How to Deal with Uninvited Guests in Wine: Copper and Copper-containing Oxidases

Abstract
:1. Introduction
2. Copper
2.1. Occurrence and Effects of Cu in Must and Wine
2.2. Measurements to Reduce Cu Concentrations in Must and Wine
2.2.1. Chemical and Physical Methods
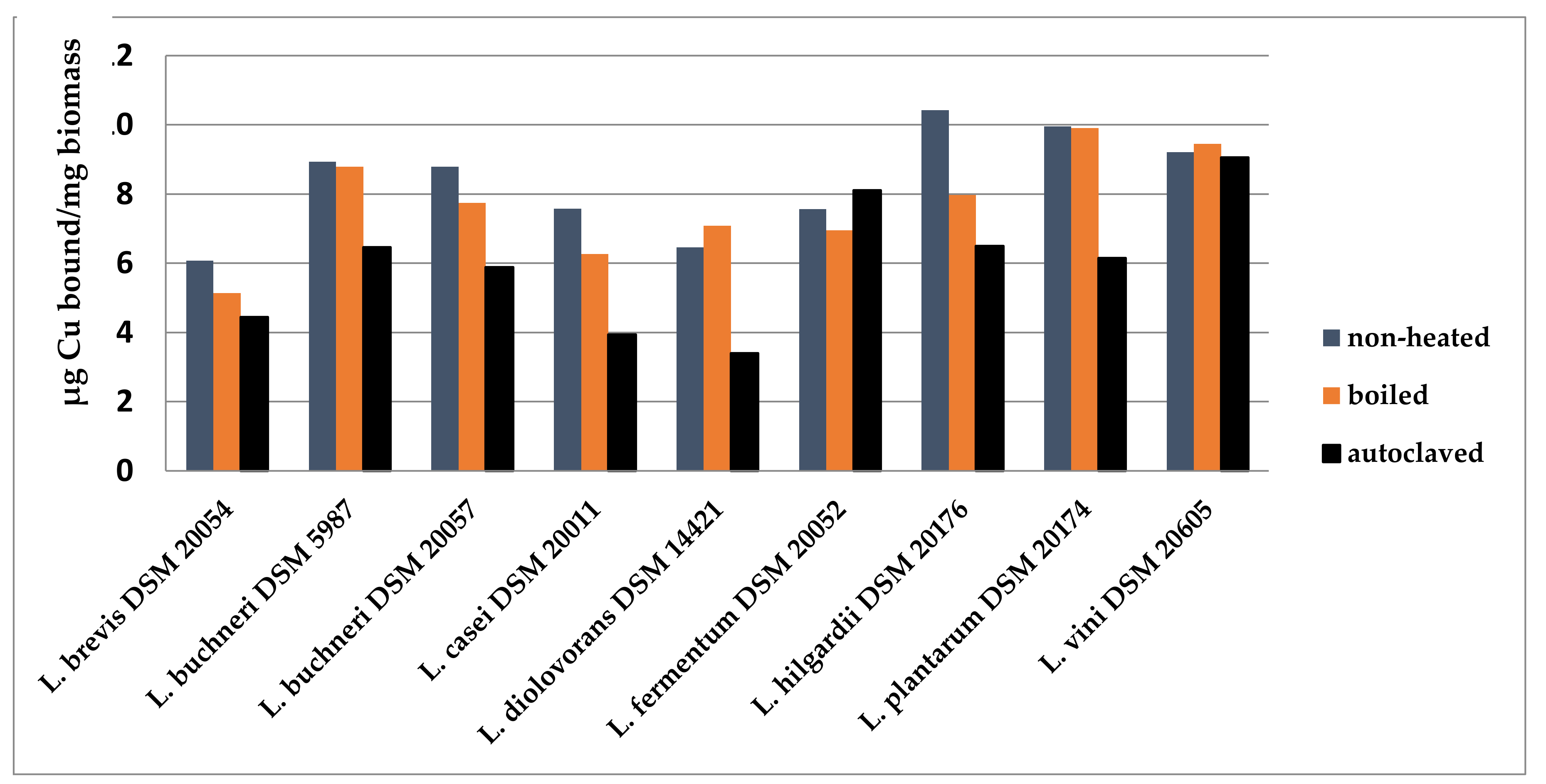
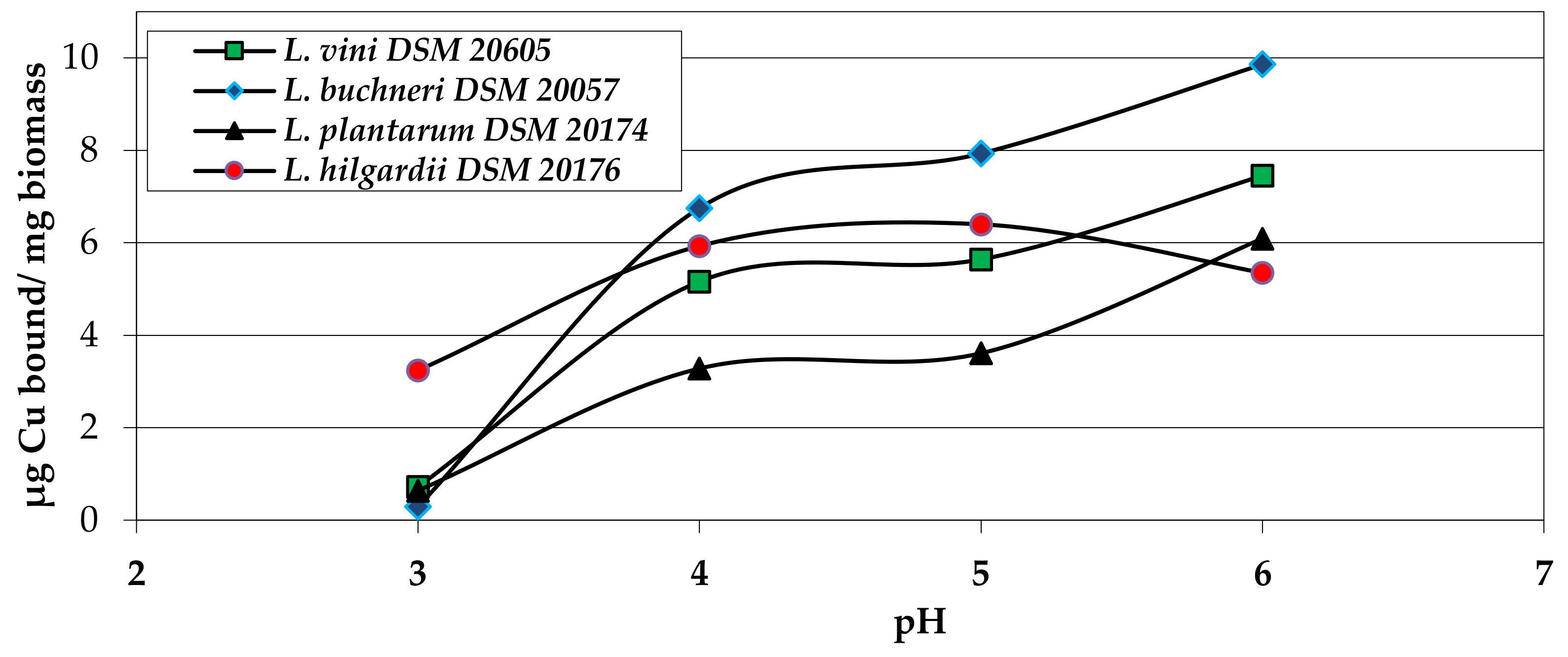
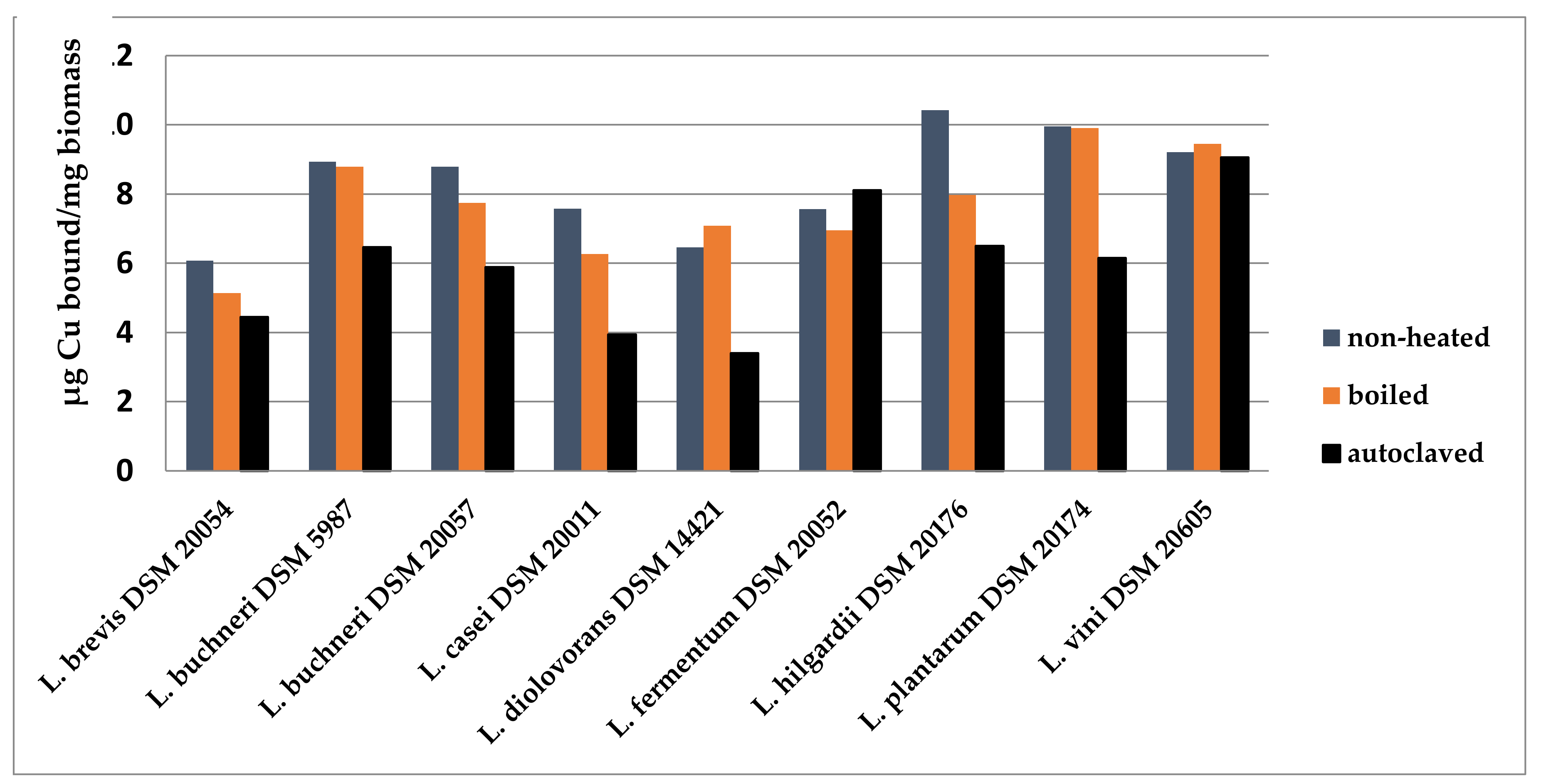
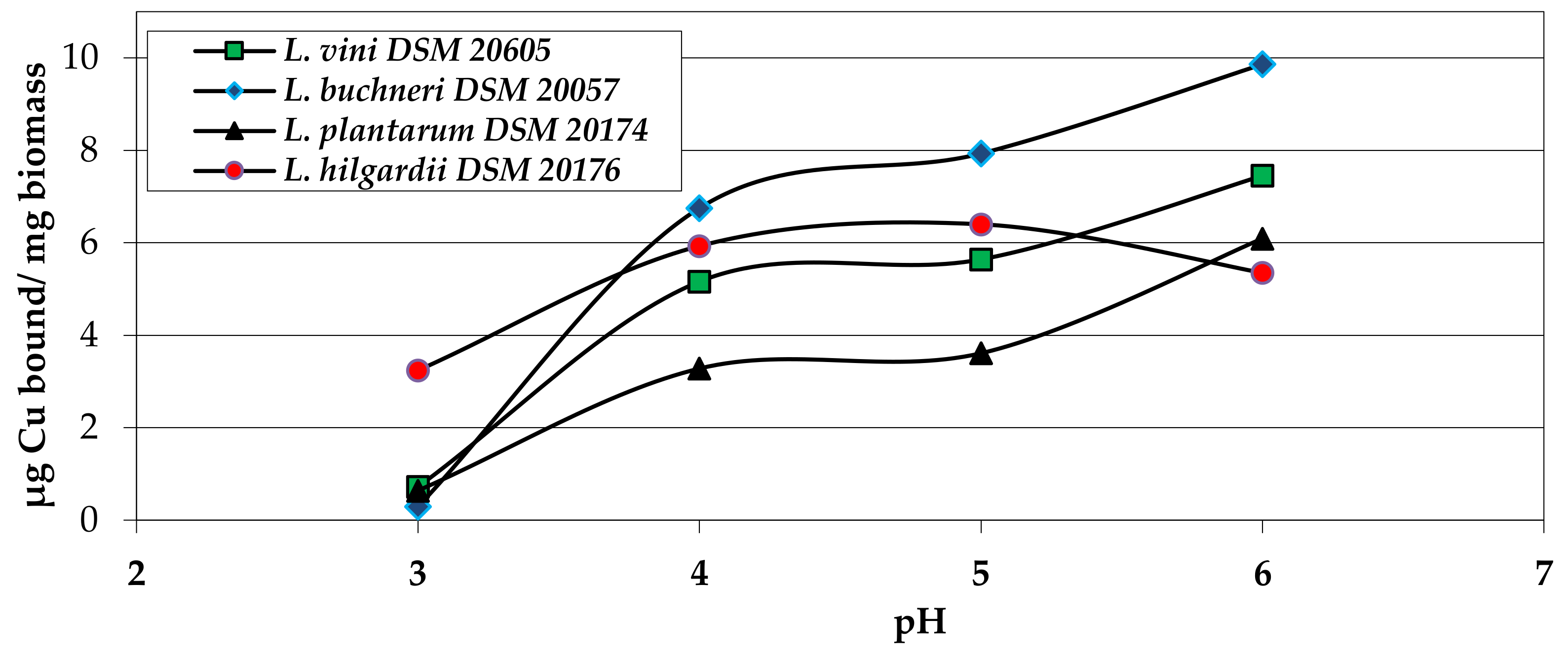
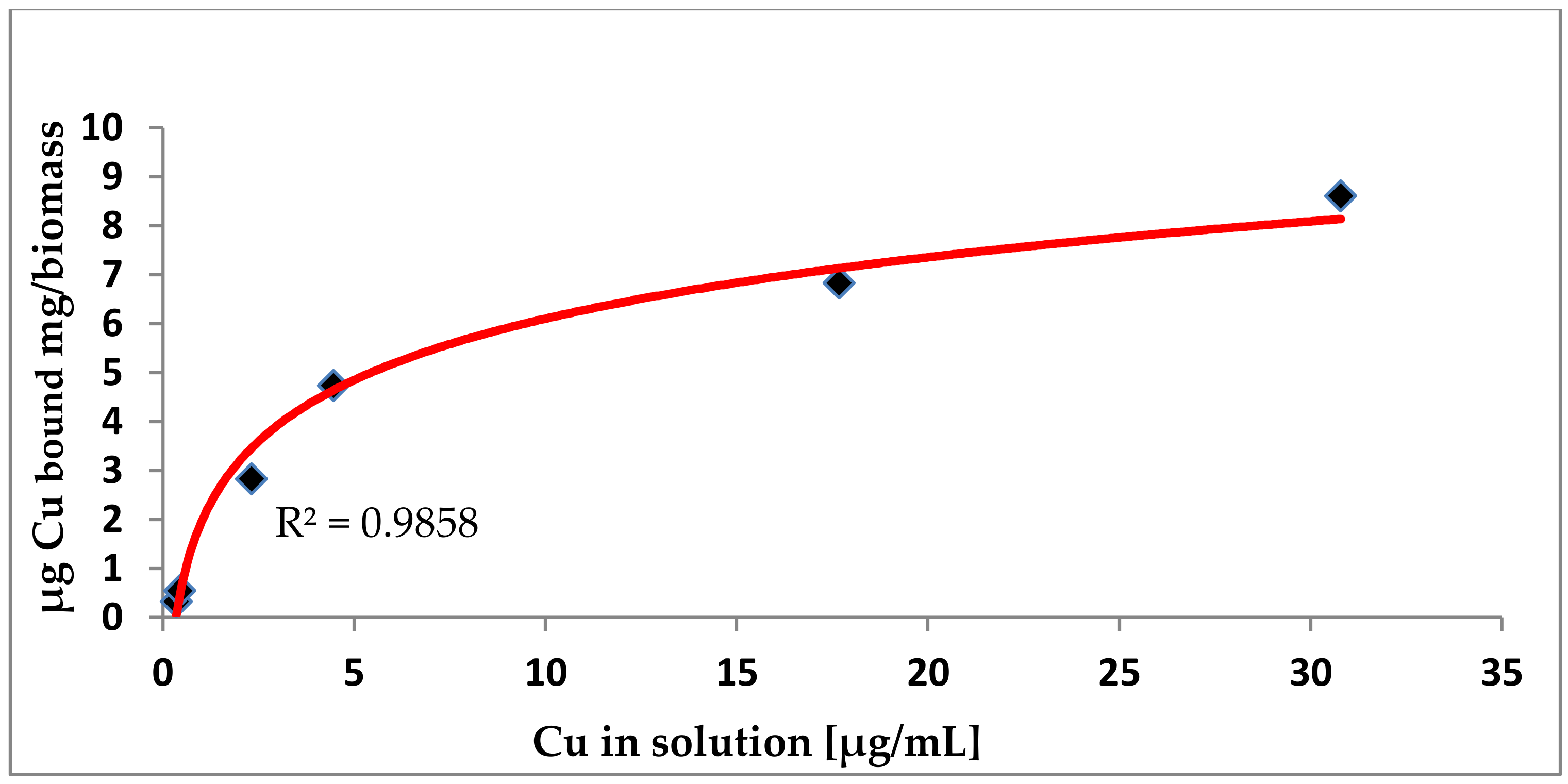
2.2.2. Biosorption
3. Copper-Containing Oxidases
3.1. General Features of Laccases
3.2. The Impact of Botrytis Cinerea Laccases on Wine Quality
3.3. Strategies to Eliminate Laccase Activities in Must and Wine
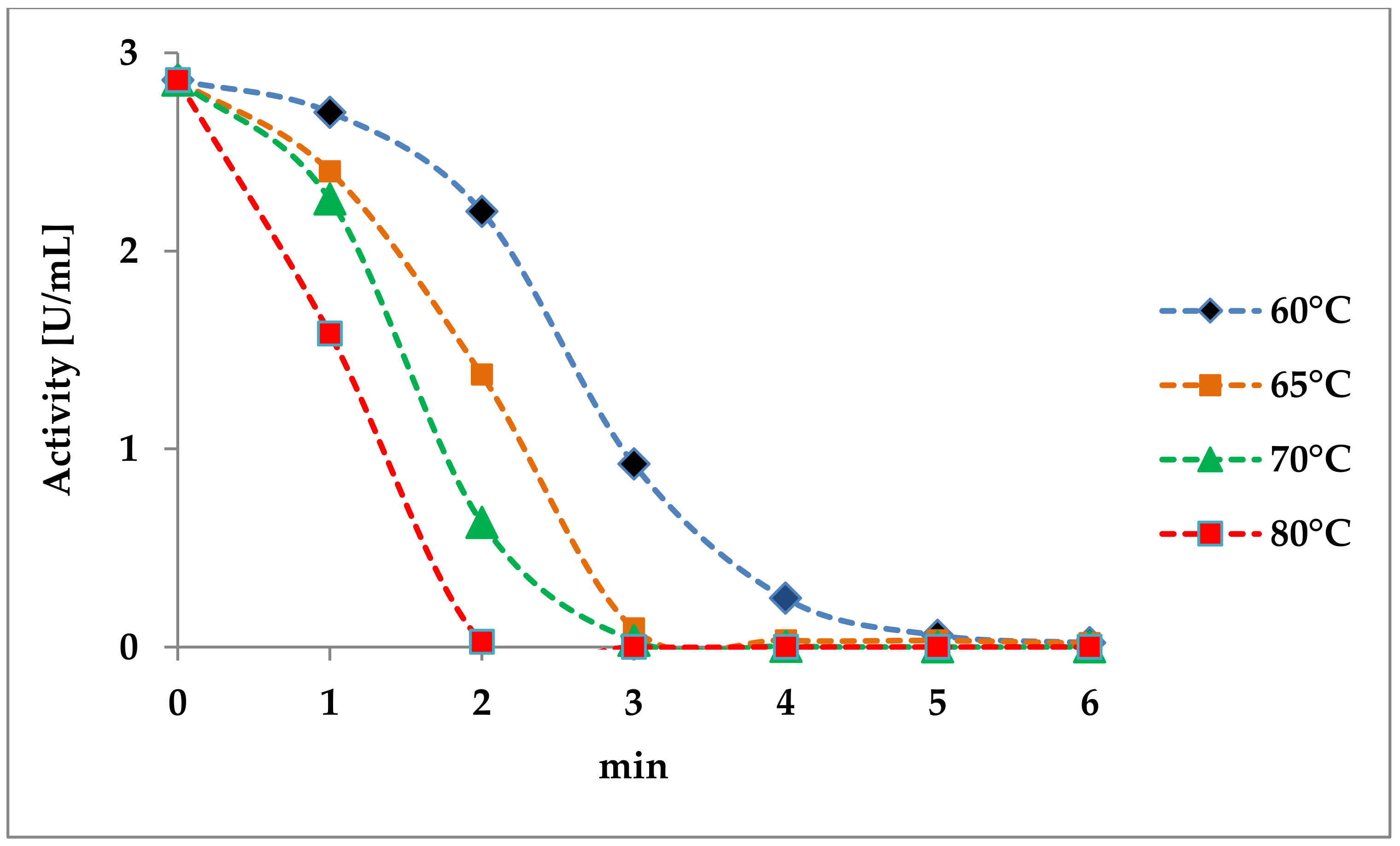
3.3.1. Heat Treatment
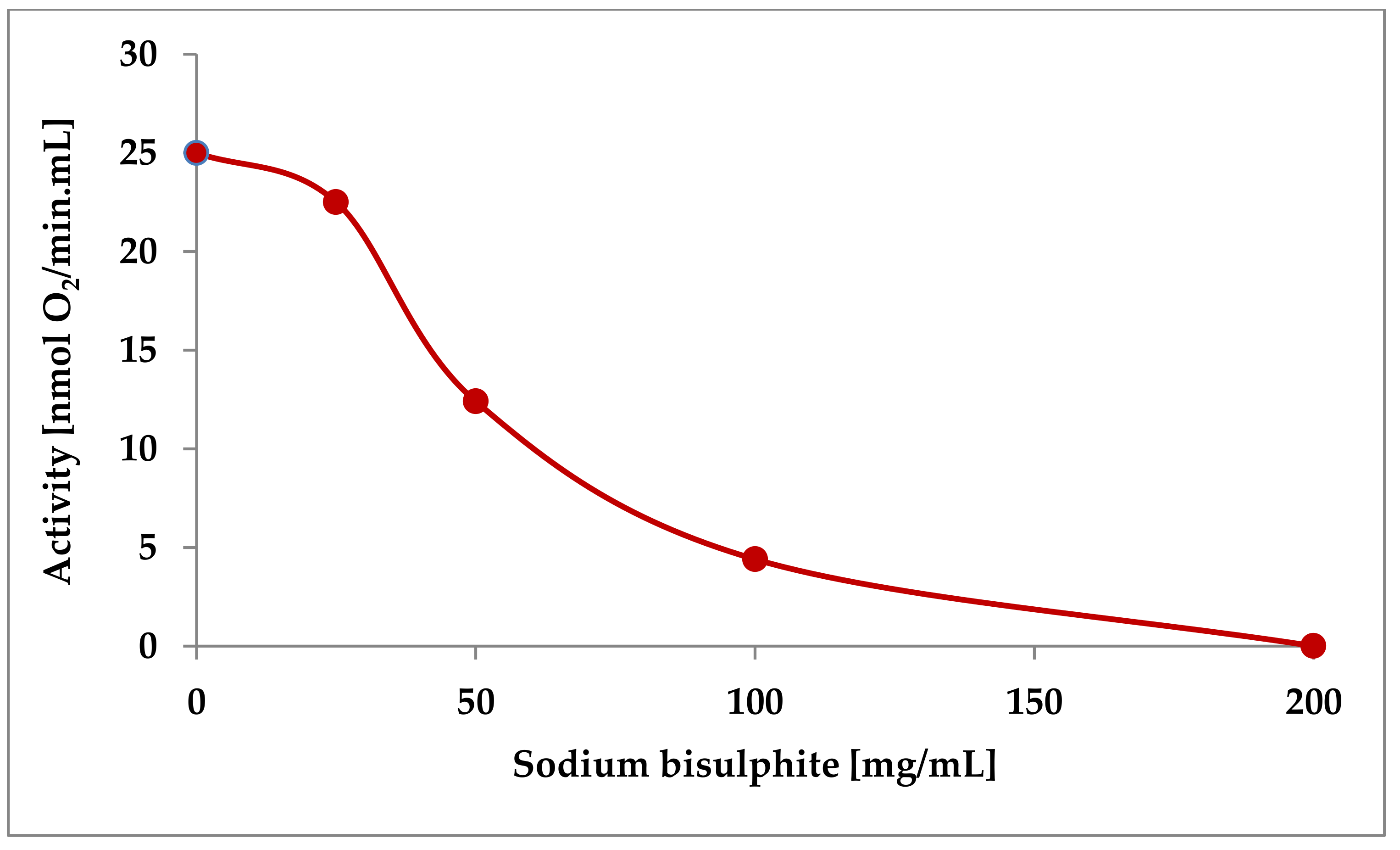
3.3.2. Addition of Sulphite
3.3.3. Bentonite as A Fining Agent for Must and Wine Clarification
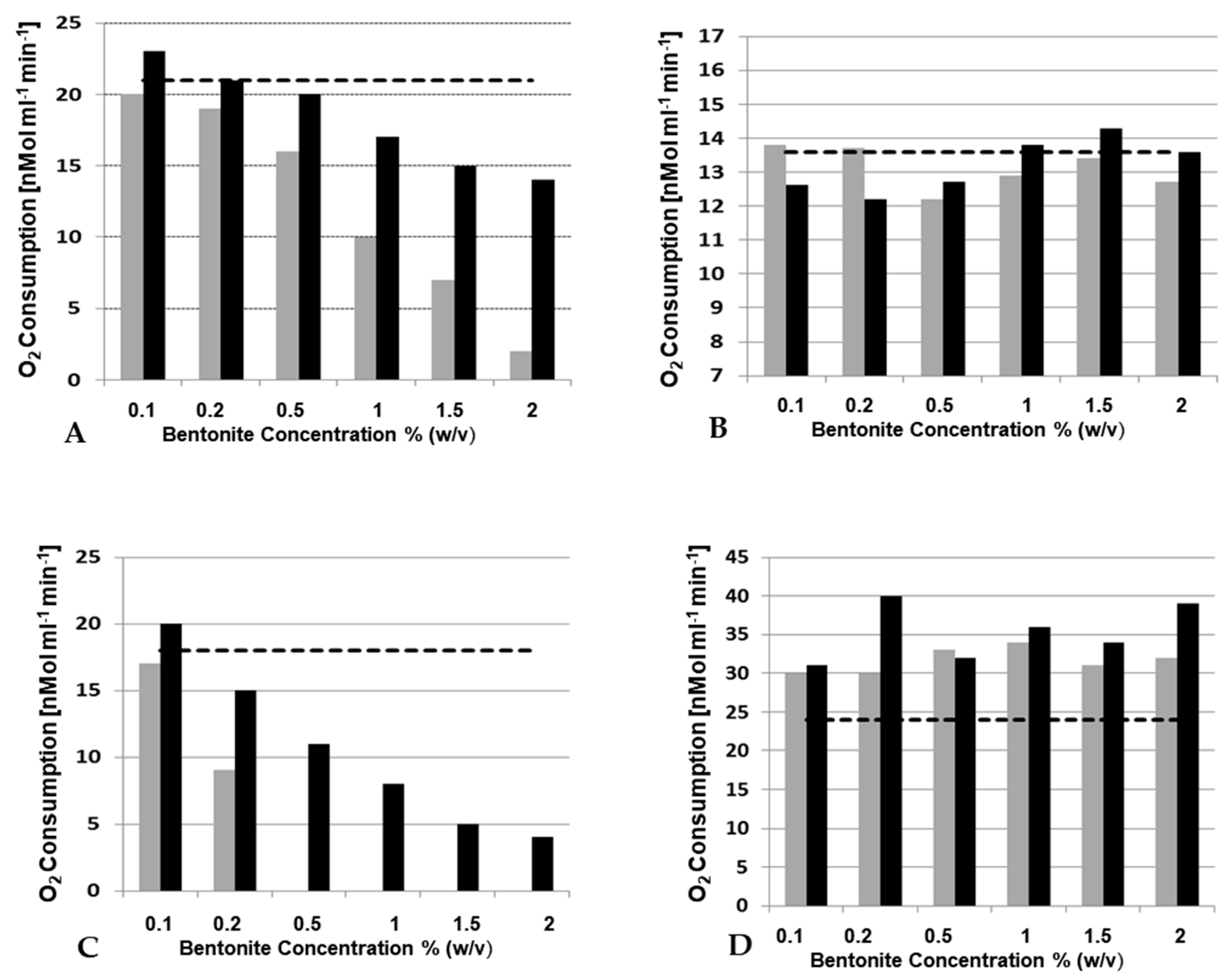
3.3.4. Influence of Bentonite on Laccase Activity?
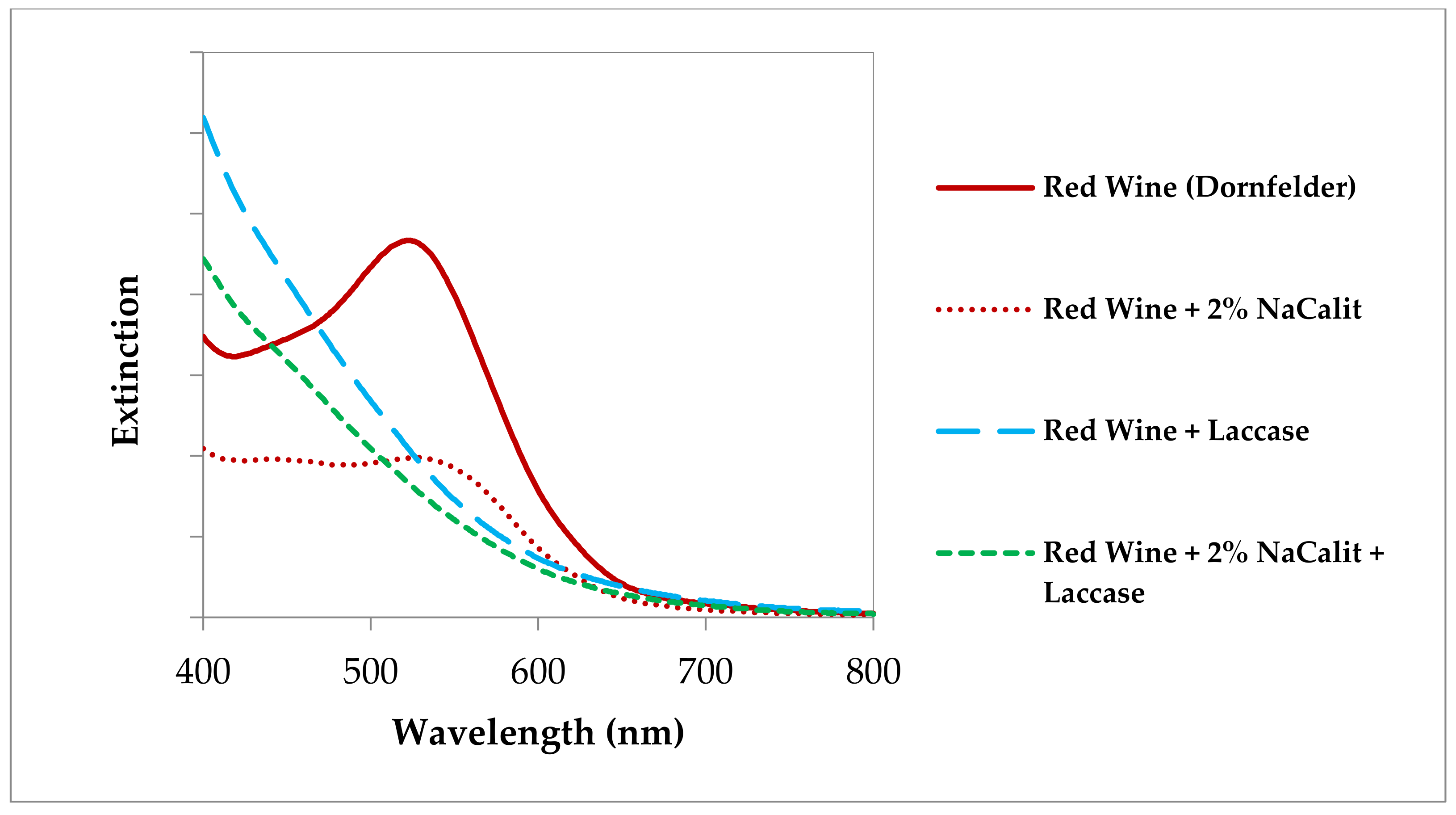
3.3.5. Ooenological Tannins to Eliminate Laccase Activities in Wine
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Singleton, V.L. Oxygen with phenols and related reactions in musts, wines, and model systems: Observations and practical implications. Am. J. Enol. Vitic. 1987, 38, 69–77. [Google Scholar]
- Li, H.; Guo, A.; Wang, H. Mechanisms of oxidative browning of wine. Food Chem. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Sioumis, N.; Kallithraka, S.; Tsoutsouras, E.; Makris, D.P.; Kefalas, P. Browning development in white wines: Dependence on compositional parameters and impact on antioxidant characteristics. Eur. Food Res. Technol. 2005, 220, 326–330. [Google Scholar] [CrossRef]
- Claus, H.; Sabel, A.; König, H. Wine Phenols and Laccase: An ambivalent relationship. In Wine: Phenolic Composition, Classification and Health Benefits; El Rayess, Y., Ed.; Nova Publishers: Hauppauge, NY, USA, 2014; pp. 155–185. [Google Scholar]
- Claus, H. Laccases of Botrytis cinerea. In Biology of Microorganisms on Grapes, in Must and Wine, 2nd ed.; König, H., Unden, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 339–356. [Google Scholar]
- Fronk, P.; Riebel, M.; Decker, H. Polyphenol oxidase from wine grapes. In Biology of Microorganisms on Grapes, in Must and Wine, 2nd ed.; König, H., Unden, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 357–374. [Google Scholar]
- Riebel, M.; Sabel, A.; Claus, H.; Xia, N.; Li, H.; König, H.; Decker, H.; Fronk, P. Antioxidant capacity of phenolic compounds on human cell lines as affected by grape-tyrosinase and Botrytis-laccase oxidation. Food Chem. 2017, 229, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Claus, H. Copper-containing oxidases: Occurrence in soil microorganisms, properties and applications. In Soil Heavy Metals, Soil Biology Vol. 19; Sherameti, I., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 281–313. [Google Scholar]
- Araya, M.; Olivares, M.; Pizzaro, F. Copper in human health. Int. J. Environ. Health 2007, 1, 608–620. [Google Scholar] [CrossRef]
- Wagner, E.; Augustin, T.; Brunn, H. Metallgehalt in Wein—Bestimmung in deutschen und ausländischen Weinen. Dtsch. Lebensm. Rundsch. 2010, 7, 385–388. [Google Scholar]
- Nogueirol, R.C.; Alleoni, L.R.F.; Nachtigall, G.R.; de Melo, G.W. Sequential extraction and availability of copper in Cu fungicide-amended vineyard soil from Southern Brazil. J. Hazard. Mater. 2010, 181, 931–937. [Google Scholar] [CrossRef] [Green Version]
- Zoecklein, B.W.; Fugelsang, K.C.; Gump, B.H.; Nury, F.S. Wine Analysis and Production; Springer: Boston, MA, USA, 1995. [Google Scholar]
- Clark, A.C.; Wilkes, E.N.; Scollary, G.R. Chemistry of copper in white wine: A review. Aust. J. Grape Wine Res. 2015, 21, 339–350. [Google Scholar] [CrossRef]
- Volpe, M.G.; la Cara, F.; Volpe, F.; de Mattia, A.; Serino, V.; Petitto, F.; Zavalloni, C.; Limone, F.; Pellecchia, R.; de Prisco, P.P.; et al. Heavy metal uptake in the enological food chain. Food Chem. 2009, 117, 553–560. [Google Scholar] [CrossRef]
- Schut, S.; Claus, H.; König, H. Der Einsatz von Laktobazillen zur Entfernung von Kupfer aus Most und Wein. In Deutsches Weinbau Jahrbuch; Schultz, H.R., Stoll, M., Eds.; Ulmer Verlag Stuttgart: Stuttgart, Germany, 2012; pp. 137–145. [Google Scholar]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, S. Handbook of Enology, Vol 2, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006; ISBN 978-0-470-01037-2. [Google Scholar]
- Schubert, M.; Glomb, M.A. Analysis and chemistry of migrants from wine fining polymers. J. Agric. Food Chem. 2010, 58, 8300–8304. [Google Scholar] [CrossRef]
- Mira, H.; Leite, P.; Catarino, S.; Ricardo-da-Silva, J.M.; Curvelo-Garcia, A.S. Metal reduction in wine using PVI-PVP colpolymer and its effects on chemical and sensory characters. VITIS 2007, 46, 138–147. [Google Scholar]
- Ficagna, E.; Gava, A.; Rossato, S.B.; Rombaldi, C.V.; Borsato, D. Effect on Merlot red wine of fining agents mixture: Application of the simplex centroid design. Food Sci. Technol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.P.; Huang, C.P.; Morehart, A.L. The removal of Cu(II) from dilute aqueous solution by Saccharomyces cerevisiae. Water Res. 1990, 24, 433–439. [Google Scholar] [CrossRef]
- Junghans, K.; Straube, G. Biosorption of copper by yeasts. Biol. Met. 1991, 4, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, F.; Halttunen, T.; Tahvonen, R.; Salminen, S. Probiotic bacteria as potential detoxification tools: Assessing their heavy metal binding isotherms. Can. J. Microbiol. 2006, 52, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.H.A.; Kim, S.J.; Jung, A.Y.; Joo, J.H.; Oh, S.E.; Yang, J.E. Biosorptive capacity of Cd(II) and Cu(II) by lyophilized cells of Pseudomonas stutzeri. J. Gen. Appl. Microbiol. 2009, 55, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Schut, S.; Zauner, S.; Hampel, G.; König, H.; Claus, H. Biosorption of copper by wine-relevant lactobacilli. Int. J. Food Microbiol. 2011, 145, 126–131. [Google Scholar] [CrossRef]
- Naja, G.M.; Murphy, V.; Volesky, B. Biosorption, Metals. In Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology; Flickinger, M.C., Ed.; Wiley Online Library: Hoboken, NJ, USA, 2010. [Google Scholar]
- Hansda, A.; Kumar, V. Biosorption of copper by bacterial adsorbents: A review. Res. J. Environ. Tox. 2015, 9, 45–58. [Google Scholar]
- König, H.; Fröhlich, J. Lactic acid bacteria. In Biology of Microorganisms on Grapes, in Must and Wine, 2nd ed.; König, H., Unden, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 3–41. [Google Scholar]
- Naughton, D.P.; Petróczi, A. Heavy metal ions in wines: Meta-analysis of target hazard quotients reveal health risks. Chem. Cent. J. 2008, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Pardo, R.; Herguedas, M.; Barrado, E. Biosorption of cadmium, copper, lead and zinc by inactive biomass of Pseudomonas putida. Anal. Bioanal. Chem. 2003, 376, 26–32. [Google Scholar] [CrossRef]
- Mrvčić, J.; Prebeg, T.; Barišić, L.; Stanzer, D.; Bačun-Družina, V.; Stehlik-Tomas, V. Zinc binding by lactic acid bacteria. Food Technol. Biotechnol. 2009, 47, 381–388. [Google Scholar]
- Mrvčić, J.; Stanzer, D.; Bačun-Družina, V.; Stehlik-Tomas, V. Copper binding by lactic acid bacteria (LAB). Biosci. Microflora 2009, 28, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Cabuka, A.; Akar, T.; Tunali, S.; Gedikli, S. Biosorption of Pb(II) by industrial strain of Saccharomyces cerevisiae immobilized on the biomatrix of cone biomass of Pinus nigra: Equilibrium and mechanism analysis. Chem. Eng. J. 2007, 131, 293–300. [Google Scholar] [CrossRef]
- Strong, P.J.; Claus, H. Laccase: A review of its past and its future in bioremediation. Crit. Rev. Environ. Sci. Technol. 2011, 41, 373–434. [Google Scholar] [CrossRef]
- El Rayess, Y. Wine: Phenolic Composition, Classification and Health Benefits; Nova Publishers: New York, NY, USA, 2014; ISBN 978-1-63321-048-6. [Google Scholar]
- De Leonardis, A.; Lustrato, G.; Macciola, V.; Ranalli, G. APpLication of chemical and physical agents in model systems to controlling phenoloxidase enzymes. Eur. Food Res. Technol. 2010, 23, 603–610. [Google Scholar] [CrossRef]
- Zivkovic, K.; König, H.; Claus, H. Wirkung von Bentonit auf die Laccase-Aktivität in Most und Wein. Dtsch. Lebensm. Rundsch. 2011, 107, 575–582. [Google Scholar]
- Campos, F.M.; Couto, J.A.; Hogg, T. Utilisation of natural and by-products to improve wine safety. In Wine Safety, Consumer Preference, and Human Health; Moreno-Arribas, M.V., Bartolomé Sualdea, B., Eds.; Springer: Cham, Switzerland, 2016; pp. 27–49. [Google Scholar]
- Marchal, R.; Jeandet, P. Use of Enological additives for colloid and tartrate salt stabilisation in white wines and for improvement of sparkling wine foaming properties. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, M.C., Eds.; Springer: New York, NY, USA, 2010; pp. 127–158. [Google Scholar]
- Waters, E.J.; Colby, C.B. Proteins. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, C., Eds.; Springer: New York, NY, USA, 2009; pp. 213–226. [Google Scholar]
- Jaeckels, N.; Tenzer, S.; Rosch, A.; Scholten, G.; Decker, H.; Fronk, P. β-glucosidase removal due to bentonite fining during wine making. Eur. Food Res. Technol. 2015, 241, 253–262. [Google Scholar] [CrossRef]
- Blade, H.; Boulton, R. Adsorption of protein by bentonite in a model wine solution. Am. J. Enol. Vitic. 1988, 39, 193–199. [Google Scholar]
- Claus, H.; Filip, Z. Behaviour of phenoloxidases in the presence of clays and other soil-related adsorbents. Appl. Microbiol. Biotechnol. 1988, 28, 506–511. [Google Scholar] [CrossRef]
- Stanković, S.; Jović, S.; Živković, J. Bentonite and gelatine impact on the red wine coloured matter. Food Technol. Biotechnol. 2004, 42, 183–188. [Google Scholar]
- Versari, A.; du Toit, W.; Parpinello, G.P. Oenological tannins: A review. Aust. J. Grape Wine Res. 2013, 19, 1–10. [Google Scholar] [CrossRef]
- Vignault, A.; Pascual, O.; Jourdes, M.; Moine, V.; Fermaud, M.; Roudet, J.; Canals, J.M.; Teissedre, P.L.; Zamora, F. Impact of enological tannins on laccase activity. OENO One 2019, 53. [Google Scholar] [CrossRef]
- Vignault, A.; Gombau, J.; Jourdes, M.; Moine, V.; Canais, J.M.; Fermaud, M.; Roudet, J.; Zamora, F.; Teissedre, P.L. Oenological tannins to prevent Botrytis cinerea damage in grapes and musts: Kinetics and electrophoresis characterization of laccase. Food Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Selenska-Pobell, S.; Merroun, M. Accumulation of heavy metals by microorga nisms: Biomineralization and nanocluster formation. In Prokaryotic Cell Wall Compounds Structure and Biochemistry; König, H., Claus, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 483–500. [Google Scholar]
- Dohm, N.; Petri, A.; Schlander, M.; Schlott, B.; König, H.; Claus, H. Molecular and biochemical properties of the S-layer protein from the wine bacterium Lactobacillus hilgardii B706. Arch. Microbiol. 2011, 193, 251–261. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; de Keersmaecker, S.C.J. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, V.; Rodriguez, H.; Carrascosa, A.V.; de las Rivas, B.; Garcia-Murono, M.R. In vitro removal of ochratoxin A by wine lactic acid bacteria. J. Food Prot. 2007, 70, 2155–2160. [Google Scholar] [CrossRef]
- Brandolini, V.; Tedeschi, P.; Capece, A.; Maietti, A.; Mazzotta, D.; Salzano, G.; Paparella, A.; Romano, P. Saccharomyces cerevisiae wine strains differing in copper resistance exhibit different capability to reduce copper content in wine. World J. Microbiol. Biotechnol. 2002, 18, 499–503. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Limits and Normal Values/Effects | Cu Concentration (mg/L) |
|---|---|
| Normal value in wine | 0.7–0.8 |
| Maximum content allowed in wine | 2.0 |
| Concentration range in must | 0.09–0.99 |
| Copper catalyzed oxidations (browning reaction) | in traces |
| Copper casse | > 0.5 |
| Influencing fermentation of sparkling wine (very rare) | > 20 |
| Sluggish must fermentation | 25–50 |
| Inhibition of Oenococcus oeni Microenos B1 | 5*; 10** |
| Inhibition of Saccharomyces cerevisiae SN9 / SN41 / CCT0472 | 32/320/75 |
| Inhibition of Lactobacillus fermentum CCT1400 / CCT0559 | 75/300 |
| Inhibition of Lactobacillus mesenteroides CCT0582 / CCT0367 | 75/150 |
| Biosorbent | pH Optimum | Biomass Added (mg/mL) | qh (µg Cu2+ /mg Cells) |
|---|---|---|---|
| Bacteria | |||
| Arthrobacter sp. | 3.5–6.0 | 0.4 | 148.0 |
| Bacillus sp. (ATS-1) | 5.0 | 2.0 | 16.3 |
| Bacillus subtilis IAM 1026 | 5.0 | 0.5 | 20.8 |
| Enterobacter sp. J1 | 5.0 | 1.0 | 32.5 |
| Lactobacillus brevis ID9262 | 5.0 | 0.5 | 26.5 |
| Lactobacillus buchneri DSM 20057 (living / dead) | 5.0 | 1.0 | 10.5/9.9 |
| Lactobacillus hilgardii DSM 20176 (living/dead) | 5.0 | 1.0 | 10.0/8.8 |
| Lactobacillus plantarum DSM 20174 (living /dead) | 5.0 | 1.0 | 9.7/8.6 |
| Lactobacillus plantarum ID9263 | 5.0 | 0.5 | 15.5 |
| Lactobacillus vini DSM 20605 (living /dead) | 5.0 | 1.0 | 12.3/9.7 |
| Leuconostoc mesenteroides ID9261 | 5.0 | 0.5 | 26.3 |
| Micrococcus luteus | 5.0 | 0.5 | 33.5 |
| Pseudomonas aeruginosa PU21 | 5.0 | 1.0 | 23.1 |
| Pseudomonas cepacia | 7.0 | n. d | 65.3 |
| Pseudomonas putida | 6.6 | n. a. | 6.6 |
| Pseudomonas putida CZ1 | 4.5 | 1.0 | 15.8 |
| Pseudomonas stutzeri IAM 12097 | 5.0 | 1.0 | 22.9 |
| Pseudomonas stutzeri KCCM 34719 | 5.0 | 1.0 | 36.2 |
| Pseudomonas syringae | n.d. | 0.28 | 25.4 |
| Sphaerotilus natans | 5.5 | n.d. | 5.4 |
| Streptomyces noursei | 5.5 | 3.5 | 9.0 |
| Synechocystis sp. | 4.5 | 1.0 | 23.4 |
| Zoogloea ramigera | 5.5 | 0.83 | 270.0 |
| Zoogloea ramigera | 4.0 | n. d | 29 |
| Yeasts | |||
| Cryptococcus terreus | 5.5 | 1.0 | 71.8 |
| Pichia guilliermondii | 5.5 | 1.0 | 34.0 |
| Saccharomyces cerevisiae1) | 5.5 | 1.0 | 43.3 |
| Saccharomyces cerevisiae2) (living/dead) | 4.0 | 2.0 | 0.8/0.4 |
| Medium | pH | µg Cu2+/mL in Solution | µg Cu2+/mL Removed (%) | |
|---|---|---|---|---|
| Control | + L. buchneri DSM 20057 | |||
| Water | 5.0 | 3.90 | 0.76 | 3.14 (81.0) |
| Must (white) | 3.5 | 5.64 | 5.20 | 0.44 (7.8) |
| Must (red) | 3.6 | 4.17 | 3.96 | 0.21 (5.0) |
| Wine (white) | 3.3 | 4.16 | 3.60 | 0.56 (13.5) |
| Wine (red) | 3.1 | 3.89 | 2.86 | 1.03 (26.5) |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claus, H. How to Deal with Uninvited Guests in Wine: Copper and Copper-containing Oxidases. Fermentation 2020, 6, 38. https://doi.org/10.3390/fermentation6010038
Claus H. How to Deal with Uninvited Guests in Wine: Copper and Copper-containing Oxidases. Fermentation. 2020; 6(1):38. https://doi.org/10.3390/fermentation6010038
Chicago/Turabian StyleClaus, Harald. 2020. "How to Deal with Uninvited Guests in Wine: Copper and Copper-containing Oxidases" Fermentation 6, no. 1: 38. https://doi.org/10.3390/fermentation6010038
APA StyleClaus, H. (2020). How to Deal with Uninvited Guests in Wine: Copper and Copper-containing Oxidases. Fermentation, 6(1), 38. https://doi.org/10.3390/fermentation6010038