Physicochemical, Microbiological and Functional Properties of Camelina Meal Fermented in Solid-State Using Food Grade Aspergillus Fungi

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Samples
2.2.1. Preparation of Fungi Inocula
2.2.2. Solid-State Fermentation
2.3. pH, Total Titratable Acidity and Colour Attributes
2.4. Proximate Analysis
2.5. Anti-Nutritional Factors
2.5.1. Phytic Acid
2.5.2. Total Glucosinolates
2.6. Total Phenolic Contents
2.7. Fibre Fractions
2.8. In Vitro Enzyme Protein Digestion (IVPD)
2.9. Microbiological Analysis
2.10. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.11. Functional Properties
2.11.1. Bulk Density
2.11.2. Water Absorption Capacity
2.11.3. Oil Absorption Capacity
2.11.4. Swelling Index
2.11.5. Swelling Capacity
2.11.6. Dispersibility
2.12. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition
3.2. pH, Total Titratable Acidity and Colour Attributes
3.3. Fibre Fractions
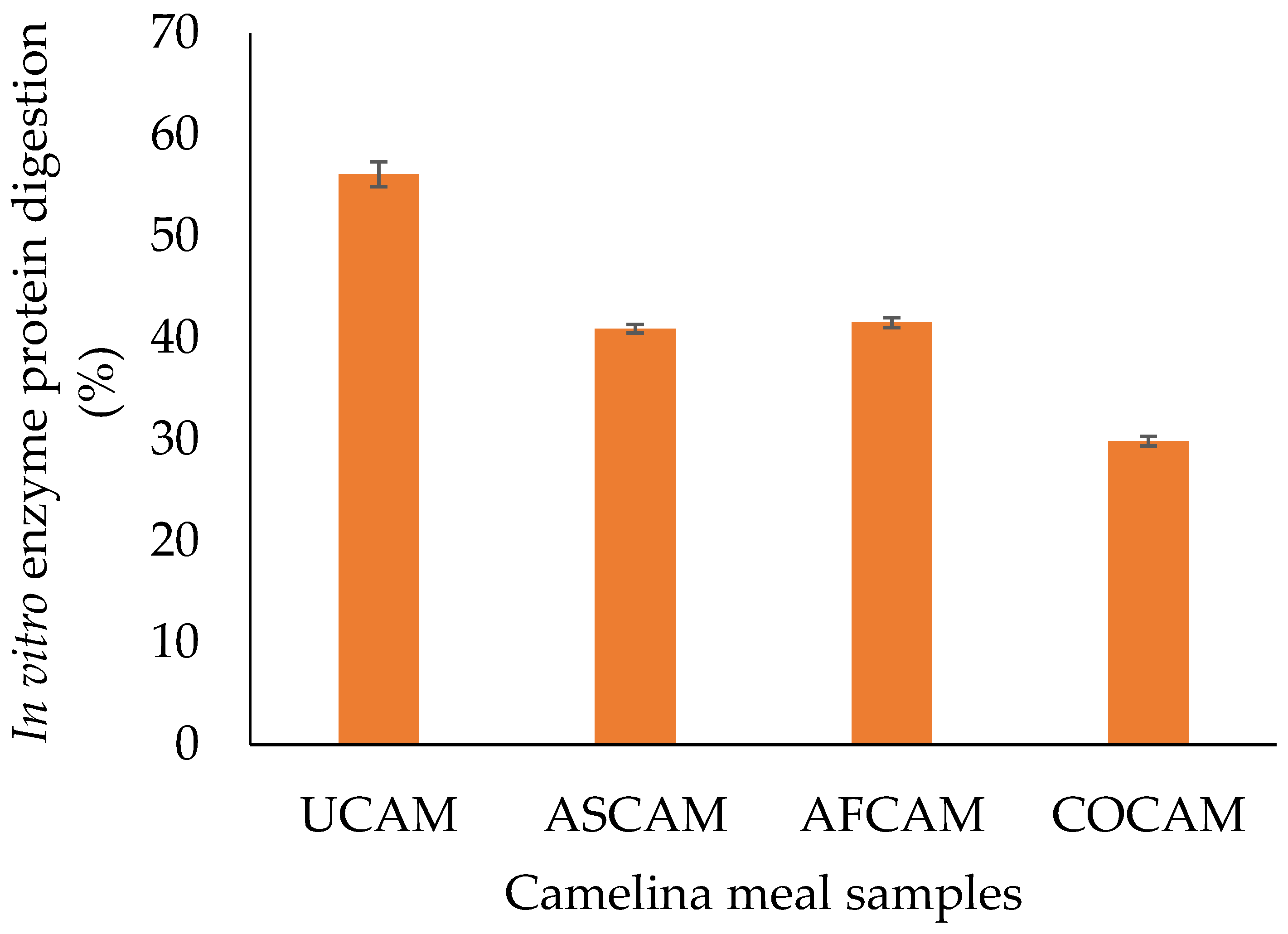
3.4. In Vitro Enzyme Protein Digestion (IVPD)
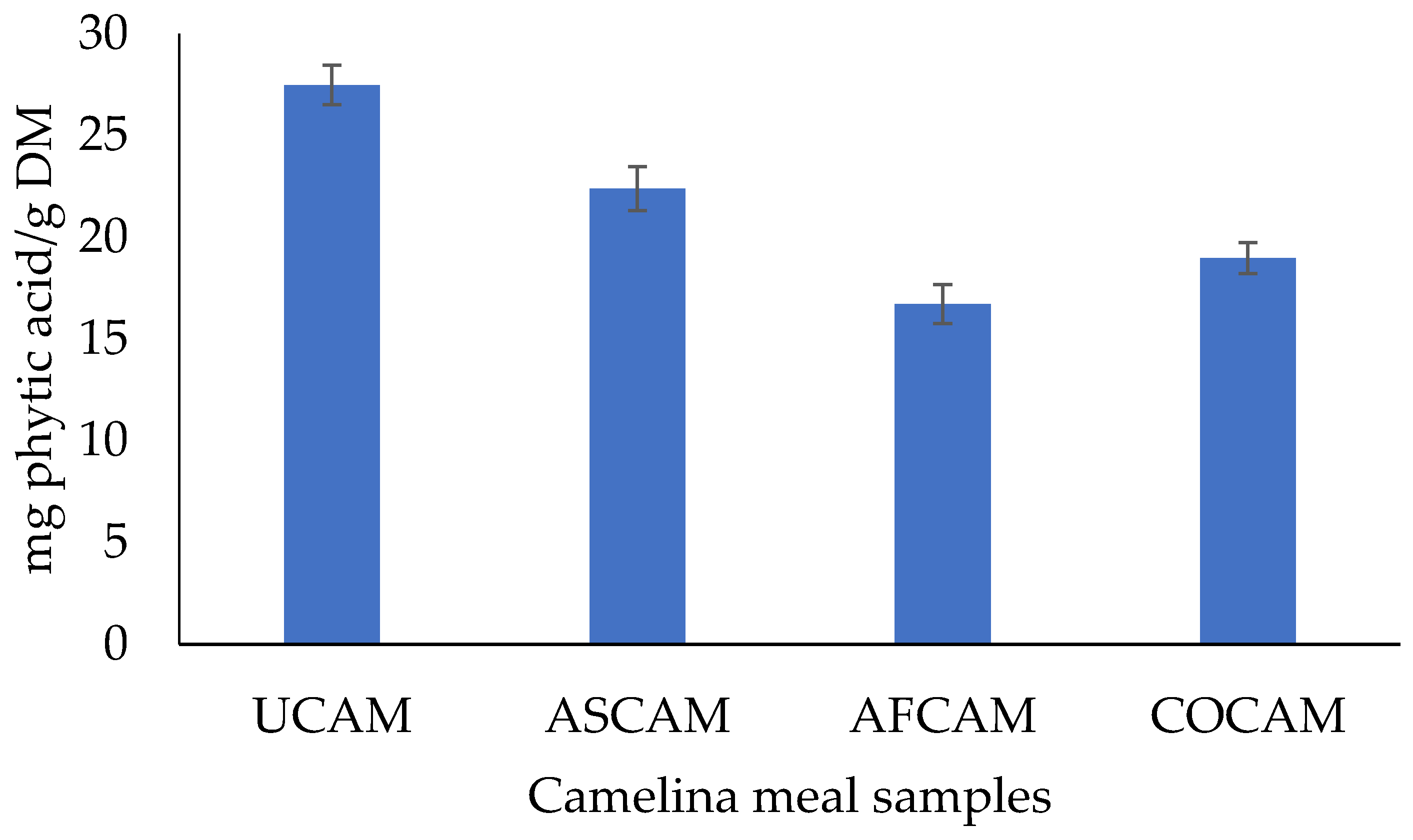
3.5. Total Glucosinolates, Total Phenolic Contents and Phytic Acid
3.6. Microbiological Composition
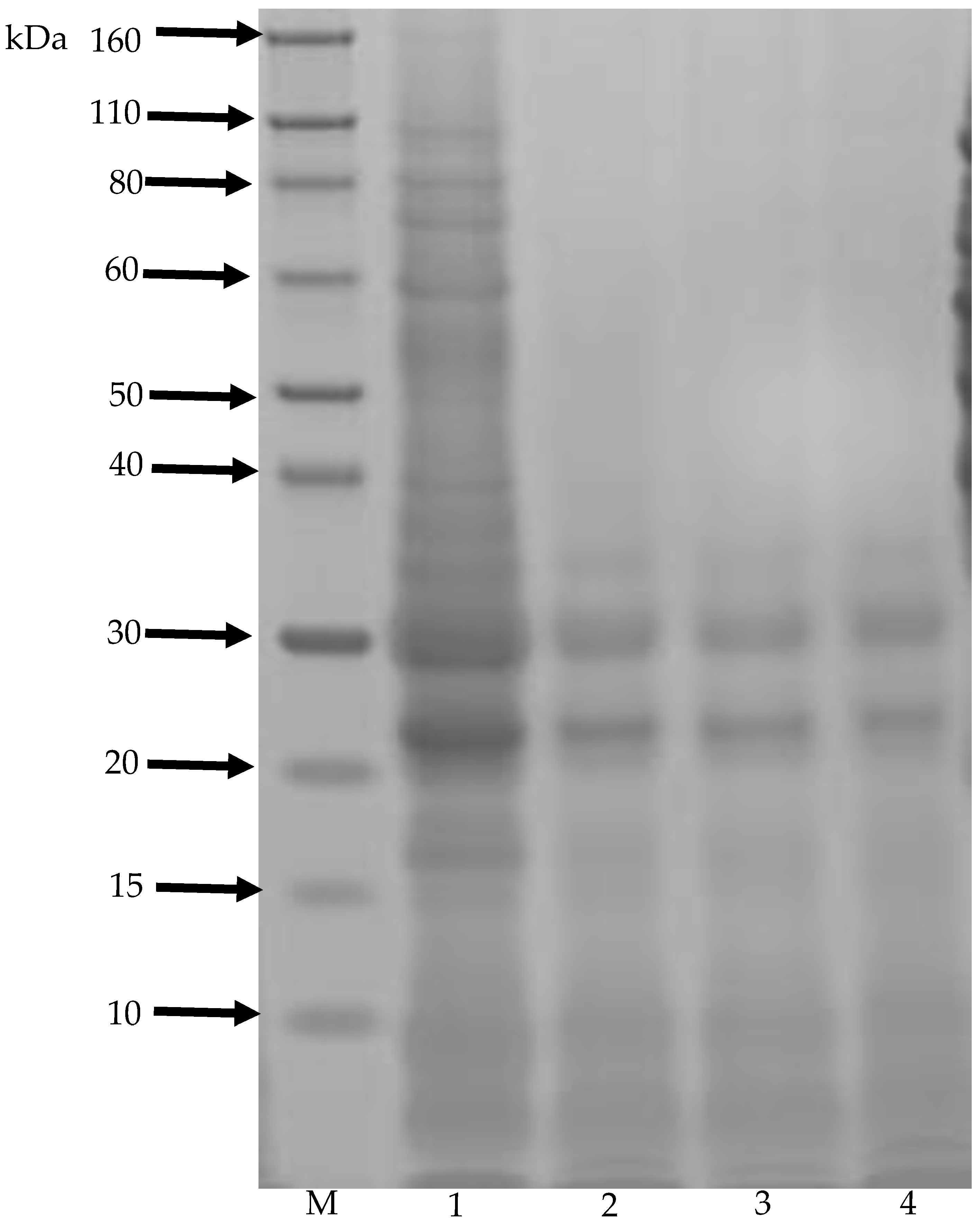
3.7. SDS-PAGE Analysis
3.8. Functional Properties
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wittkop, B.; Snowdon, R.J.; Friedt, W. Status and perspectives of breeding for enhanced yield and quality of oilseed crops for Europe. Euphytica 2009, 170, 131–140. [Google Scholar] [CrossRef]
- Das, N.; Berhow, M.A.; Angelino, D.; Jeffery, E.H. Camelina sativa defatted seed meal contains both alkyl sulfinyl glucosinolates and quercetin that synergize bioactivity. J. Agric. Food Chem. 2014, 62, 8385–8391. [Google Scholar] [CrossRef] [PubMed]
- Aziza, A.E.; Quezada, N.; Cherian, G. Antioxidative effect of dietary camelina meal in fresh, stored, or cooked broiler chicken meat. Poult. Sci. 2010, 89, 2711–2718. [Google Scholar] [CrossRef] [PubMed]
- Boyle, C.; Hansen, L.; Hinnenkamp, C.; Ismail, B.P. Emerging camelina protein: Extraction, modification, and structural/functional characterization. J. Am. Oil Chem. Soc. 2018, 95, 1049–1062. [Google Scholar] [CrossRef]
- Russo, R.; Reggiani, R. Antinutritive compounds in twelve Camelina sativa genotypes. Am. J. Plant Sci. 2012, 3, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Thacker, P.; Widyaratne, G. Effects of expeller pressed camelina meal and/or canola meal on digestibility, performance and fatty acid composition of broiler chickens fed wheat–soybean meal-based diets. Arch. Anim. Nutr. 2012, 66, 402–415. [Google Scholar] [CrossRef]
- Brandao, V.L.N.; Silva, L.G.; Paula, E.M.; Monteiro, H.F.; Dai, X.; Lelis, A.L.J.; Faccenda, A.; Poulson, S.R.; Faciola, A.P. Effects of replacing canola meal with solvent-extracted camelina meal on microbial fermentation in a dual-flow continuous culture system. J. Dairy Sci. 2018, 101, 9028–9040. [Google Scholar] [CrossRef]
- Pekel, A.Y.; Horn, N.L.; Adeola, O. The efficacy of dietary xylanase and phytase in broiler chickens fed expeller-extracted camelina meal. Poult. Sci. 2017, 96, 98–107. [Google Scholar] [CrossRef]
- Ajila, C.M.; Brar, S.K.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Solid-state fermentation of apple pomace using Phanerocheate chrysosporium–liberation and extraction of phenolic antioxidants. Food Chem. 2011, 126, 1071–1080. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Y.; Li, A.; Wang, Z.; Zhang, X.; Yun, T.; Qui, L.; Yin, Y. Effects of fermented rapeseed meal on antioxidant functions, serum biochemical parameters and intestinal morphology in broilers. Food Agric. Immunol. 2016, 27, 182–193. [Google Scholar] [CrossRef]
- Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Nutritional analysis of solid-state fermented canola meal (an improved protein source for broilers). In Proceedings of the 22nd European Symposium on Poultry Nutrition, Gdansk, Poland, 10–13 June 2019; p. 167. [Google Scholar]
- Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Solid-state fermented plant protein sources in the diets of broiler chickens: A review. Anim. Nutr. 2019, 5, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Aljuobori, A.; Idrus, Z.; Soleimani, A.F.; Abdullah, N.; Juan Boo, L. Response of broiler chickens to dietary inclusion of fermented canola meal under heat stress condition. Ital. J. Anim. Sci. 2017, 16, 546–551. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, R.L.; Ren, Z.Q.; Fan, Y.W.; Hu, S.B.; Zhuo, C.F.; Deng, Z.Y. Improvement of protein quality and degradation of allergen in soybean meal fermented by Neurospora crassa. LWT-Food Sci. Technol. 2019, 101, 220–228. [Google Scholar] [CrossRef]
- Li, J.; Sun, H.; Nie, C.; Ge, W.; Wang, Y.; Zhang, W. Oligopeptide derived from solid-state fermented cottonseed meal significantly affect the immunomodulatory in BALB/c mice treated with cyclophosphamide. Food Sci. Biotechnol. 2018, 27, 1791–1799. [Google Scholar] [CrossRef] [Green Version]
- Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Zhang, D.; Li, X.; Sultanbawa, Y. Phytic acid reduction in canola and camelina meals by fungal fermentation for potential broiler feeding. In Proceedings of the 30th Annual Australian Poultry Science Symposium, Sydney, Australia, 17–20 February 2019; p. 203. [Google Scholar]
- Olukomaiya, O.O.; Adiamo, O.Q.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Effect of solid-state fermentation on proximate composition, anti-nutritional factor, microbiological and functional properties of lupin flour. Food Chem. 2020, 315, 126238. [Google Scholar] [CrossRef]
- Heerd, D.; Yegin, S.; Tari, C.; Fernandez-Lahore, M. Pectinase enzyme-complex production by Aspergillus spp. in solid-state fermentation: A comparative study. Food Bioprod. Process. 2012, 90, 102–110. [Google Scholar] [CrossRef]
- Shin, H.Y.; Kim, S.M.; Lee, J.H.; Lim, S.T. Solid-state fermentation of black rice bran with Aspergillus awamori and Aspergillus oryzae: Effects on phenolic acid composition and antioxidant activity of bran extracts. Food Chem. 2019, 272, 235–241. [Google Scholar] [CrossRef]
- Nair, V.C.; Duvnjak, Z. Phytic acid content reduction in canola meal by various microorganisms in a solid-state fermentation process. Acta Biotechnol. 1991, 11, 211–218. [Google Scholar] [CrossRef]
- Chen, L.; Vadlani, P.V.; Mall, R.L. High-efficiency removal of phytic acid in soy meal using two-stage temperature-induced Aspergillus oryzae solid-state fermentation. J. Sci. Food Agric. 2014, 94, 113–118. [Google Scholar] [CrossRef]
- Chen, L.; Vadlani, P.V.; Madl, R.L.; Gibbons, W. Degradation of phytic acid and soy protein in soy meal via co-fermentation of Aspergillus oryzae and Aspergillus ficuum. J. Am. Oil Chem. Soc. 2016, 93, 45–50. [Google Scholar] [CrossRef]
- Li, N.; Qi, G.; Sun, X.S.; Xu, F.; Wang, D. Adhesion properties of camelina protein fractions isolated with different methods. Ind. Crops Prod. 2015, 69, 263–272. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, D.; Sun, X.S. Physico-chemical properties of camelina protein altered by sodium bisulfite and guanidine-HCl. Ind. Crops Prod. 2016, 83, 453–461. [Google Scholar] [CrossRef]
- Rahman, M.J.; de Camargo, A.C.; Shahidi, F. Phenolic profiles and antioxidant activity of defatted camelina and sophia seeds. Food Chem. 2018, 240, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Krungleviciute, V.; Juodeikiene, G.; Vidmantiene, D.; Maknickiene, Z. Solid-state fermentation with lactic acid bacteria to improve the nutritional quality of lupin and soya bean. J. Sci. Food Agric. 2015, 95, 1336–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Karkalas, J. An improved enzymic method for the determination of native and modified starch. J. Sci. Food Agric. 1985, 36, 1019–1027. [Google Scholar] [CrossRef]
- Dolan, S.P.; Nortrup, D.A.; Bolger, P.M.; Capar, S.G. Analysis of dietary supplements for arsenic, cadmium, mercury, and lead using inductively coupled plasma mass spectrometry. J. Agric. Food Chem. 2003, 51, 1307–1312. [Google Scholar] [CrossRef]
- Gao, Y.; Shang, C.; Maroof, M.A.; Biyashev, R.M.; Grabau, E.A.; Kwanyuen, P.; Burton, J.W.; Buss, G.R. A modified colorimetric method for phytic acid analysis in soybean. Crop Sci. 2007, 47, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- Gallaher, C.M.; Gallaher, D.D.; Peterson, S. Development and validation of a spectrophotometric method for quantification of total glucosinolates in cruciferous vegetables. J. Agric. Food Chem. 2012, 60, 1358–1362. [Google Scholar] [CrossRef]
- Mawlong, I.; Sujith Kumar, M.S.; Gurung, B.; Singh, K.H.; Singh, D. A simple spectrophotometric method for estimating total glucosinolates in mustard de-oiled cake. Int. J. Food Prop. 2017, 20, 3274–3281. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Van Soest, P.J. Use of detergents in the analysis of fibrous feeds. 1. Preparation of fiber residues of low nitrogen content. J. Assoc. Off. Agric. Chem. 1963, 46, 825–829. [Google Scholar]
- Van Soest, P.J. Use of detergents in the analysis of fibrous feeds. 2. A rapid method for the determination of fiber and lignin. J. Assoc. Off. Agric. Chem. 1963, 46, 829–835. [Google Scholar]
- Nalle, C.L. Nutritional Evaluation of Grain Legumes for Poultry. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2009; p. 183. [Google Scholar]
- Kasprowicz-Potocka, M.; Zaworska, A.; Gulewicz, P.; Nowak, P.; Frankiewicz, A. The effect of fermentation of high alkaloid seeds of Lupinus angustifolius var. Karo by Saccharomyces cerevisieae, Kluyveromyces lactis, and Candida utilis on the chemical and microbial composition of products. J. Food Process. Preserv. 2018, 42, e13487. [Google Scholar] [CrossRef]
- Oladele, A.K.; Aina, J.O. Chemical composition and functional properties of flour produced from two varieties of tigernut (Cyperus esculentus). Afr. J. Biotechnol. 2007, 6, 2473–2476. [Google Scholar] [CrossRef] [Green Version]
- Diniz, F.M.; Martin, A.M. Effects of the extent of enzymatic hydrolysis on functional properties of shark protein hydrolysate. LWT-Food Sci. Technol. 1997, 30, 266–272. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Ukpabi, U.J.; Ndimele, C. Evaluation of the quality of gari produced in Imo State. Nig. Food J. 1990, 8, 105–110. [Google Scholar]
- Okaka, J.C.; Potter, N.N. Functional and storage properties of cowpea powder-wheat flour blends in bread making. J. Food Sci. 1977, 42, 828–833. [Google Scholar] [CrossRef]
- Kulkarni, K.D.; Kulkarni, D.N.; Ingle, U.M. Sorghum malt-based weaning food formulations: Preparation, functional properties and nutritive value. Food Nutr. Bull. 1991, 13, 1–7. [Google Scholar] [CrossRef]
- Plaipetch, P.; Yakupitiyage, A. Effect of replacing soybean meal with yeast-fermented canola meal on growth and nutrient retention of Nile tilapia, Oreochromis niloticus (Linnaeus 1758). Aquac. Res. 2014, 45, 1744–1753. [Google Scholar] [CrossRef]
- Shi, C.; He, J.; Yu, J.; Yu, B.; Mao, X.; Zheng, P.; Huang, Z.; Chen, D. Physicochemical properties analysis and secretome of Aspergillus niger in fermented rapeseed meal. PLoS ONE 2016, 11, e0153230. [Google Scholar] [CrossRef]
- Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Solid-state fermentation of canola meal with Aspergillus sojae, Aspergillus ficuum and their co-cultures: Effects on physicochemical, microbiological and functional properties. LWT-Food Sci. Technol. 2020, 127, 109362. [Google Scholar] [CrossRef]
- Kaczmarska, K.T.; Chandra-Hioe, M.V.; Zabaras, D.; Frank, D.; Arcot, J. Effect of germination and fermentation on carbohydrate composition of Australian sweet lupin and soybean seeds and flours. J. Agric. Food Chem. 2017, 65, 10064–10073. [Google Scholar] [CrossRef] [PubMed]
- Granito, M.; Frias, J.; Doblado, R.; Guerra, M.; Champ, M.; Vidal-Valverde, C. Nutritional improvement of beans (Phaseolus vulgaris) by natural fermentation. Eur. Food Res. Technol. 2002, 214, 226–231. [Google Scholar] [CrossRef]
- Canibe, N.; Miettinen, H.; Jensen, B.B. Effect of adding Lactobacillus plantarum or a formic acid containing-product to fermented liquid feed on gastrointestinal ecology and growth performance of piglets. Livest. Sci. 2008, 114, 251–262. [Google Scholar] [CrossRef]
- Wakil, S.M.; Onilude, A.A. Microbiological and chemical changes during production of malted and fermented cereal-legume weaning foods. Adv. Food Sci. 2009, 31, 139–145. [Google Scholar]
- Mbata, T.I.; Ikenebomeh, M.J.; Alaneme, J.C. Studies on the microbiological, nutrient composition and antinutritional contents of fermented maize flour fortified with bambara groundnut (Vigna subterranean L). Afr. J. Food Sci. 2009, 3, 165–171. [Google Scholar]
- Bilgiçli, N.; İbanoğlu, Ş. Effect of wheat germ and wheat bran on the fermentation activity, phytic acid content and colour of tarhana, a wheat flour–yoghurt mixture. J. Food Eng. 2007, 78, 681–686. [Google Scholar] [CrossRef]
- Chen, L.; Vadlani, P.V.; Madl, R.L.; Wang, W.; Shi, Y.; Gibbons, W.R. The investigation of virginiamycin-added fungal fermentation on the size and immunoreactivity of heat-sensitive soy protein. Int. J. Polym. Sci. 2015, 2015, 682596. [Google Scholar] [CrossRef]
- Ramli, M.N.; Imura, Y.; Takayama, K.; Nakanishi, Y. Bioconversion of sugarcane bagasse with Japanese koji by solid-state fermentation and its effects on nutritive value and preference in goats. Asian-Australas. J. Anim. Sci. 2005, 18, 1279–1284. [Google Scholar] [CrossRef]
- Ogodo, A.C.; Ugbogu, O.C.; Onyeagba, R.A.; Okereke, H.C. In vitro starch and protein digestibility and proximate composition of soybean flour fermented with lactic acid bacteria (LAB) consortia. Agric. Nat. Resour. 2018, 52, 503–509. [Google Scholar] [CrossRef]
- Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Nutritional composition of solid-state fermented camelina meal (an enriched protein source for broiler chickens). Proceedings 2019, 36, 106. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.M.; Guo, X.N.; Zhu, K.X. Impact of solid-state fermentation on nutritional, physical and flavor properties of wheat bran. Food Chem. 2017, 217, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.G.; Porres, J.M. Phytase enzymology, applications and biotechnology. Biotechnol. Lett. 2003, 25, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Vats, P.; Banerjee, U.C. Production studies and catalytic properties of phytases (myo-inositol hexakisphosphate phosphohydrolases): An overview. Enzym. Microb. Technol. 2004, 35, 3–14. [Google Scholar] [CrossRef]
- Achi, O.K. Traditional fermented protein condiments in Nigeria. Afr. J. Biotechnol. 2005, 4, 1612–1621. [Google Scholar]
- Kasprowicz-Potocka, M.; Zaworska, A.; Frankiewicz, A.; Nowak, W.; Gulewicz, P.; Zduńczyk, Z.; Juśkiewicz, J. The nutritional value and physiological properties of diets with raw and Candida utilis-fermented lupin seeds in rats. Food Technol. Biotechnol. 2015, 53, 286–297. [Google Scholar] [CrossRef]
- Bérot, S.; Compoint, J.P.; Larré, C.; Malabat, C.; Guéguen, J. Large scale purification of rapeseed proteins (Brassica napus L.). J. Chromatogr. B 2005, 818, 35–42. [Google Scholar] [CrossRef]
- Russo, R. Biochemical Characterization of Flour from Seeds of Camelina sativa L. (Crantz) after Chemical Extraction of Oil. Ph.D. Thesis, University of Milan (Università Degli Studi di Milano), Milan, Italy, 2012; pp. 37–39. [Google Scholar]
- Osungbade, O.R.; Gbadamosi, O.S.; Adiamo, O.Q. Effects of cooking and fermentation on the chemical composition, functional properties and protein digestibility of sandbox (Hura crepitans) seeds. J. Food Biochem. 2016, 40, 754–765. [Google Scholar] [CrossRef]
- Fagbemi, T.N. Effect of blanching and ripening on functional properties of plantain (Musa aab) flour. Plant Food Hum. Nutr. 1999, 54, 261–269. [Google Scholar] [CrossRef]
- Adebowale, O.J.; Maliki, K. Effect of fermentation period on the chemical composition and functional properties of Pigeon pea (Cajanus cajan) seed flour. Int. Food Res. J. 2011, 18, 1329–1333. [Google Scholar]
- Afoakwa, E.O.; Sefa-Dedeh, S.; Budu, A.S.; Sakyi-Dawson, E.; Asomaning, J. Influence of spontaneous fermentation on some quality characteristics of maize-based cowpea-fortified nixtamalized foods. Afr. J. Food Agric. Nutr. Dev. 2007, 7, 1–18. [Google Scholar]
- Igbabul, B.; Hiikyaa, O.; Amove, J. Effect of fermentation on the proximate composition and functional properties of mahogany bean (Afzelia africana) flour. Curr. Res. Nutr. Food Sci. 2014, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Sun, M.; Zhang, Q.; Chen, Y.; Miao, J.; Rui, X.; Dong, M. Effects of Cordyceps militaris (L.) Fr. fermentation on the nutritional, physicochemical, functional properties and angiotensin I converting enzyme inhibitory activity of red bean (Phaseolus angularis [Willd.] WF Wight.) flour. J. Food Sci. Technol. 2018, 55, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Samsher, S. Assessment of functional properties of different flours. Afr. J. Agric. Res. 2013, 8, 4849–4852. [Google Scholar]
- Ilowefah, M.; Bakar, J.; Ghazali, H.M.; Mediani, A.; Muhammad, K. Physicochemical and functional properties of yeast fermented brown rice flour. J. Food Sci. Technol. 2015, 52, 5534–5545. [Google Scholar] [CrossRef] [Green Version]
- Adebiyi, J.A.; Obadina, A.O.; Mulaba-Bafubiandi, A.F.; Adebo, O.A.; Kayitesi, E. Effect of fermentation and malting on the microstructure and selected physicochemical properties of pearl millet (Pennisetum glaucum) flour and biscuit. J. Cereal Sci. 2016, 70, 132–139. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | UCAM | ASCAM | AFCAM | COCAM |
|---|---|---|---|---|
| Crude protein (%) | 37.03 ± 0.06 a | 37.57 ± 0.06 a | 37.77 ± 0.06 a | 37.81 ± 0.01 a |
| Crude fat (%) | 15.63 ± 0.04 a | 15.47 ± 0.07 a | 15.18 ± 0.10 a | 15.31 ± 0.03 a |
| Crude ash (%) | 5.92 ± 0.19 a | 5.90 ± 0.14 a | 5.90 ± 0.06 a | 5.81 ± 0.13 a |
| Soluble carbohydrate (glucose, %) | 8.71 ± 0.02 a | 6.35 ± 0.03 c | 6.07 ± 0.05 d | 6.46 ± 0.03 b |
| Starch (%) | 0.18 ± 0.02 c | 1.13 ± 0.06 a | 0.94 ± 0.03 b | 1.03 ± 0.06 ab |
| Calcium (%) | 0.22 ± 0.10 a | 0.31 ± 0.26 a | 0.24 ± 0.16 a | 0.21 ± 0.10 a |
| Phosphorus (%) | 0.77 ± 0.13 a | 0.86 ± 0.28 a | 0.82 ± 0.21 a | 0.78 ± 0.14 a |
| Parameters | UCAM | ASCAM | AFCAM | COCAM | CV (%) |
|---|---|---|---|---|---|
| pH | 6.07 ± 0.01 a | 6.03 ± 0.01 b | 5.87 ± 0.01 d | 5.94 ± 0.01 c | 0.17 |
| Total titratable acidity | 0.45 ± 0.14 b | 1.26 ± 0.12 a | 0.95 ± 0.09 a | 1.13 ± 0.31 a | 19.39 |
| Neutral detergent fibre (%) | 33.49 ± 2.41 a | 36.94 ± 2.51 a | 38.48 ± 2.84 a | 40.36 ± 3.44 a | 7.47 |
| Acid detergent fibre (%) | 16.86 ± 0.04 b | 20.76 ± 0.67 a | 21.47 ± 1.98 a | 19.96 ± 0.76 a | 4.12 |
| Cellulose (%) | 12.25 ± 0.51 b | 14.79 ± 0.64 a | 15.44 ± 1.49 a | 14.48 ± 0.39 ab | 5.21 |
| Hemicellulose (%) | 16.63 ± 2.37 a | 16.18 ± 1.84 a | 17.01 ± 0.86 a | 20.40 ± 2.68 a | 10.95 |
| Lignin (%) | 4.60 ± 0.55 b | 5.97 ± 0.03 a | 6.03 ± 0.49 a | 5.48 ± 0.37 ab | 6.83 |
| Colour | |||||
| L | 47.71 ± 0.71 a | 44.84 ± 1.05 b | 42.81 ± 1.04 b | 43.72 ± 1.29 b | 2.30 |
| a | 29.92 ± 0.37 a | 24.78 ± 0.33 b | 24.42 ± 0.64 b | 25.07 ± 0.08 b | 1.38 |
| b | 73.76 ± 0.39 a | 71.71 ± 0.45 b | 70.46 ± 0.43 c | 70.66 ± 0.43 bc | 0.59 |
| Parameters | Total Glucosinolates (µmol SE/g DM) | Total Phenolic Contents (mg GAE/g DM) |
|---|---|---|
| UCAM | 30.35 ± 2.33 a | 2.57 ± 0.17 a |
| ASCAM | 26.48 ± 0.58 a | 2.64 ± 0.27 a |
| AFCAM | 28.80 ± 5.12 a | 2.64 ± 0.17 a |
| COCAM | 26.16 ± 0.21 a | 2.52 ± 0.10 a |
| CV (%) | 7.11 | 6.81 |
| Parameters | UCAM | ASCAM | AFCAM | COCAM |
|---|---|---|---|---|
| Total bacteria (log CFU/g) | 2.00 ± 0.27 c | 5.28 ± 0.16 a | 4.76 ± 0.21 a | 3.00 ± 0.52 b |
| Lactic acid bacteria (log CFU/g) | 1.10 ± 0.33 c | 3.37 ± 0.28 b | 4.50 ± 0.50 a | 3.93 ± 0.40 ab |
| Salmonella (/25 g) | ND | ND | ND | ND |
| Molds (log CFU/g) | 1.70 ± 0.50 b | 4.30 ± 0.20 a | 3.78 ± 0.33 a | 2.70 ± 0.22 b |
| Yeasts (log CFU/g) | 1.50 ± 0.32 b | 3.15 ± 0.51 a | 3.48 ± 0.19 a | 3.52 ± 0.25 a |
| Parameters | UCAM | ASCAM | AFCAM | COCAM | CV (%) |
|---|---|---|---|---|---|
| Bulk density (g/mL) | 0.53 ± 0.01 b | 0.56 ± 0.01 a | 0.56 ± 0.01 a | 0.57 ± 0.00 a | 1.36 |
| WAC (%) | 650 ± 110.00 a | 395 ± 25.00 b | 390 ± 0.00 b | 385 ± 55.00 b | 9.38 |
| OAC (%) | 100.00 ± 0.00 ab | 90.00 ± 10.00 b | 110.00 ± 10.00 a | 100.00 ± 0.00 ab | 5.05 |
| Swelling index | 3.19 ± 0.00 ab | 2.79 ± 0.13 b | 3.99 ± 0.67 a | 2.93 ± 0.00 b | 5.36 |
| Swelling capacity | 3.90 ± 0.10 b | 4.78 ± 0.22 a | 4.76 ± 0.17 a | 4.52 ± 0.04 a | 2.91 |
| Dispersibility (%) | 90.87 ± 0.18 a | 90.78 ± 0.35 a | 90.42 ± 0.09 a | 90.82 ± 0.31 a | 0.26 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Physicochemical, Microbiological and Functional Properties of Camelina Meal Fermented in Solid-State Using Food Grade Aspergillus Fungi. Fermentation 2020, 6, 44. https://doi.org/10.3390/fermentation6020044
Olukomaiya OO, Fernando WC, Mereddy R, Li X, Sultanbawa Y. Physicochemical, Microbiological and Functional Properties of Camelina Meal Fermented in Solid-State Using Food Grade Aspergillus Fungi. Fermentation. 2020; 6(2):44. https://doi.org/10.3390/fermentation6020044
Chicago/Turabian StyleOlukomaiya, Oladapo Oluwaseye, W. Chrishanthi Fernando, Ram Mereddy, Xiuhua Li, and Yasmina Sultanbawa. 2020. "Physicochemical, Microbiological and Functional Properties of Camelina Meal Fermented in Solid-State Using Food Grade Aspergillus Fungi" Fermentation 6, no. 2: 44. https://doi.org/10.3390/fermentation6020044
APA StyleOlukomaiya, O. O., Fernando, W. C., Mereddy, R., Li, X., & Sultanbawa, Y. (2020). Physicochemical, Microbiological and Functional Properties of Camelina Meal Fermented in Solid-State Using Food Grade Aspergillus Fungi. Fermentation, 6(2), 44. https://doi.org/10.3390/fermentation6020044