Isolation and Selection of Non-Saccharomyces Yeasts Being Capable of Degrading Citric acid and Evaluation Its Effect on Kiwifruit Wine Fermentation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Media
2.2. Fruits and Soil Used for Isolation of Potential Yeast Strains to Degrade Citric Acid
2.3. Preliminary Screening and Isolation of Yeasts
2.4. Further Screening of Selected Yeasts
2.5. Yeast Identification
2.6. Stress Resistance Analysis
2.7. Kiwifruit Juice Fermentation Experiment
2.7.1. Kiwifruit Juice Pre-Treatment
2.7.2. Fermentation Deacidification Experiment
2.8. Quality Analysis
2.8.1. Determination of Total Acidity, Total Sugars, pH and SSC (Soluble Solids Content)
2.8.2. Determination of Organic Acid
2.8.3. Determination of Total Flavonoids and Total Phenolics
2.8.4. Determination of Ethanol
2.8.5. Determination of Amino Acid Content
2.8.6. Determination of Volatile Compounds
2.9. Statistical Analysis
3. Results
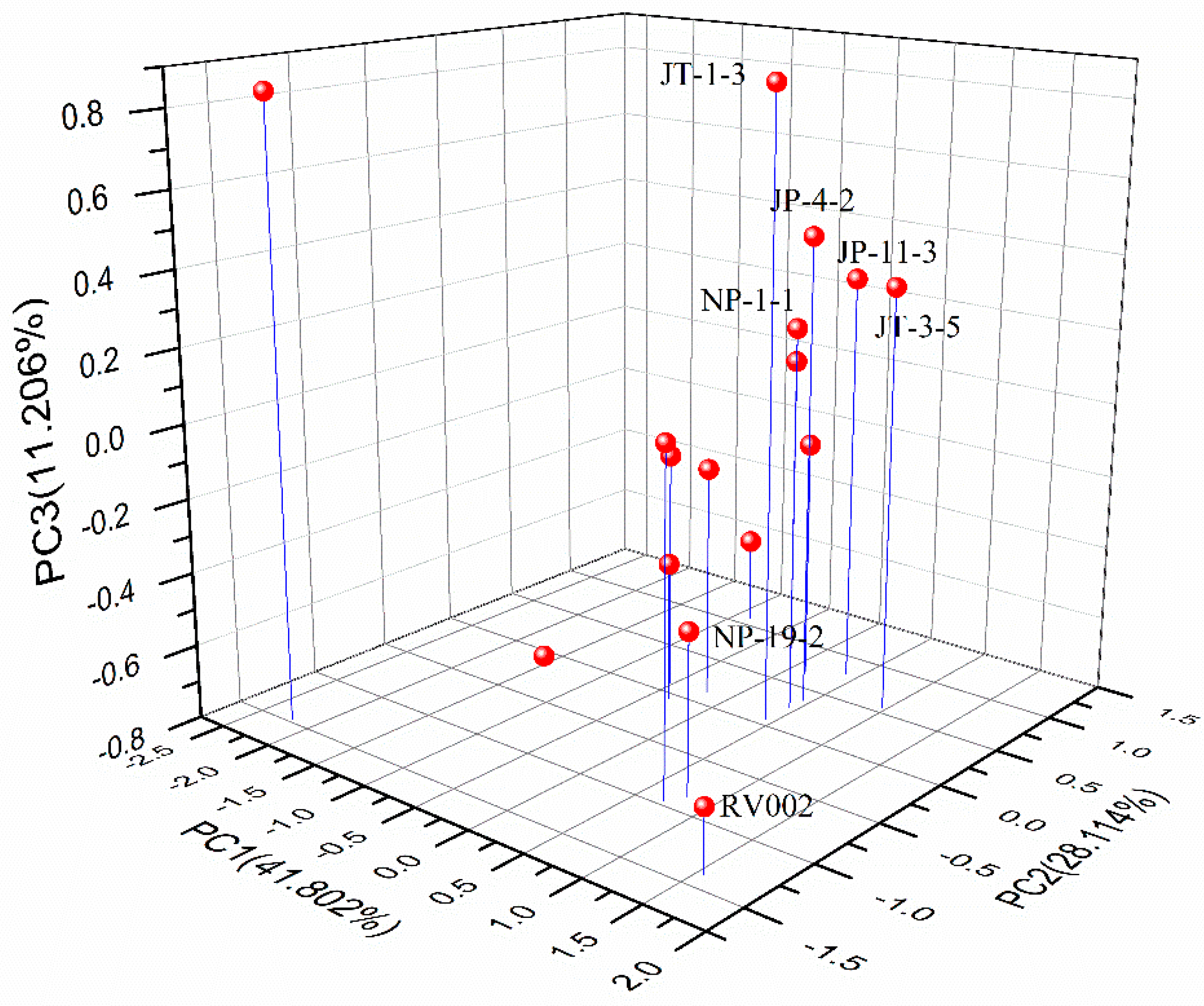
3.1. Isolation and Screening of Deacidification Yeast
3.2. Stress Resistance Analysis
3.3. Analytical Profiles of Kiwifruit Juice and Wine
3.3.1. Changes in Properties in Kiwifruit Juice and Wine
3.3.2. Changes in Amino Acids Profiles in Kiwifruit Juice and Wine
3.3.3. Changes in Volatile Components
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tavarini, S.; Degl’Innocenti, E.; Remorini, D.; Massai, R.; Guidi, L. Antioxidant capacity, ascorbic acid, total phenols and carotenoids changes during harvest and after storage of Hayward kiwifruit. Food Chem. 2008, 107, 282–288. [Google Scholar] [CrossRef]
- Du, G.; Li, M.; Ma, F.; Dong, L. Antioxidant capacity and the relationship with polyphenol and Vitamin C in Actinidia fruits. Food Chem. 2009, 113, 557–562. [Google Scholar] [CrossRef]
- Soufleros, E.H.; Pissa, I.; Petridis, D.; Lygerakis, M.; Mermelas, K.; Boukouvalas, G.; Tsimitakis, E. Instrumental analysis of volatile and other compounds of Greek kiwi wine; sensory evaluation and optimisation of its composition. Food Chem. 2001, 75, 487–500. [Google Scholar] [CrossRef]
- Chen, A.-J.; Fu, Y.-Y.; Jiang, C.; Zhao, J.-L.; Liu, X.-P.; Liu, L.; Ma, J.; Liu, X.-Y.; Shen, G.-H.; Li, M.-L.; et al. Effect of mixed fermentation (Jiuqu and Saccharomyces cerevisiae EC1118) on the quality improvement of kiwi wine. CyTA J. Food 2019, 17, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Maja, M.P.; Valentina, S.; Ana, S.; Franci, S.; Robert, V. Composition of sugars, organic acids, and total phenolics in 25 wild or cultivated berry species. J. Food Sci. 2012, 77, C1064–C1070. [Google Scholar]
- Ma, T.; Sun, X.; Zhao, J.; You, Y.; Lei, Y.; Gao, G.; Zhan, J. Nutrient compositions and antioxidant capacity of kiwifruit (Actinidia) and their relationship with flesh color and commercial value. Food Chem. 2017, 218, 294–304. [Google Scholar] [CrossRef]
- Matsumoto, S.; Obara, T.; Luh, B.S. Changes in chemical constituents of kiwifruit during post-harvest ripening. J. Food Sci. 1983, 48, 607–611. [Google Scholar] [CrossRef]
- Macrae, E.A.; Stec, M.G.H.; Triggs, C.M. Effects of postharvest treatment on the sensory qualities of kiwifruit harvested at different maturities. J. Sci. Food Agric. 2010, 50, 533–546. [Google Scholar] [CrossRef]
- Walton, E.F.; Jong, T.M.D. Growth and compositional changes in kiwifruit berries from three Californian locations. Ann. Bot. 1990, 66, 285–298. [Google Scholar] [CrossRef]
- Nagai, R.; Nagai, M.; Shimasaki, S.; Baynes, J.W.; Fujiwara, Y. Citric acid inhibits development of cataracts, proteinuria and ketosis in streptozotocin (Type 1) diabetic rats. Biochem. Biophys. Res. Commun. 2010, 393, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Penniston, K.L.; Nakada, S.Y.; Holmes, R.P.; Assimos, D.G. Quantitative assessment of citric acid in lemon juice, lime juice, and commercially-available fruit juice products. J. Endourol. 2008, 22, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Ryall, R.L. Urinary inhibitors of calcium oxalate crystallization and their potential role in stone formation. World J. Urol. 1997, 15, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Tanner, G.A.; Vijayalakshmi, K.; Tanner, J.A. Effects of potassium citrate/citric acid intake in a mouse model of polycystic kidney disease. Nephron 2000, 84, 270–273. [Google Scholar] [CrossRef]
- Lan, W.; Lan, W.; Chan, C.; Hsieh, C.; Chang, M.; Jeng, J. The effects of extracellular citric acid acidosis on the viability, cellular adhesion capacity and protein synthesis of cultured human gingival fibroblasts. Aust. Dent. J. 1999, 44, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Rapp, A.; Versini, G. Influence of nitrogen compounds in grapes on aroma compounds of wines. In Developments in Food Science; Charalambous, G., Ed.; Elsevier: Amsterdam, The Netherlands, 1995; Volume 37, pp. 1659–1694. [Google Scholar]
- Abraham, A.O.; Laing, M.D.; Bower, J.P. Isolation and in vivo screening of yeast and Bacillus antagonists for the control of Penicillium digitatum of citrus fruit. Biol. Control 2010, 53, 32–38. [Google Scholar] [CrossRef]
- Wen, L.K.; Wang, L.F.; Wang, G.Z. Degradation of L-malic and critic acids by issatchenkia terricola. Food Sci. 2011, 32, 220–223. [Google Scholar]
- Wang, L.F.; Zhang, W.; Wen, L.K. Screening and Identification of a strain degrading L-malic acid and critic acid. Food Sci. 2010, 31, 279–282. [Google Scholar]
- Zhao, Y.P. Screening yeast degrading citric acid in hawthorn fruit juice and its degrading characteristics. Acta Microbiol. Sin. 2004, 44, 235–239. [Google Scholar]
- Casal, M.; Paiva, S.; Queirós, O.; Soares-Silva, I. Transport of carboxylic acids in yeasts. FEMS Microbiol. Rev. 2008, 32, 974–994. [Google Scholar] [CrossRef] [Green Version]
- Cássio, F.; Leao, C. Low-and high-affinity transport systems for citric acid in the yeast Candida utilis. Appl. Environ. Microbiol. 1991, 57, 3623–3628. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, I.; Fukuda, T.; Shimohashi, A.; Oota, T. Sugar and organic acid composition in the fruit juice of different actinidia varieties. Food Sci. Technol. Int. Tokyo 2008, 14, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Barnett, J.A.; Payne, R.W.; Yarrow, D. Yeasts: Characteristics and Identification, 3rd ed.; Cambridge University Press: Cambridge, UK, 2001; Volume 18, p. 547. [Google Scholar]
- Edwards, C.G. Illustrated Guide to Microbes and Sediments in Wine, Beer and Juice; Gusmer Enterprises: Mountainside, NJ, USA, 2005. [Google Scholar]
- Zhong, W.; Li, X.; Yang, H.; Li, E. A novel, effective, and feasible method for deacidifying kiwifruit wine by weakly basic ion exchange resins. J. Food Process Eng. 2018, 42, e12969. [Google Scholar] [CrossRef]
- Mu, Z.; Yang, X.; Yuan, H. Detection and identification of wild yeast in Koumiss. Food Microbiol. 2012, 31, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, P.; Querol, A.; del Olmo, M. Analysis of the stress resistance of commercial wine yeast strains. Arch. Microbiol. 2001, 175, 450–457. [Google Scholar] [CrossRef]
- Hu, X.H.; Wang, M.H.; Tan, T.; Li, J.R.; Yang, H.; Leach, L.; Zhang, R.M.; Luo, Z.W. Genetic dissection of ethanol tolerance in the budding yeast saccharomyces cerevisiae. Genetics 2007, 175, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Tao, C.; Zhu, B.; Bai, W.; Zhang, L.; Wang, Z.; Liang, X. Identification of Zygosaccharomyces mellis strains in stored honey and their stress tolerance. Food Sci. Biotechnol. 2016, 25, 1645–1650. [Google Scholar] [CrossRef]
- Egydio, A.P.M.; Catarina, C.S.; Floh, E.I.S.; Santos, D.Y.A.C.D. Free amino acid composition of Annona (Annonaceae) fruit species of economic interest. Ind. Crop. Prod. 2013, 45, 373–376. [Google Scholar] [CrossRef]
- Pak, C. Citrate and renal calculi: An update. Miner. Electrolyte Metab. 1994, 20, 371–377. [Google Scholar]
- Clemente-Jimenez, J.M.; Mingorance-Cazorla, L.; Martínez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Influence of sequential yeast mixtures on wine fermentation. Int. J. Food Microbiol. 2005, 98, 301–308. [Google Scholar] [CrossRef]
- Hallsworth, J.E. Ethanol-induced water stress in yeast. J. Ferment. Bioeng. 1998, 85, 125–137. [Google Scholar] [CrossRef]
- Pina, C.; Santos, C.; Couto, J.A.; Hogg, T. Ethanol tolerance of five non-Saccharomyces wine yeasts in comparison with a strain of Saccharomyces cerevisiae—Influence of different culture conditions. Food Microbiol. 2004, 21, 439–447. [Google Scholar] [CrossRef]
- Gil, J.; Mateo, J.; Jiménez, M.; Pastor, A.; Huerta, T. Aroma compounds in wine as influenced by apiculate yeasts. J. Food Sci. 1996, 61, 1247–1250. [Google Scholar] [CrossRef]
- Bauer, F.; Pretorius, I.S. Yeast stress response and fermentation efficiency: How to survive the making of wine-a review. S. Afr. J. Enol. Vitic. 2000, 21, 27–51. [Google Scholar] [CrossRef]
- Sivakumaran, S.; Huffman, L.; Sivakumaran, S.; Drummond, L. The nutritional composition of Zespri® SunGold Kiwifruit and Zespri® Sweet Green Kiwifruit. Food Chem. 2018, 238, 195–202. [Google Scholar] [CrossRef]
- Mingorance-Cazorla, L.; Clemente-Jiménez, J.; Martínez-Rodríguez, S.; Las Heras-Vázquez, F.; Rodríguez-Vico, F. Contribution of different natural yeasts to the aroma of two alcoholic beverages. World J. Microbiol. Biotechnol. 2003, 19, 297–304. [Google Scholar] [CrossRef]
- Anfang, N. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2010, 15, 1–8. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
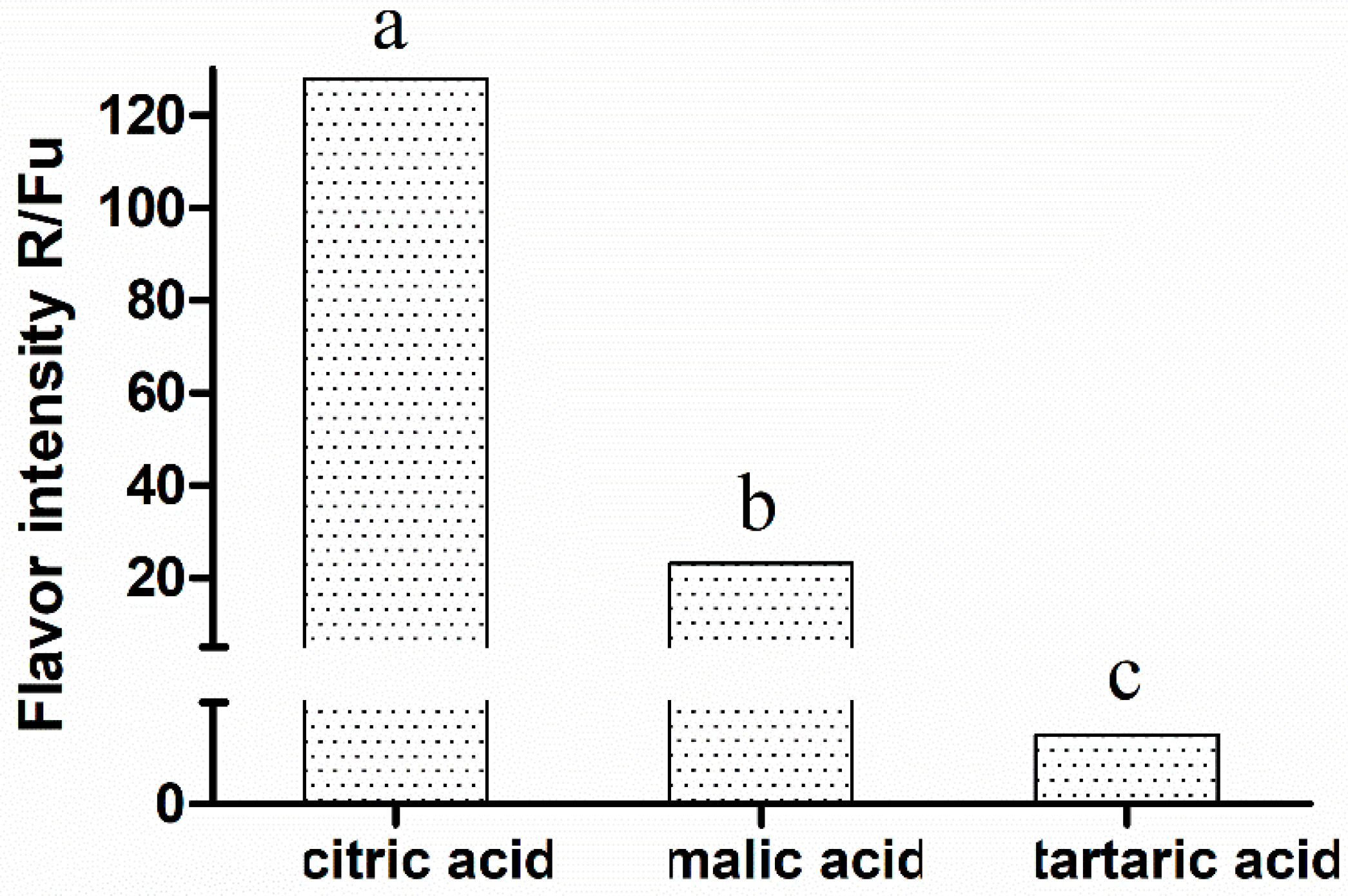
- Neta, E.R.D.; Johanningsmeier, S.D.; Drake, M.A.; McFeeters, R.F. Effects of pH Adjustment and Sodium Ions on Sour Taste Intensity of Organic Acids. J. Food Sci. 2009, 74, S165–S169. [Google Scholar] [CrossRef]
- Chtourou, M.; Ammar, E.; Nasri, M.; Medhioub, K. Isolation of a yeast, Trichosporon cutaneum, able to use low molecular weight phenolic compounds: Application to olive mill waste water treatment. J. Chem. Technol. Biotechnol. 2004, 79, 869–878. [Google Scholar] [CrossRef]
- Adav, S.S.; Chen, M.Y.; Lee, D.J.; Ren, N.Q. Degradation of phenol by aerobic granules and isolated yeast Candida tropicalis. Biotechnol. Bioeng. 2007, 96, 844–852. [Google Scholar] [CrossRef]
- Li, W.-Q.; Hu, Q.-P.; Xu, J.-G. Changes in physicochemical characteristics and free amino acids of hawthorn (Crataegus pinnatifida) fruits during maturation. Food Chem. 2015, 175, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Sola, A.; Maaheimo, H.; Ylonen, K.; Ferrer, P.; Szyperski, T. Amino acid biosynthesis and metabolic flux profiling of Pichia pastoris. Eur. J. Biochem. 2004, 271, 2462–2470. [Google Scholar] [CrossRef] [PubMed]
- Viana, F.; Gil, J.V.; Genovés, S.; Vallés, S.; Manzanares, P. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Shiota, H. Kiwifruit. Koryo 1982, 137, 59–64. [Google Scholar]
- Young, H.; Perera, C.O.; Paterson, V.J. Identification of E-Hex-3-enal as an important contributor to the off-flavour aroma in kiwifruit juice. J. Sci. Food Agric. 2010, 58, 519–522. [Google Scholar] [CrossRef]
- Young, H.; Paterson, V.J.; Burns, D.J.W. Volatile aroma constituents of kiwifruit. J. Sci. Food Agric. 2010, 34, 81–85. [Google Scholar] [CrossRef]
- Garcia, C.V.; Quek, S.-Y.; Stevenson, R.J.; Winz, R.A. Kiwifruit flavour: A review. Trends Food Sci. Technol. 2012, 24, 82–91. [Google Scholar] [CrossRef]
- Paterson, V.J.; Macrae, E.A.; Young, H. Relationships between sensory properties and chemical composition of kiwifruit (Actinidia deliciosa). J. Sci. Food Agric. 1991, 57, 235–251. [Google Scholar] [CrossRef]
- Li, X.; Xing, Y.; Cao, L.; Xu, Q.; Li, S.; Wang, R.; Jiang, Z.; Che, Z.; Lin, H. Effects of six commercial saccharomyces cerevisiae strains on phenolic attributes, antioxidant activity, and aroma of kiwifruit (Actinidia deliciosa cv.) wine. BioMed Res. Int. 2017, 2017, 2934743. [Google Scholar] [CrossRef] [Green Version]
- Powers, J.J.; Howell, A.; Lillard, D.; Vacinek, S. Effect of temperature on threshold values for citric acid, malic acid and quinine sulphate—Energy of activation and extreme-value determination. J. Sci. Food Agric. 1971, 22, 543–547. [Google Scholar] [CrossRef]
- Bo, W. Effect of Different Yeasts on Organic Acids in Blueberry Wine; Shenyang Agriculture University: Shenyang, China, 2018. (In Chinese) [Google Scholar]
- Stewart, J.E.; Feinle-Bisset, C.; Golding, M.; Delahunty, C.; Clifton, P.M.; Keast, R.S.J. Oral sensitivity to fatty acids, food consumption and BMI in human subjects. Br. J. Nutr. 2010, 104, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, Y.; Endo, S.; Sakai, F.; Yoshimura, I. A new whole-mouth gustatory test procedure: I. Thresholds and principal components analysis in healthy men and women. Acta Oto-Laryngol. 2002, 122, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Tempere, S.; Cuzange, E.; Malak, J.; Bougeant, J.C.; de Revel, G.; Sicard, G. The training level of experts influences their detection thresholds for key wine compounds. Chemosens. Percept. 2011, 4, 99–115. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Strain Number | Putative Species | Residual Citric Acid Concentration (g/L) | GenBank No. |
|---|---|---|---|
| LJD-7-2 | Candida natalensis | 0.13 ± 0.10b | MN736495 |
| NP-19-2 | Candida xylopsoci | 0.16 ± 0.02b | MN736497 |
| JP-4-2 | Meyerozyma guilliermondii | 0.17 ± 0.13b | MN736499 |
| NP-1-1 | Meyerozymaguilliermondii | 0.18 ± 0.21b | MN736500 |
| JST-18-3 | Pichia guilliermondii | 0.20 ± 0.05b | MN653214 |
| JT-1-3 | Pichia fermentans | 0.22 ± 0.34b | MN314412 |
| NP-5-5 | Meyerozyma guilliermondii | 0.24 ± 0.21b | MN736501 |
| JT-3-5 | Pichia fermentans | 0.25 ± 0.36b | MN650654 |
| JP-17-2 | Pichia fermentans | 0.35 ± 0.22b | MN736502 |
| JT-12-2 | Saturnispora silvae | 0.37 ± 0.30b | MN650662 |
| JP-17-1 | Pichia fermentans | 0.37 ± 0.20b | MN736503 |
| JST-7-1 | Debaryomyces hansenii | 0.39 ± 0.44b | MN704642 |
| NP-10-12 | Meyerozyma guilliermondii | 0.45 ± 0.00b | MN736522 |
| JT-20-3 | Pichia fermentans | 0.46 ± 0.20b | MN653215 |
| JP-11-3 | Pichia fermentans | 0.47 ± 0.34b | MN736541 |
| JST-8-2 | Pichia fermentans | 0.49 ± 0.40b | MN704646 |
| JT-1-5 | Pichia fermentans | 0.55 ± 0.21b | MN704563 |
| JST-9-1 | Saturnispora silvae | 0.61 ± 0.08b | MN704647 |
| NP-8-5 | Pichia fermentans | 0.64 ± 0.00b | MN736543 |
| JT-6-2 | Pichia fermentans | 0.67 ± 0.06b | MN704574 |
| JP-1-11 | Pichia fermentans | 0.76 ± 0.12b | MN736544 |
| JT-14-1 | Pichia fermentans | 0.81 ± 0.19b | MN704650 |
| JT-20-4 | Pichia fermentans | 0.84 ± 0.00b | MN736545 |
| NP-7-5 | Saccharomyces cerevisiae | 4.98 ± 0.00a | MN736547 |
| Kiwifruit Juice | Kiwifruit Wine | |
|---|---|---|
| pH | 3.41 ± 0.02a | 3.45 ± 0.05a |
| SSC (%) | 13.0 ± 0.0 | 6.0 ± 0.0 |
| Total sugars (g/100 g) | 11.56 ± 0.58a | 4.33 ± 0.34b |
| Acids (g/L) | ||
| Tartaric acid | 0.61 ± 0.03a | 0.41 ± 0.04b |
| Malic acid | 3.09 ± 0.18a | 2.02 ± 0.12b |
| Citric acid | 12.30 ± 0.09a | 11.00 ± 0.10b |
| Total acid | 16.80 ± 0.04a | 15.83 ± 0.60b |
| Active ingredients | ||
| VC (mg/100 mL) | 4.85 ± 0.96a | 5.53 ± 1.41a |
| Total flavonoids (mg/mL) | 0.078 ± 0.002a | 0.088 ± 0.001a |
| Total phenol (mg/L) | 109.89 ± 0.29a | 60.46 ± 3.49b |
| Amino Acids Content (g/100 g) | Kiwifruit Juice | Kiwifruit Wine |
|---|---|---|
| Aspartic acid | 0.07 ± 0.00a | 0.06 ± 0.01a |
| Threonine | 0.03 ± 0.00 | 0.03 ± 0.00 |
| Serine | 0.03 ± 0.00 | 0.02 ± 0.00 |
| Glutamic acid | 0.21 ± 0.00a | 0.16 ± 0.00a |
| Glycine | 0.03 ± 0.00 | 0.03 ± 0.00 |
| Alanine | 0.03 ± 0.00 | 0.03 ± 0.00 |
| Valine | 0.03 ± 0.00 | 0.03 ± 0.00 |
| Methionine | n.d. | n.d. |
| Isoleucine | 0.03 ± 0.00 | 0.03 ± 0.00 |
| Leucine | 0.03 ± 0.00 | 0.02 ± 0.00 |
| Tyrosine | 0.02 ± 0.00 | 0.02 ± 0.00 |
| Phenylalanine | 0.02 ± 0.00 | 0.02 ± 0.00 |
| Lysine | 0.03 ± 0.00 | 0.03 ± 0.00 |
| Histidine | 0.02 ± 0.01a | 0.01 ± 0.00a |
| Arginine | 0.07 ± 0.00 | 0.05 ± 0.00 |
| Proline | 0.02 ± 0.00 | 0.03 ± 0.00 |
| EAA | 0.17 ± 0.00 | 0.16 ± 0.00 |
| TAA | 0.66 ± 0.01a | 0.57 ± 0.01b |
| E/T (%) | 25.76 ± 0.39b | 28.32 ± 0.25a |
| Compound Name | Kiwifruit Juice | Kiwifruit Wine |
|---|---|---|
| Alcohols | ||
| (Z)-2-Hexen-1-ol | 2.141 ± 0.107 | n.d. |
| 3-methyl-1-Butanol | n.d. | 26.849 ± 0.856 |
| 2-methyl-1-Butanol | n.d. | 14.355 ± 0.792 |
| 2,3-Butanediol | n.d. | 1.134 ± 0.000 |
| 1-Hexanol | n.d. | 2.677 ± 0.513 |
| Phenylethyl Alcohol | 1.672 ± 0.427b | 154.082 ± 11.247a |
| (Z)-3,7-dimethyl-3,6-Octadien-1-ol | n.d. | 3.020 ± 0.000 |
| Subtotal | 3.813 | 202.117 |
| Aldehydes | ||
| Furfural | 1.645 ± 1.236 | n.d. |
| €-2-Heptenal | 0.456 ± 0.000 | n.d. |
| Benzaldehyde | 8.672 ± 0.083a | 3.183 ± 0.335b |
| (E,E)-2,4-Heptadienal | 1.272 ± 0.025 | n.d. |
| Benzeneacetaldehyde | 0.761 ± 0.085a | 1.912 ± 0.041a |
| €-2-Octenal | 1.394 ± 0.769 | n.d. |
| Nonanal | 1.728 ± 0.009 | n.d. |
| 4-ethyl-Benzaldehyde | n.d. | 1.464 ± 0.000 |
| Decanal | 0.429 ± 0.007a | 0.833 ± 0.103a |
| 2,4-dimethyl-Benzaldehyde | 5.369 ± 0.222 | n.d. |
| €-2-Decenal | 2.870 ± 0.071a | 2.983 ± 0.320a |
| 2-Undecenal | n.d. | 3.104 ± 0.000 |
| Subtotal | 24.596 | 13.479 |
| Ketones | ||
| 3,5-Octadien-2-one | 0.745 ± 0.000 | n.d. |
| (E)-1-(2,6,6-trimethyl-1,3-cyclohexadien-1-yl)-2-Buten-1-one | 0.323 ± 0.014 | n.d. |
| (E)-6,10-dimethyl-5,9-Undecadien-2-one | 0.405 ± 0.024 | n.d. |
| 5-hexyldihydro-2(3H)-Furanone | n.d. | 1.437 ± 0.027 |
| Subtotal | 1.473 | 1.437 |
| Esters | ||
| Propanoic acid, 2-methyl-, 3-hydroxy-2,2,4-trimethylpentyl ester | 0.357 ± 0.013 | n.d. |
| 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | 0.394 ± 0.013 | n.d. |
| Decanoic acid, ethyl ester | n.d. | 0.897 ± 0.058 |
| Dodecanoic acid, ethyl ester | n.d. | 2.105 ± 0.334 |
| Tetradecanoic acid, ethyl ester | n.d. | 1.109 ± 0.155 |
| Ethyl 9-hexadecenoate | n.d. | 20.050 ± 2.586 |
| Hexadecanoic acid, ethyl ester | n.d. | 17.555 ± 1.772 |
| Linoleic acid ethyl ester | n.d. | 9.533 ± 0.717 |
| (E)-9-Octadecenoic acid ethyl ester | n.d. | 20.949 ± 0.997 |
| Subtotal | 0.751 | 72.198 |
| Acids | ||
| n-Hexadecanoic acid | 0.506 ± 0.128a | 0.894 ± 0.122a |
| Oleic Acid | 1.794 ± 1.328a | 0.859 ± 0.08a |
| (E)-9-Octadecenoic acid | n.d. | 1.078 ± 0.114 |
| Octadecanoic acid | 0.616 ± 0.099 | n.d. |
| Subtotal | 2.916 | 2.831 |
| Phenols | ||
| 4-ethyl-Phenol | n.d. | 7.852 ± 0.466 |
| 4-ethyl-2-methoxy-Phenol | 1.078 ± 0.000b | 39.663 ± 3.220a |
| Subtotal | 1.078 | 47.515 |
| Alkene | ||
| D-Limonene | n.d. | 1.224 ± 1.040 |
| Subtotal | 0.000 | 1.224 |
| Total | 34.627 | 340.801 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, W.; Chen, T.; Yang, H.; Li, E. Isolation and Selection of Non-Saccharomyces Yeasts Being Capable of Degrading Citric acid and Evaluation Its Effect on Kiwifruit Wine Fermentation. Fermentation 2020, 6, 25. https://doi.org/10.3390/fermentation6010025
Zhong W, Chen T, Yang H, Li E. Isolation and Selection of Non-Saccharomyces Yeasts Being Capable of Degrading Citric acid and Evaluation Its Effect on Kiwifruit Wine Fermentation. Fermentation. 2020; 6(1):25. https://doi.org/10.3390/fermentation6010025
Chicago/Turabian StyleZhong, Wu, Tuo Chen, Hong Yang, and Erhu Li. 2020. "Isolation and Selection of Non-Saccharomyces Yeasts Being Capable of Degrading Citric acid and Evaluation Its Effect on Kiwifruit Wine Fermentation" Fermentation 6, no. 1: 25. https://doi.org/10.3390/fermentation6010025
APA StyleZhong, W., Chen, T., Yang, H., & Li, E. (2020). Isolation and Selection of Non-Saccharomyces Yeasts Being Capable of Degrading Citric acid and Evaluation Its Effect on Kiwifruit Wine Fermentation. Fermentation, 6(1), 25. https://doi.org/10.3390/fermentation6010025