang, H.; Zhang, D. Bacillus subtilis: A universal cell factory for industry, agriculture, biomaterials and medicine. Microb. Cell Factories 2020, 19, 173. [Google Scholar] [CrossRef]
- Lim, J.S.; Garcia, C.V.; Lee, S.P. Optimized production of GABA and γ-PGA in a turmeric and roasted soybean mixture co-fermented by Bacillus subtilis and Lactobacillus plantarum. Food Sci. Technol. Res. 2016, 22, 209–217. [Google Scholar] [CrossRef]
- Park, E.J.; Garcia, C.V.; Youn, S.J.; Park, C.D.; Lee, S.P. Fortification of γ-aminobutyric acid and bioactive compounds in Cucurbita moschata by novel two-step fermentation using Bacillus subtilis and Lactobacillus plantarum. Lebensm.-Wiss. Technol. 2019, 102, 22–29. [Google Scholar] [CrossRef]
- Durrani, D.; Idrees, R.; Idrees, H.; Ellahi, A. Vitamin B6: A new approach to lowering anxiety, and depression? Ann. Med. Surg. 2022, 82, 104663. [Google Scholar] [CrossRef]
- Sakashita, M.; Nakamura, U.; Horie, N.; Yokoyama, Y.; Kim, M.; Fujita, S. Oral supplementation using gamma-aminobutyric acid and whey protein improves whole body fat-free mass in men after resistance training. J. Clin. Med. Res. 2019, 11, 428. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, C.; Bouley, C.; Cayuela, C.; Bouttier, S.; Bourlioux, P.; Bellon-Fontaine, M.N. Cell surface characteristics of Lactobacillus casei subsp. casei, Lactobacillus paracasei subsp. paracasei, and Lactobacillus rhamnosus strains. Appl. Environ. Microbiol. 1997, 63, 1725–1731. [Google Scholar] [CrossRef]
- Park, K.B.; Oh, S.H. Cloning, sequencing and expression of a novel glutamate decarboxylase gene from a newly isolated lactic acid bacterium, Lactobacillus brevis OPK-3. Bioresour. Technol. 2007, 98, 312–319. [Google Scholar] [CrossRef]
- Zhu, N.; Wang, T.; Ge, L.; Li, Y.; Zhang, X.; Bao, H. γ-Amino butyric acid (GABA) synthesis enabled by copper-catalyzed carboamination of alkenes. Org. Lett. 2017, 19, 4718–4721. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.H. Production of GABA (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Heli, Z.; Hongyu, C.; Dapeng, B.; Yee Shin, T.; Yejun, Z.; Xi, Z.; Yingying, W. Recent advances of γ-aminobutyric acid: Physiological and immunity function, enrichment, and metabolic pathway. Front. Nutr. 2022, 9, 1076223. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Liu, Z.; Xie, F.; Bilal, M.; Liu, L.; Yang, R.; Wang, Z. Microbial production of gamma-aminobutyric acid: Applications, state-of-the-art achievements, and future perspectives. Crit. Rev. Biotechnol. 2021, 41, 491–512. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, Y.; Fukaya, T.; Kishino, S.; Ogawa, J. Production of GABA-enriched tomato juice by Lactiplantibacillus plantarum KB1253. J. Biosci. Bioeng. 2022, 134, 424–431. [Google Scholar] [CrossRef]
- Pakdeeto, A.; Phuengjayaem, S.; Arayakarn, T.; Phitchayaphon, C.; Tungkajiwangkoon, S.; Tanasupawat, S. Identification of gamma-aminobutyric acid (GABA)-producing lactic acid bacteria from plant-based Thai fermented foods and genome analysis of Lactobacillus brevis GPB7-4. ScienceAsia 2022, 48, 254–262. [Google Scholar] [CrossRef]
- An, Y.J.; Sung, N.J.; Lee, S.P. Production of highly enriched GABA through Lactobacillus plantarum fermentation of katsuobushi protein hydrolyzate made from Dendropanax morbiferus extract fermented by Bacillus subtilis. Food Sci. Preserv. 2023, 30, 146–154. [Google Scholar] [CrossRef]
- Yoon, W.K. Optimum Production of γ-PGA and GABA in Hovenia dulcis Extract by Co-Fermentation using Bacillus subtilis HA and Lactobacillus plantarum EJ2014. Master’s Thesis, Keimyung University, Daegu, Republic of Korea, 2019. [Google Scholar]
- Kang, H.M. Optimization of the Production for Poly-γ-Glutamic acid and γ-Aminobutyric Acid in Toxicodendron vernicifluum Extract by Co-Fermentation of Bacillus subtilis HA and Lactobacillus plantarum KS2020. Master’s Thesis, Keimyung University, Daegu, Republic of Korea, 2022. [Google Scholar]
- Lim, J.S.; Park, Y.H.; Jo, B.S.; Kim, J.E.; Lee, S.P. Increased production of γ-aminobutyric acid using the extract of Phellinus linteus fruiting body by Lactobacillus plantarum KS2020. Food Sci. Preserv. 2022, 29, 472–481. [Google Scholar] [CrossRef]
- Sun, B.; Zou, K.; Zhao, Y.; Tang, Y.; Zhang, F.; Chen, W.; Tang, X.; Chang, C.; Zheng, Y. The fermentation optimization for alkaline protease production by Bacillus subtilis BS-QR-052. Front. Microbiol. 2023, 14, 1301065. [Google Scholar] [CrossRef]
- Sun, X.; Sun, Y.; Li, Y.; Wu, Q.; Wang, L. Identification and characterization of the seed storage proteins and related genes of Cannabis sativa L. Front. Nutr. 2021, 8, 678421. [Google Scholar] [CrossRef]
- Karabulut, G.; Feng, H.; Yemiş, O. Physicochemical and antioxidant properties of industrial hemp seed protein isolate treated by high-intensity ultrasound. Plant Foods Hum. Nutr. 2022, 77, 577–583. [Google Scholar] [CrossRef]
- Jobgen, W.; Meininger, C.J.; Jobgen, S.C.; Li, P.; Lee, M.J.; Smith, S.B.; Spencer, T.E.; Fried, S.K.; Wu, G. Dietary L-arginine supplementation reduces white fat gain and enhances skeletal muscle and brown fat masses in diet-induced obese rats. J. Nutr. 2009, 139, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, S.H.; Phillips, S.M. Branched-chain amino acids (leucine, isoleucine, and valine) and skeletal muscle. In Nutrition and Skeletal Muscle; Academic Press: New York, NY, USA, 2019; pp. 283–298. [Google Scholar] [CrossRef]
- Gao, T.; Wong, Y.; Ng, C.; Ho, K. L-lactic acid production by Bacillus subtilis MUR1. Bioresour. Technol. 2012, 121, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Payot, T.; Chemaly, Z.; Fick, M. Lactic acid production by Bacillus coagulans—Kinetic studies and optimization of culture medium for batch and continuous fermentations. Enzym. Microb. Technol. 1999, 24, 191–199. [Google Scholar] [CrossRef]
- Ou, Q.; Zhao, J.; Sun, Y.; Zhao, Y.; Zhang, B. Utilization of lemon peel for the production of vinegar by a combination of alcoholic and acetic fermentations. Foods 2023, 12, 2488. [Google Scholar] [CrossRef] [PubMed]
- Rozes, N.; Peres, C. Effect of oleuropein and sodium chloride on viability and metabolism of Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 1996, 45, 839–843. [Google Scholar] [CrossRef]
- Shi, F.; Xie, Y.; Jiang, J.; Wang, N.; Li, Y.; Wang, X. Directed evolution and mutagenesis of glutamate decarboxylase from Lactobacillus brevis Lb85 to broaden the range of its activity toward a near-neutral pH. Enzym. Microb. Technol. 2014, 61, 35–43. [Google Scholar] [CrossRef]
- Kim, E.J.; Shin, H.J.; Lee, J.H. Developments of GABA process for the mass production. Korean Soc. Biotechnol. Bioeng. J. 2021, 36, 254–259. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, D.H.; Kang, H.J.; Shin, M.; Yang, S.Y.; Yang, J.; Jung, Y.H. Enhanced production of γ-aminobutyric acid (GABA) using Lactobacillus plantarum EJ2014 with simple medium composition. LWT 2021, 137, 110443. [Google Scholar] [CrossRef]
- Yao, C.; Liu, Y.; Hu, X.; Wang, X. Improve gamma-aminobutyric acid production in Corynebacterium glutamicum by optimizing the metabolic flux. Syst. Microbiol. Biomanuf. 2022, 2, 305–316. [Google Scholar] [CrossRef]
- Tan, S.W.; Zu Koh, Y.; Siva, S.; Wasoh, H.; Mohamed, M.S.; Sobri, Z.M.; Halim, M. Optimization of fermentative parameters to improve gamma-aminobutyric acid (GABA) production by Lactiplantibacillus plantarum B13. J. Biochem. Microbiol. Biotechnol. 2024, 12, 7–16. [Google Scholar] [CrossRef]
- Sassi, S.; Ilham, Z.; Jamaludin, N.S.; Halim-Lim, S.A.; Shin Yee, C.; Weng Loen, A.W.; Poh Suan, O.; Ibrahim, M.F.; Wan-Mohtar, W.A.A.Q.I. Critical optimized conditions for gamma-aminobutyric acid (GABA)-producing Tetragenococcus halophilus strain KBC from a commercial soy sauce Moromi in batch fermentation. Fermentation 2022, 8, 409. [Google Scholar] [CrossRef]
- Thongruck, K.; Maneerat, S. Enhanced production of gamma-aminobutyric acid (GABA) from Lactobacillus futsaii CS3 using agri-food industries by-products under batch and fed-batch fermentation. Indian J. Microbiol. 2023, 63, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Uehara, E.; Hokazono, H.; Tomonaga, A.; Fukagawa, M. Effects of γ-aminobutyric acid (GABA) intake in combination with exercise on muscle strength in humans with decreased mobility: A randomized, double-blind, placebo-controlled, parallel-group study. Funct. Foods Health Dis. 2024, 14, 248–269. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid (mg/100 g) | Con 0 h | H20 0 h | Con 24 h | H20 24 h |
|---|---|---|---|---|
| Asp | 2.01 ± 0.00 1) | 28.33 ± 0.29 | 5.83 ± 0.58 | 26.33 ± 0.29 |
| Glu | 87.83 ± 1.53 | 77.67 ± 1.12 | 6.00 ± 0.00 | 29.33 ± 1.16 |
| His | 38.24 ± 3.61 | 43.33 ± 0.29 | 12.00 ± 0.50 | 27.354 ± 0.29 |
| Arg | 3.00 ± 0.00 | 3.50 ± 0.00 | 39.02 ± 1.80 | 66.50 ± 0.29 |
| Lys | 13.14 ± 1.00 | 6.00 ± 0.00 | 28.00 ± 0.00 | 36.50 ± 0.00 |
| Phe | 30.17 ± 1.61 | 34.83 ± 0.76 | 14.83 ± 2.47 | 32.17 ± 1.53 |
| Tyr | 6.17 ± 0.58 | 2.83 ± 0.29 | 32.83 ± 1.04 | 37.00 ± 0.00 |
| Ala | 46.50 ± 0.00 | 46.03 ± 0.00 | 41.84 ± 0.29 | 45.00 ± 1.73 |
| Ile | 32.83 ± 0.76 | 35.83 ± 0.06 | 14.00 ± 0.00 | 37.00 ± 0.00 |
| Leu | 50.17 ± 0.29 | 58.83 ± 0.29 | 22.83 ± 1.26 | 68.17 ± 2.93 |
| Met | 18.00 ± 0.00 | 19.60 ± 0.29 | 12.17 ± 0.29 | 20.33 ± 0.29 |
| Val | 41.83 ± 0.76 | 45.50 ± 0.17 | 22.00 ± 0.00 | 42.67 ± 0.29 |
| Thr | 28.17 ± 0.29 | 19.50 ± 1.32 | 10.17 ± 1.61 | 22.6 + 7 ± 0.76 |
| Ser | 2.04 ± 0.00 | 9.67 ± 0.23 | 15.01 ± 2.18 | 42.33 ± 0.58 |
| Cys | 20.83 ± 1.76 | 0.50 ± 0.00 | 14.00 ± 0.00 | 3.00 ± 0.00 |
| Gly | 36.02 ± 0.00 | 28.67 ± 0.76 | 16.12 ± 1.32 | 17.67 ± 0.76 |
| Pro | 91.00 ± 1.80 | 123.7 ± 0.12 | 63.33 ± 0.17 | 84.50 ± 0.00 |
| Total | 547.50 ± 1.60 | 584.33 ± 1.62 | 372.83 ± 3.02 | 678.50 ± 1.48 |
| Co-Fermentation | GABA (mg/g) | Glutamic Acid (mg/g) |
|---|---|---|
| HSP hydrolysate added MSG 5% | 33.98 ± 1.24 1) | 14.05 ± 0.82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.-H.; Kim, J.-H.; Shin, D.-M.; Lee, S.-P. Hemp Seed Protein Hydrolysate Enriched with γ-Aminobutyric Acid and Peptides by Microbial Bioconversion. Fermentation 2024, 10, 483. https://doi.org/10.3390/fermentation10090483
Park Y-H, Kim J-H, Shin D-M, Lee S-P. Hemp Seed Protein Hydrolysate Enriched with γ-Aminobutyric Acid and Peptides by Microbial Bioconversion. Fermentation. 2024; 10(9):483. https://doi.org/10.3390/fermentation10090483
Chicago/Turabian StylePark, Yun-Ho, Joo-Hyeong Kim, Dong-Min Shin, and Sam-Pin Lee. 2024. "Hemp Seed Protein Hydrolysate Enriched with γ-Aminobutyric Acid and Peptides by Microbial Bioconversion" Fermentation 10, no. 9: 483. https://doi.org/10.3390/fermentation10090483
APA StylePark, Y.-H., Kim, J.-H., Shin, D.-M., & Lee, S.-P. (2024). Hemp Seed Protein Hydrolysate Enriched with γ-Aminobutyric Acid and Peptides by Microbial Bioconversion. Fermentation, 10(9), 483. https://doi.org/10.3390/fermentation10090483