Inhibition of Growth and Ammonia Production of Ruminal Hyper Ammonia-Producing Bacteria by Chinook or Galena Hops after Long-Term Storage

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
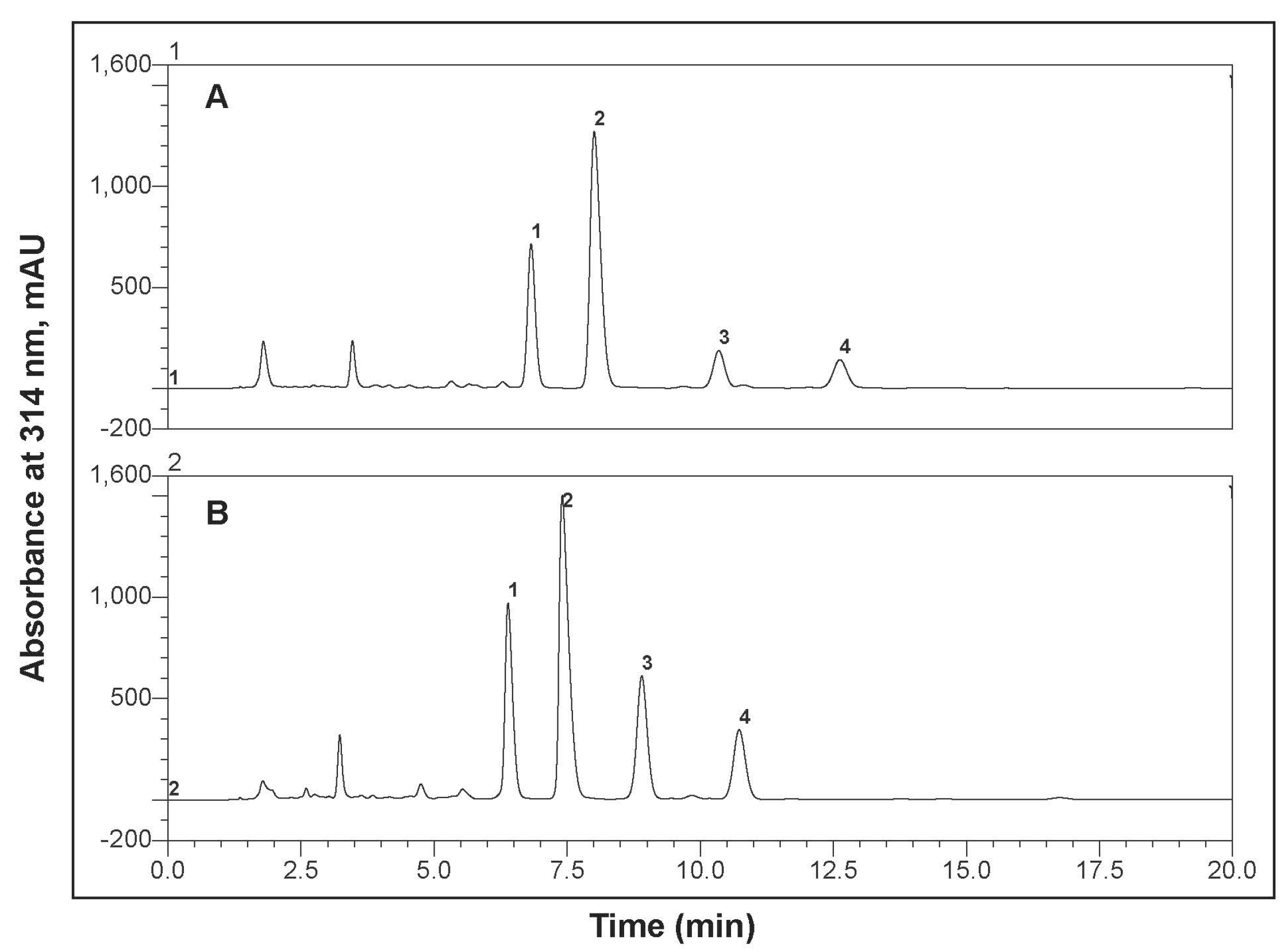
2.1. Hops and Bitter Acid Extraction
2.2. Bitter Acid Quantification
2.3. Medium Composition and Culture Conditions
2.4. Bacterial Growth Experiments
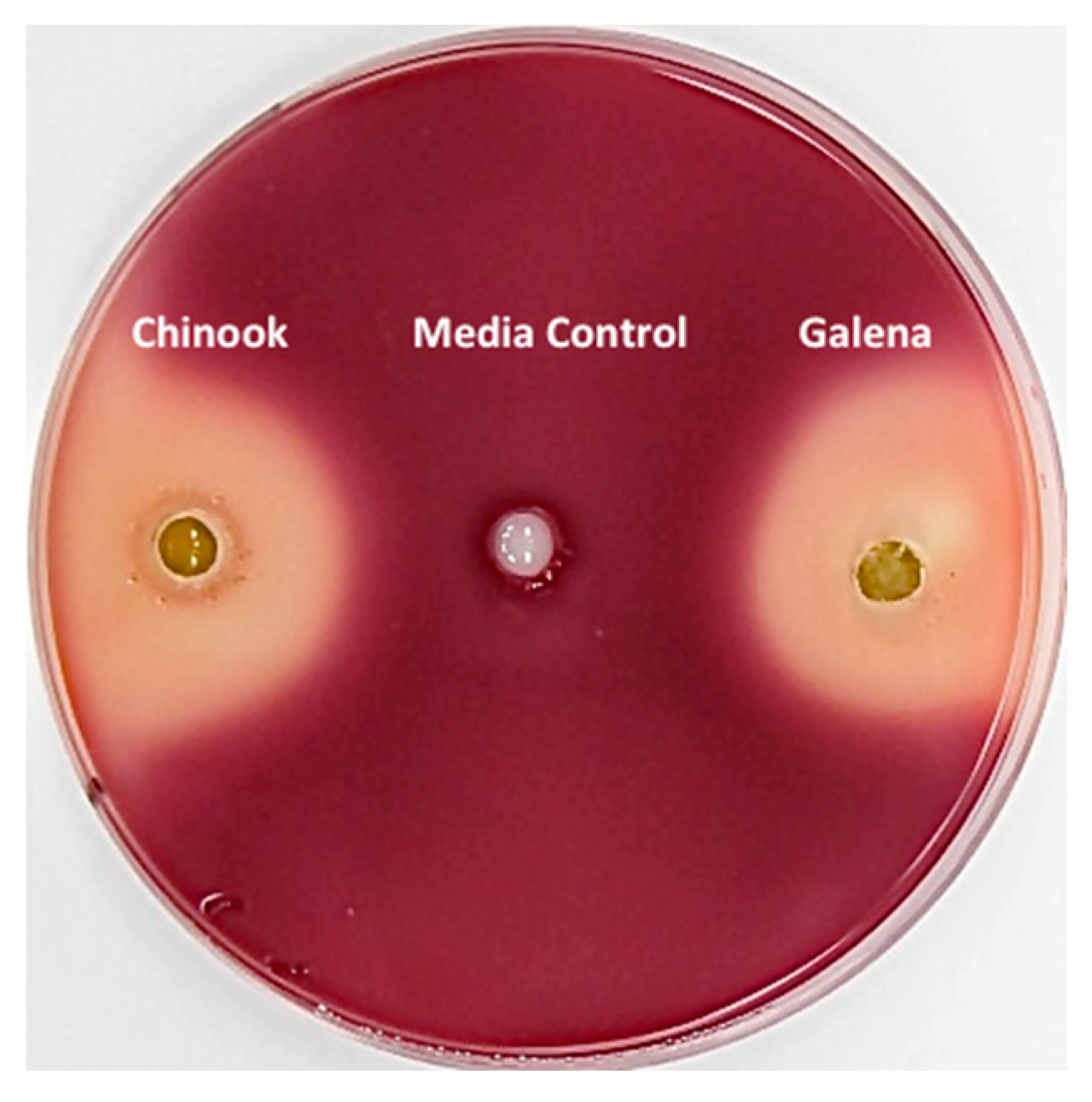
2.5. Petri Plate Bioassays
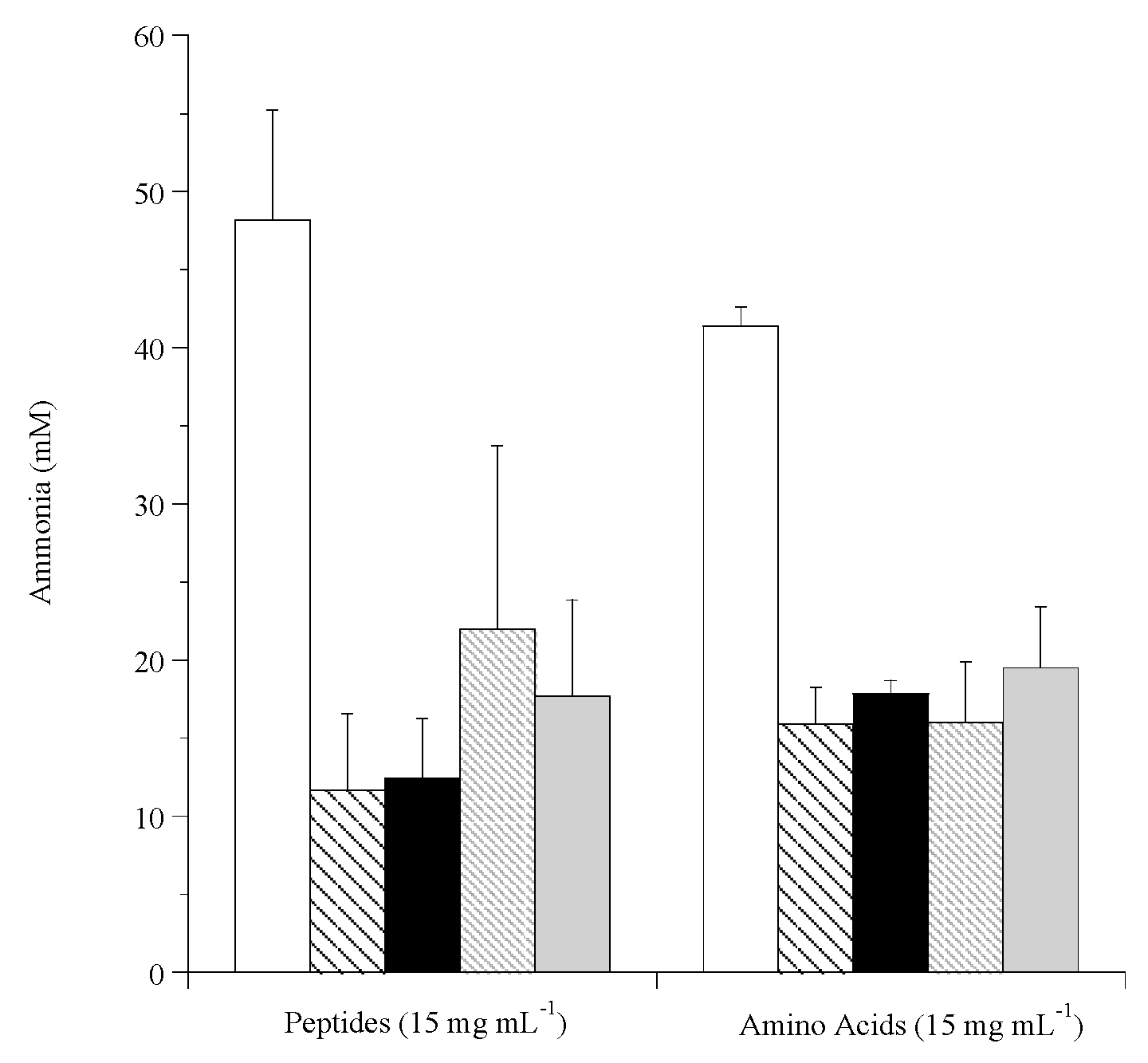
2.6. Rumen Microbe Suspensions
2.7. Replication and Statistics
3. Results
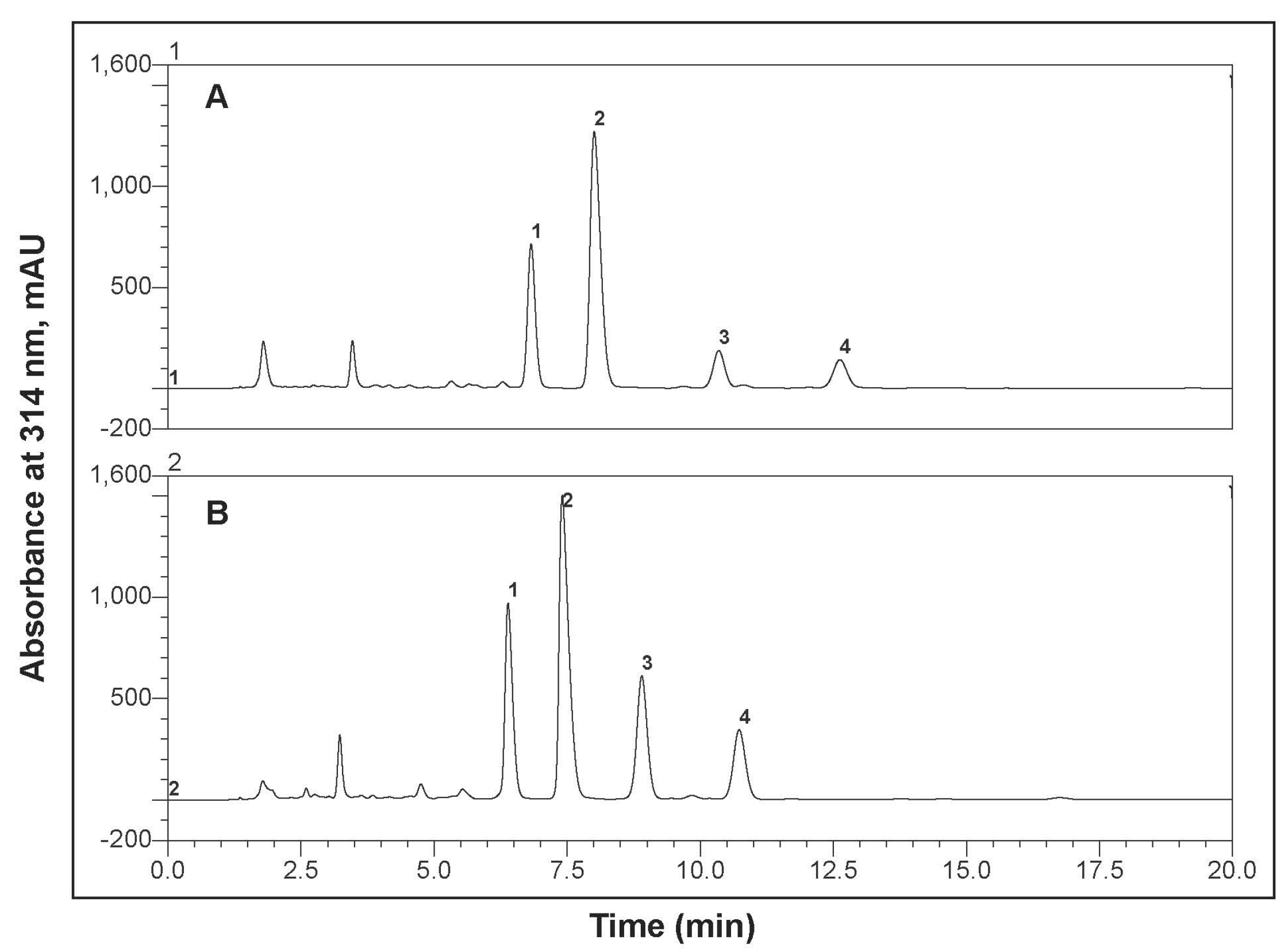
3.1. Bitter Acids in Stored Hop Pellets
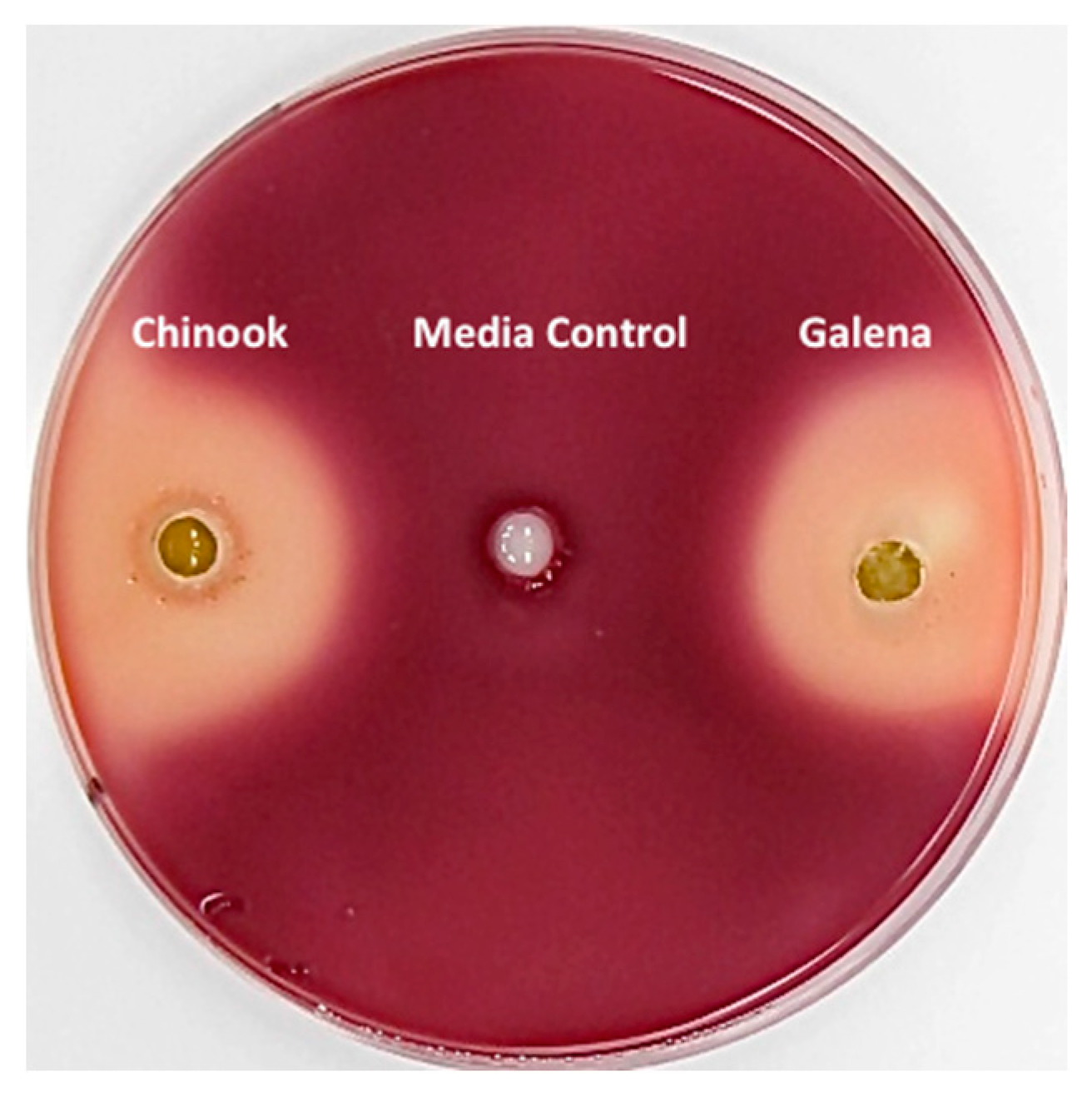
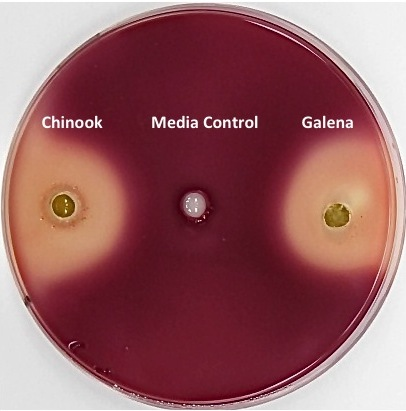
3.2. Inhibition of Clostridium sticklandii SR by Stored Hops
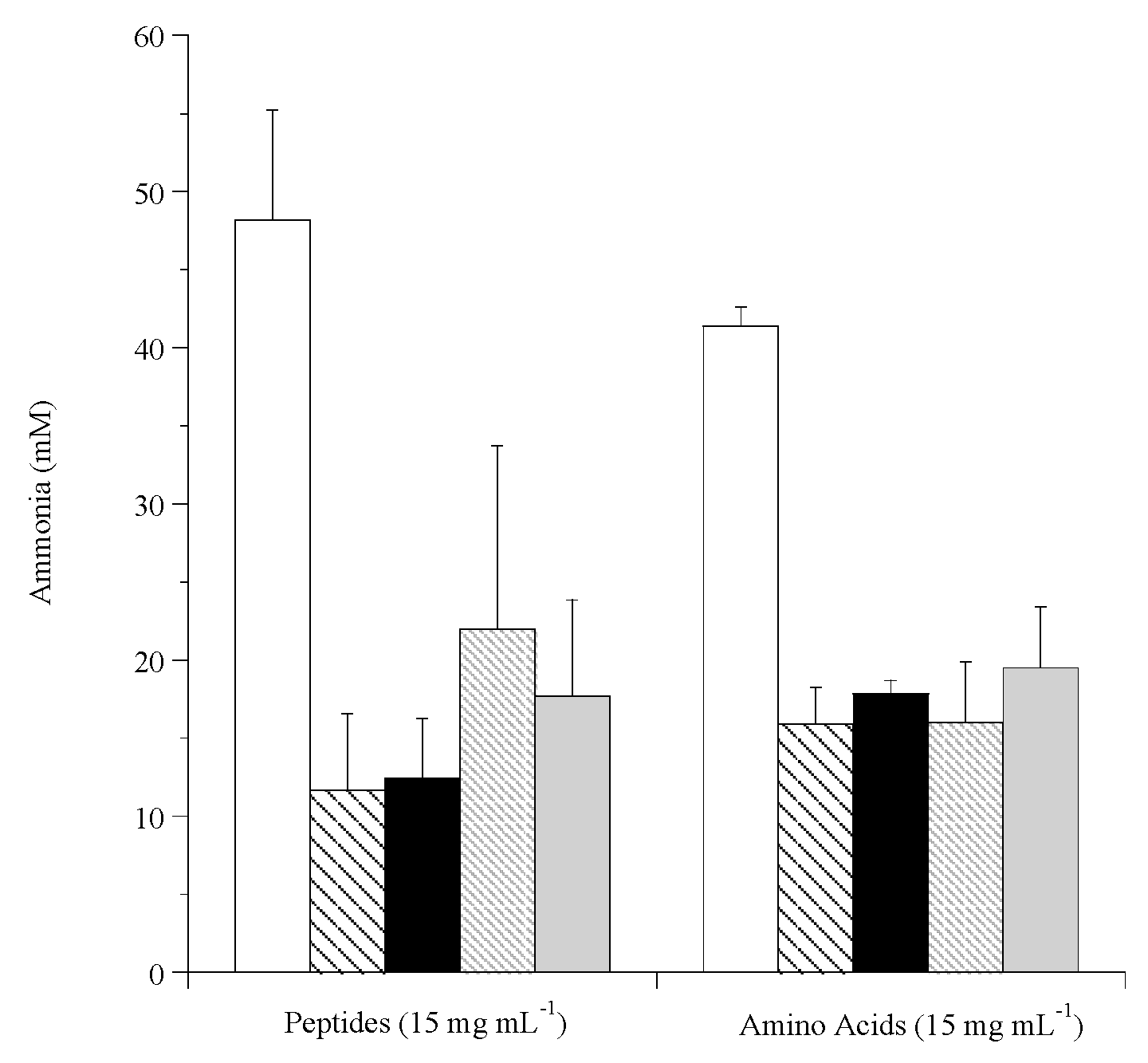
3.3. Effect of Stored Hops on Ammonia Production by Mixed Rumen Microorganisms
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- United States Agricultural Statistics Board, National Agricultural Statistics Service. Acreage; National Agricultural Statistics Service: Washington, DC, USA, 2017; ISBN 1949-1522. Available online: http://usda.mannlib.cornell.edu/usda/current/Acre/Acre-06-30-2017.pdf (accessed on 8 January 2017).
- Flythe, M.D.; Kagan, I.A.; Wang, Y.; Narvaez, N. Hops (Humulus lupulus L.) Bitter Acids: Modulation of Rumen Fermentation and Potential as an Alternative Growth Promoter. Front. Vet. Sci. 2017, 4, 131. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Strobel, H.J. Effect of ionophores on ruminal fermentation. Appl. Environ. Microbiol. 1989, 55, 1–6. [Google Scholar] [PubMed]
- Narvaez, N.; Wang, Y.; Xu, Z.; McAllister, T. Effects of hops on in vitro ruminal fermentation of diets varying in forage content. Livest. Sci. 2011, 138, 193–201. [Google Scholar] [CrossRef]
- Flythe, M.D.; Aiken, G.E. Effects of hops (Humulus lupulus L.) extract on volatile fatty acid production by rumen bacteria. J. Appl. Microbiol. 2010, 109, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Lavrenčič, A.; Levart, A.; Košir, I.J.; Čerenak, A. Influence of two hop (Humulus lupulus L.) varieties on in vitro dry matter and crude protein degradability and digestibility in ruminants. J. Sci. Food Agric. 2013, 94, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Flythe, M.D. The antimicrobial effects of hops (Humulus lupulus L.) on ruminal hyper ammonia-producing bacteria. Lett. Appl. Microbiol. 2009, 118, 242–248. [Google Scholar]
- Sahlstrom, A.; Rostad, S. HPLC determination of α and β acids in hops. Concordia Coll. J. Anal. Chem. 2011, 2, 78–83. [Google Scholar]
- Pedjie, N. Determination of α Acids in Hops and Beers; Application Note; Perkin Elmer: Waltham, MA, USA, 2012. [Google Scholar]
- Chen, G.J.; Russell, J.B. More monensin-sensitive, ammonia-producing bacteria from the rumen. Appl. Environ. Microbiol. 1989, 55, 1052–1057. [Google Scholar] [PubMed]
- Russell, J.B.; Strobel, H.J.; Chen, G.J. Enrichment and isolation of a ruminal bacterium with a very high specific activity of ammonia production. Appl. Environ. Microbiol. 1988, 54, 872–877. [Google Scholar] [PubMed]
- Paster, B.J.; Russell, J.B.; Yang, C.M.; Chow, J.M.; Woese, C.R.; Tanner, R. Phylogeny of the ammonia-producing ruminal bacteria Peptostreptococcus anaerobius, Clostridium sticklandii, and Clostridium aminophilum sp. nov. Int. J. Syst. Bacteriol. 1993, 43, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Kagan, I.A.; Flythe, M.D. Thin-Layer Chromatographic (TLC) separations and bioassays of plant extracts to identify antimicrobial compounds. J. Vis. Exp. 2014, 85, e51411. [Google Scholar] [CrossRef] [PubMed]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [PubMed]
- Lowry, O.H.; Rosenbough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Dresel, M.; Vogt, C.; Dunkel, A.; Hofmann, T. The Bitter Chemodiversity of Hops (Humulus lupulus L.). J. Agric. Food Chem. 2016, 64, 7789–7799. [Google Scholar] [CrossRef] [PubMed]
- Teuber, M.; Schmalreck, A.F. Membrane Leakage in Bacillus subtilis 169 induced by the hop constituents lupulone, humulone, isohumulone and humulinic acid. Arch. Microbiol. 1973, 94, 159–171. [Google Scholar]
- Bento, C.B.P.; De Azevedo, A.C.; Detmann, E.; Mantovani, H.C. Biochemical and genetic diversity of carbohydrate-fermenting and obligate amino acid-fermenting hyper-ammonia-producing bacteria from Nellore steers fed tropical forages and supplemented with casein. BMC Microbiol. 2015, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Eschenlauer, S.C.; McKain, N.; Walker, N.D.; McEwan, N.R.; Newbold, C.J.; Wallace, R.J. Ammonia production by ruminal microorganisms and enumeration, isolation, and characterization of bacteria capable of growth on peptides and amino acids from the sheep rumen. Appl. Environ. Microbiol. 2002, 68, 4925–4931. [Google Scholar] [CrossRef] [PubMed]
- Flythe, M.; Andries, K. The effects of monensin on amino acid catabolizing bacteria isolated from the Boer goat rumen. Small Rumin. Res. 2009, 81, 178–181. [Google Scholar] [CrossRef]
- Flythe, M.D.; Aiken, G.E.; Gellin, G.L.; Klotz, J.L.; Goff, B.M.; Andries, K.M. Hops (Humulus lupulus) ß-Acid as an Inhibitor of Caprine Rumen Hyper-Ammonia-Producing Bacteria In Vitro. Agric. Food Anal. Bacteriol. 2015, 5, 29–36. [Google Scholar]
- Krause, D.O.; Nagaraja, T.G.; Wright, A.D.G.; Callaway, T.R. Board-invited review: Rumen microbiology: Leading the way in microbial ecology. J. Anim. Sci. 2013, 91, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Pszczolkowski, V.L.; Bryant, R.W.; Harlow, B.E.; Aiken, G.E.; Martin, L.J.; Flythe, M.D. Effects of spent craft brewers’ yeast on fermentation and methane production by rumen microorganisms. Adv. Microbiol. 2016, 6, 716–723. [Google Scholar] [CrossRef]
- Harlow, B.E.; Bryant, R.W.; Cohen, S.D.; O’Connell, S.P.; Flythe, M.D. Degradation of spent craft brewer’s yeast by caprine rumen hyper ammonia-producing bacteria. Lett. Appl. Microbiol. 2016, 63, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Siragusa, G.R.; Haas, G.J.; Matthews, P.D.; Smith, R.J.; Buhr, R.J.; Dale, N.M.; Wise, M.G. Antimicrobial activity of lupulone against Clostridium perfringens in the chicken intestinal tract jejunum and caecum. J. Antimicrob. Chemother. 2008, 61, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Harlow, B.E.; Lawrence, L.M.; Kagan, I.A.; Flythe, M.D. Inhibition of fructan-fermenting equine faecal bacteria and Streptococcus bovis by hops (Humulus lupulus L.) β-acid. J. Appl. Microbiol. 2014, 117, 329–339. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bitter Acid | Chinook | Galena | ||||
|---|---|---|---|---|---|---|
| 2012 (Label) a | 2017 (Equation) b | 2017 (Standard Curve) c | 2012 (Label) | 2017 (Equation) | 2017 (Standard Curve) | |
| α | 11.4% | 12.9% | 12.3% | 13.9% | 13.3% | 13.4% |
| β | 3.2% | 3.6% | 3.4% | 9.4% | 8.5% | 8.6% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flythe, M.D.; Harlow, B.E.; Aiken, G.E.; Gellin, G.L.; Kagan, I.A.; Pappas, J. Inhibition of Growth and Ammonia Production of Ruminal Hyper Ammonia-Producing Bacteria by Chinook or Galena Hops after Long-Term Storage. Fermentation 2017, 3, 68. https://doi.org/10.3390/fermentation3040068
Flythe MD, Harlow BE, Aiken GE, Gellin GL, Kagan IA, Pappas J. Inhibition of Growth and Ammonia Production of Ruminal Hyper Ammonia-Producing Bacteria by Chinook or Galena Hops after Long-Term Storage. Fermentation. 2017; 3(4):68. https://doi.org/10.3390/fermentation3040068
Chicago/Turabian StyleFlythe, Michael D., Brittany E. Harlow, Glen E. Aiken, Gloria L. Gellin, Isabelle A. Kagan, and Jesse Pappas. 2017. "Inhibition of Growth and Ammonia Production of Ruminal Hyper Ammonia-Producing Bacteria by Chinook or Galena Hops after Long-Term Storage" Fermentation 3, no. 4: 68. https://doi.org/10.3390/fermentation3040068
APA StyleFlythe, M. D., Harlow, B. E., Aiken, G. E., Gellin, G. L., Kagan, I. A., & Pappas, J. (2017). Inhibition of Growth and Ammonia Production of Ruminal Hyper Ammonia-Producing Bacteria by Chinook or Galena Hops after Long-Term Storage. Fermentation, 3(4), 68. https://doi.org/10.3390/fermentation3040068