1. Introduction
Chitin (β-(1-4)-poly-
N-acetyl-
d-glucosamine), the second most abundant polysaccharide occurring in nature after cellulose, is found in the exoskeletons of crustaceans, such as crayfish, crabs and shrimps, as well as in the cell walls of fungi, such as mushrooms [
1]. Due to its high biocompatibility, predictable biodegradability, anti-microbial activity and non-toxicity to cells, chitin and its deacetylated derivative chitosan have been deployed in a wide range of emerging biomedical applications [
2]. Prominent examples include the successful fabrication of chitin and chitosan into hydrogels and scaffolds for tissue repair and regeneration, the development of chitosan derivatives into vaccine adjuvants for enhanced immune response against pathogens, the use of chitin-based nanoparticles for drug encapsulation and delivery as well as the coating of chitosan on semiconductor nanocrystals or quantum dots for bioimaging in cancer diagnosis [
3].
Crustacean chitin exists in three different crystalline allomorphs—α, β and γ, classified by the orientation of the microfibrils [
4]. While α-chitin is made up of two
N-
N’-diacetyl-glucosamine units forming two polymer chains in an anti-parallel and stable arrangement, β-chitin enjoys a more flexible structure composed of only one poly-
N-acetylglucosamine unit arranged in parallel polymer chains with no inter-sheet H bonds [
5]. γ-chitin, however, is composed of mixed anti-parallel and parallel arrangements [
6]. Varying degrees of deacetylation have been reported in both crustaceans and fungi, with a continuum of structure between fully acetylated chitin and fully deacetylated chitosan [
7]. Chitin is highly insoluble, and the
N-deacetylation of chitin to form chitosan enables its solubility in diluted acetic and formic acids, increasing its versatility [
8]. The degree of deacetylation indicates the degree of transformation of chitosan from chitin, and chitin is classified as chitosan when the degree of deacetylation rises above 50% [
9].
Industrial extraction of chitin from seafood waste requires the demineralization of minerals such as calcium carbonate and calcium phosphate as well as the deproteinization of proteins contained in crustacean shells [
10]. Demineralization is usually achieved by hydrochloric acid (HCl) treatment to dissolve the calcite minerals while deproteinization is generally obtained after sodium hydroxide (NaOH) processing to remove the proteins [
11]. Both processes require high concentrations of acids and bases as well as elevated temperature conditions, leading to high energy consumption and increased production costs, as well as posing a hazardous threat to the environment [
12]. Hence scientific interest is piqued by the use of biotechnological fermentation to perform economical and environmentally friendly microbial extraction of chitin and chitosan on low-cost industrial by-products and wastes [
13]. Co-fermentation of lactic acid-producing bacteria, such as
Lactobacillus plantarum, protease-producing bacteria, such as
Bacillus subtilis and
Pseudomonas aeruginosa, as well as fungi, such as
Aspergillus niger and
Rhizopus oligosporus, have been reported to facilitate the demineralization and deproteinization of shrimp shells through the secretion of microbial proteases and organic acids [
14].
The fungal species
Mucor circinelloides of the Zygomycetes class has ignited much scientific interest due to the high chitin and chitosan content in its mycelia, which is around 35% of cell wall dry weight, comparable with the 20–30% chitin content found in crustacean shells [
15]. Unlike crustacean chitin, which is a hard composite of highly mineralized chitin protein, the fungal cell wall is a polysaccharide-based three-dimensional network whereby chitin is linked with branched β-1,3- and β-1,6-glucan via a β-1,4 linkage, and hence does not require harsh demineralization [
16]. While the main commercial sources of chitin remain crab and shrimp shells with 80,000 tons of chitin extracted from marine by-products annually, the use of seafood waste is subject to seasonal variations and discontinuous supply [
17]. The ease of cultivation of fungi in the laboratory on cheap substrates can supplement the constant quality and supply of chitosan in the commercial market [
18].
Mucor circinelloides has been reported to be a dimorphic fungus that can be cultured on a wide range of lignocellulosic sugars, including pentoses and hexoses, to produce valuable biochemical products, such as bioethanol, polyunsaturated fatty acids, such as gamma-linolenic acid (omega 6) and essential lipids, such as oleic acids, for biodiesel synthesis [
19]. However, few studies have been performed on the efficacy of
Mucor circinelloides in releasing proteolytic enzymes for the extraction of chitin from shrimp shell waste [
20]. In this paper, we explore a variety of waste substrates for the simultaneous production of chitin from prawn shells and cultivation of fungal chitosan from
Mucor circinelloides by placing prawn shell waste in direct contact with the fermentation of filamentous fungi. The protease secreted from the
Mucor circinelloides is expected to hydrolyze the proteins in the prawn shells, with the released amino acids serving as a nitrogen source for fungal growth and lowering the pH of the fermentation medium, thereby resulting in further demineralization of the prawn shells. The objective of this work is to examine the yield and quality of the extracted crustacean chitin and fungal chitosan in a concurrent fermentation by profile characterization using Fourier Transform Infrared Spectroscopy (FTIR) and Elemental Analysis (EA).
2. Materials and Methods
2.1. Fermentation of Prawn Shell Waste using Mucor circinelloides Van Tieghem ATCC 24905
Mucor circinelloides van Tieghem ATCC 24905 was purchased from the American Type Culture Collection (ATCC). The freeze-dried culture was rehydrated with sterile deionized (DI) water and stock cultures were generated by cultivation of the rehydrated fungal cells on a potato dextrose agar (PDA) plate incubated at 30 °C for 1 day. A total of 5 g of prawn shells was mixed with 10 g of waste as a carbon source. Five types of waste substrates were tested: okara, coffee residues, discarded durian and avocado seeds as well as red grape pomace. The waste residues were collected from food processing industrial factories in Singapore, milled into a fine powder using a blender and dried in a vacuum oven at 60 °C for 1 day before being used for fermentation. The experimental setups were autoclaved at 121 °C for sterilization before 100 mL of sterile DI water was added into each fermentation flask.
A total of 10 mL of sterile DI water was added to the PDA plate and the entire mycelial growth on the agar surface was scraped using a sterile cell spreader to obtain a homogeneous spore suspension. Then 1 mL of the fungal spore suspension was inoculated into each fermentation flask and the mixture was gently shaken to suspend the fungal spores in water. The flasks were then placed in the incubator to ferment at 30 °C, 200 rpm, for 7 days. After 7 days, the deproteinated and demineralized prawn shells as well as the cultured Mucor circinelloides fungal biomass were removed from the fermentation flasks and separated manually with a laboratory metal spoon, washed with DI water and sprayed with ethanol for sterilization before being placed in a 60 °C vacuum oven to dry for 24 h. The dried crustacean chitin was immediately analyzed whereas the dried fungal biomass underwent further chitosan extraction from its cell walls.
Lactobacillus plantarum subsp. plantarum ATCC 14917 and Bacillus subtilis subsp. subtilis ATCC 6051 were inoculated in 5 mL of De Man, Rogosa and Sharpe (MRS) broth and Luria broth (LB), respectively, and incubated overnight at 200 rev · min−1 at 30 °C. The dried fungal biomass was then dropped into a 50 mL Lactobacillus plantarum and Bacillus subtilis growing culture (i.e., 25 mL of MRS broth and 25 mL of LB broth) and left to ferment at 30 °C, 200 rpm, for 5 days. After 5 days, the demineralized and deproteinized fungal mycelia were removed, washed with DI water and sprayed with ethanol for sterilization. They were then dried in the 60 °C vacuum oven for 24 h before analysis.
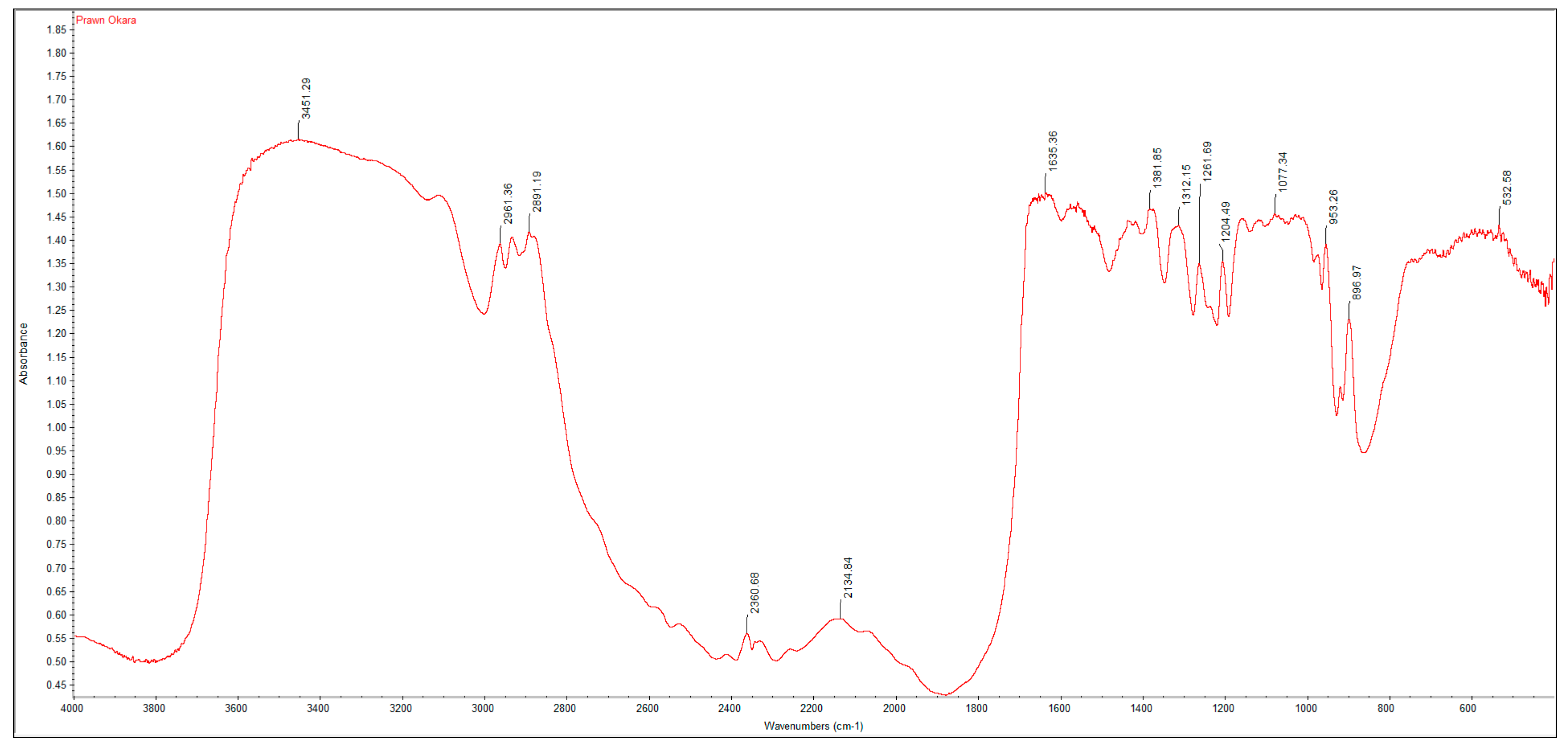
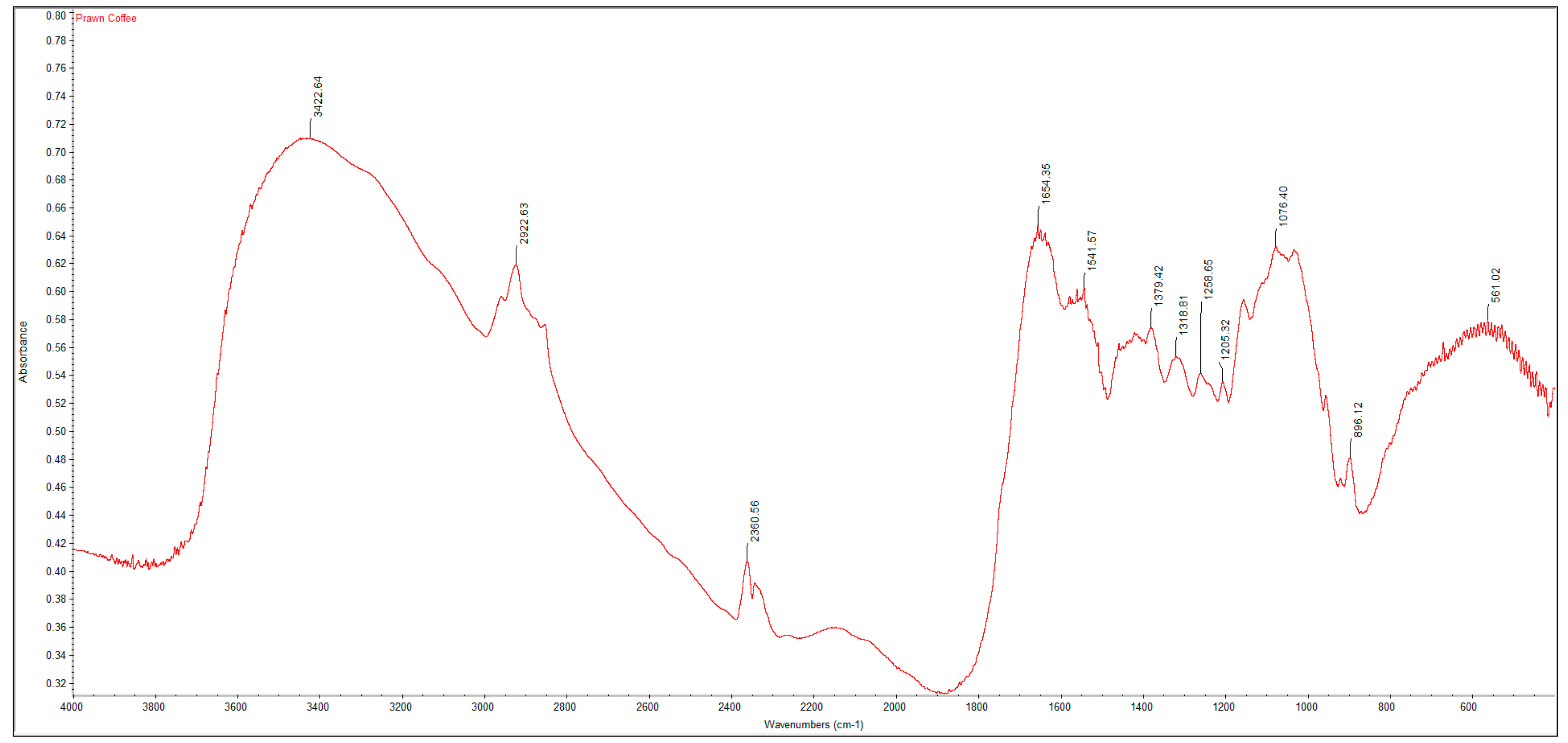
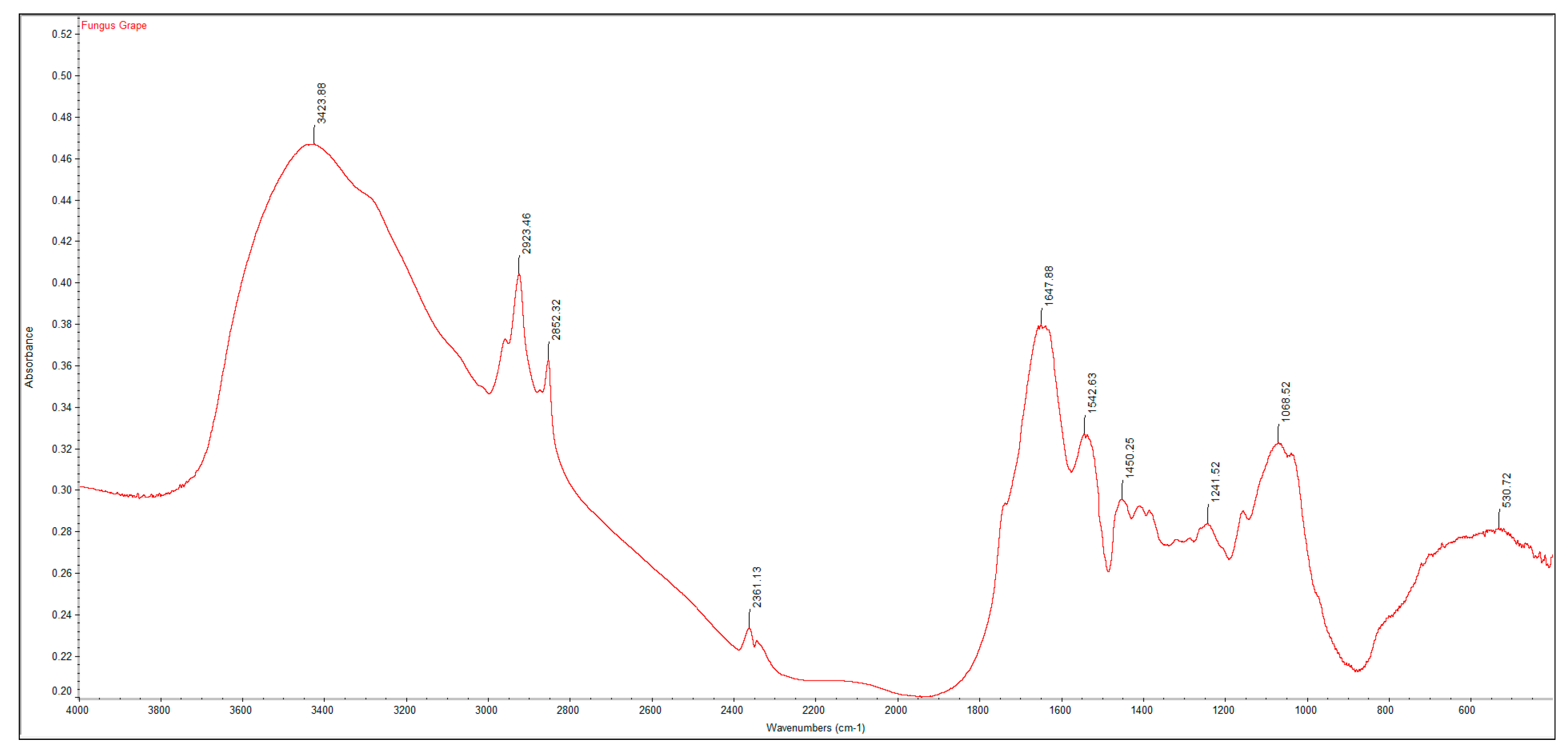
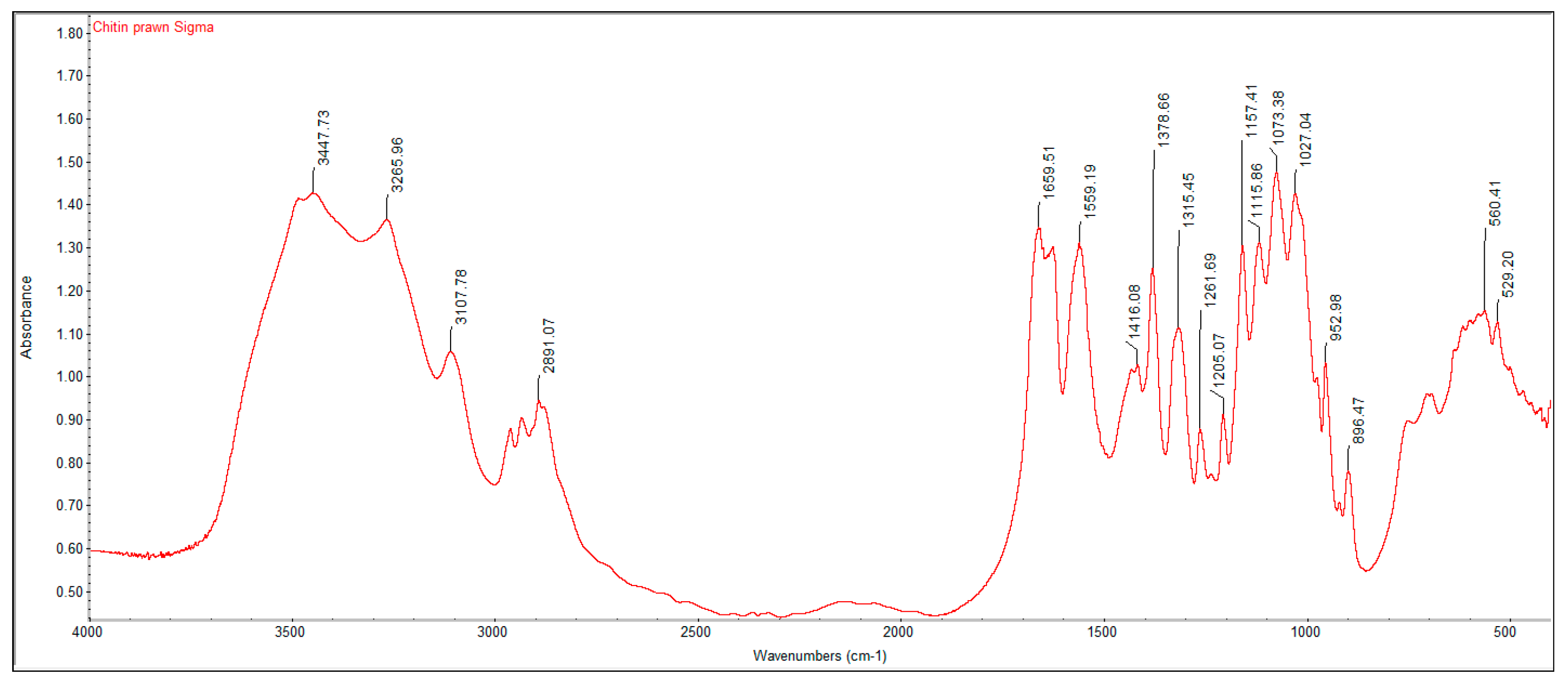
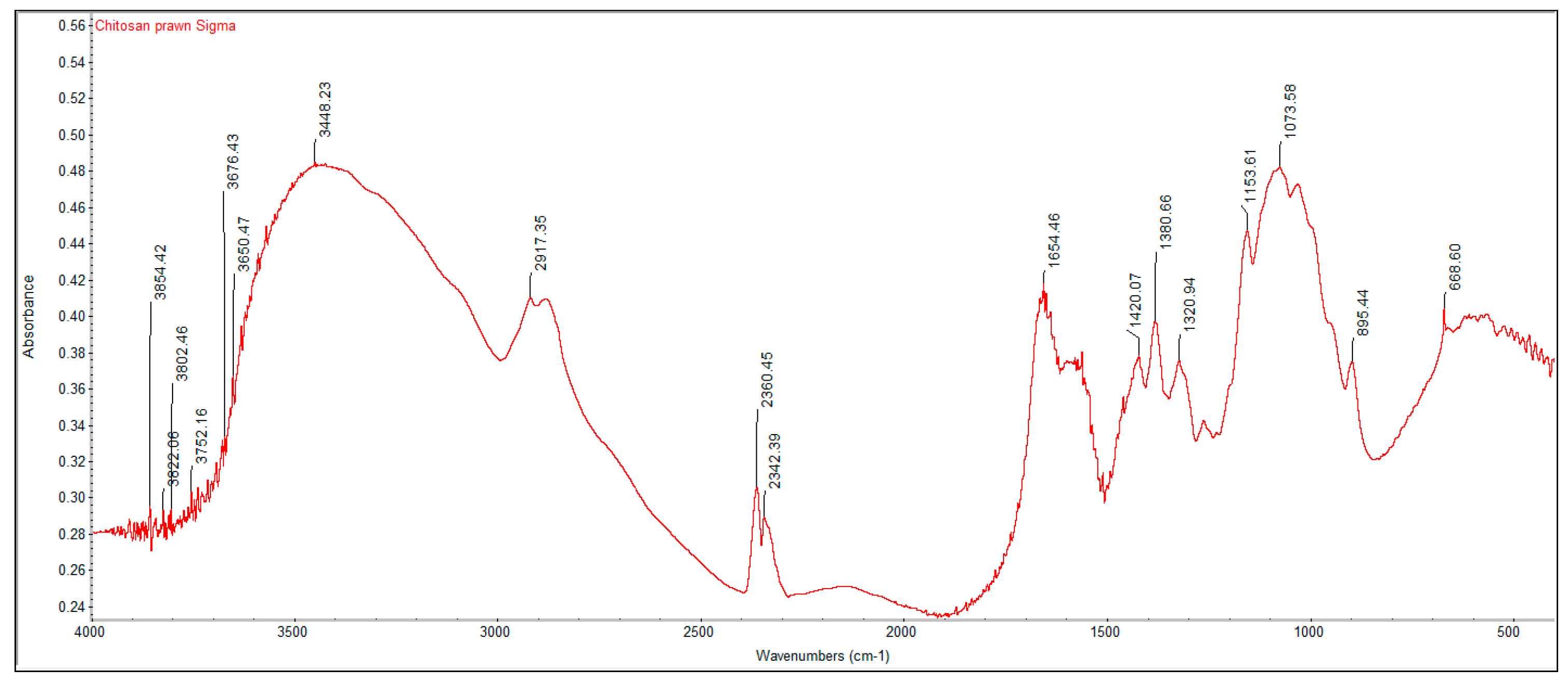
2.2. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
A PerkinElmer Spectrum One Fourier Transform Infrared Spectroscopy (FTIR) was used to characterize the crustacean chitin and fungal chitosan samples. The samples were ground with dried potassium bromide (KBr) and a pressure of 10 tons was applied for 2 min to form a disc. Analysis was performed across the range 4000–500 cm
−1. The degree of deacetylation (DD%) was calculated using the formula:
where “A” represents the absorbance of the respective wavenumbers 1650 and 3450 cm
−1. The amide-I band (1650 cm
−1) was used as the analytical band and the hydroxyl band (3450 cm
−1) as the internal reference band. The factor “1.33” denotes the value of the ratio of A
1650/A
3450 for fully
N-acetylated chitin [
21].
2.3. Elemental Analysis (EA)
An Elementar Vario EL III Elemental Analyzer was used to determine the carbon/nitrogen ratio of the crustacean chitin and fungal chitosan extracted. A total of 5 mg of each sample was added to a tin foil boat, which was folded into a pellet and loaded into a sample carousel. The samples were then dropped inside the instrument, where catalytic tube combustion occurred in an oxygenated, high temperature CO2 atmosphere. The carbon and nitrogen components were carried by helium through specific adsorption columns to separate them, after which their respective concentrations were determined using a thermal conductivity detector.
2.4. Statistical Analysis
All measurements were taken in triplicate and the results were expressed as the mean ± standard deviation.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}