1. Introduction
The possibility of microbial synthesis of the bivalent alcohol 2,3-butanediol (2,3-BD) was investigated for the first time more than a century ago by Harden and Walpole in
Klebsiella pneumoniae and by Donker in
Paenibacillus polymyxa [
1,
2]. Although its future use was only speculative at that time, today, the 2,3-BD market is notably fast-growing and is expected to reach USD 220 million in 2027 [
3]. The most indispensable application of 2,3-BD is in the chemical, pharmaceutical and food industries, as a starting reagent in the production of rubber, solvents, varnishes, polyesters, polyurethanes and methacrylate. It is also a common constituent of liquid fuels, antifreeze, cosmetic products, drugs, antiperspirants and ointments [
4,
5,
6]. A recent study reports 2,3-BD integration into biodegradable biofilms [
7]. However, the emerging ecological problems, fossil fuel exhaustion and climate changes impose the development of biotechnologies for a bio-based alternative of 2,3-BD chemical synthesis despite cost competitiveness issues [
8,
9].
Current strategies for increased microbial production of 2,3-BD include (i) isolation of new bacterial producers, preferably non-pathogenic, that can utilize cheap, renewable and non-nutritional substrates, (ii) development of engineered strains that may produce optically pure isomers and (iii) selection of strains amenable to process optimization for increased 2,3-BD titer, yield and productivity. The “generally regarded as safe” (GRAS) 2,3-BD producers are the most desirable on an industrial scale; therefore,
Paenibacillus polymyxa [
10,
11],
Bacillus subtilis [
12,
13],
B. amyloliquefaciens [
14,
15],
B. pumilus,
B. siamensis [
16],
B. vallismortis [
17],
B. licheniformis,
B. velezensis,
B. toyonensis and
B. safensis have been evaluated as particularly promising [
18,
19,
20,
21].
However,
B. licheniformis combines the greatest number of benefits. It has the potential to convert a variety of substrates into 2,3-BD, such as starch and corncob hydrolyzates [
22,
23], inulin [
24] and most of the sugars included in plant biomass, such as glucose, cellobiose, galactose, mannose, xylose and arabinose. Although several attempts to optimize the medium content and culture conditions for
B. licheniformis have been made, several unresolved issues with the species and the peculiar nature of the process remain to be studied in more detail. First, several process optimizations refer to thermophilic
B. licheniformis strains [
16,
25], but information concerning the optimal conditions for high 2,3-BD production by mesophilic representatives is scarce. Second, it has to be noted that 2,3-BD is obtained via mixed acid fermentation yielding multiple products and the majority of them are undesirable and decrease the 2,3-BD yield. Beside 2,3-BD and its precursor, acetoin, some
B. licheniformis strains produce mainly acetic acid and ethanol as by-products [
26], while others secrete also lactate, formate and glycerol [
27]. Therefore, different optimization approaches are required to decrease the specific spectrum of final metabolites produced by each industrial strain.
On the other hand, oxygen supply is a critical factor for acetoin and 2,3-BD inter-conversion. High levels of dissolved oxygen lead to acetoin synthesis, while lower levels favor 2,3-BD synthesis [
21]. In addition, 2,3-BD can also serve as a carbon source for
B. licheniformis at diminished glucose concentrations [
28]. That is why a fed-batch process performance that requires additional improvement is frequently applied [
16,
20].
In our recent study, we revealed that Bulgarian isolate
B. licheniformis 24 possesses particularly high 2,3-BD productivity using glucose, mannose and cellobiose, reaching 0.77, 0.64 and 0.46 g/L/h, respectively [
18]. 2,3-BD yield from glucose was 83% of the theoretical maximum without any process optimization. Moreover, the strain was able to maintain low levels of acetoin even when the substrate was depleted and displayed significant natural extracellular cellulase activity [
18]. Since
B. licheniformis 24 is particularly promising as a producer of 2,3-BD from cellulose-containing substrates, we focused on detailed process optimization to obtain the highest production values from glucose. The purpose of the study was achieved by the use of Plackett–Burman and central composite design (CCD) for medium optimization and response surface design methodology for optimization of process parameters applied to both batch and fed-batch operation performance. The obtained record 2,3-BD amount also elucidates the maximal tolerance of the strain to this target metabolite.
4. Discussion
For more than a decade, the fermentation of 2,3-BD has been the subject of continuous laboratory research, including the use of different types of natural and modified producers, a variety of substrates, innovative methods to control the process and new, promising ways of product extraction [
5,
33]. At present, however, this process has never been successfully commercialized. The reasons lie in the high price of the substrates, the pathogenic nature of the best producers and the expensive extraction of the product from the fermentation mixture (further discouraged by significant losses).
B. licheniformis 24 has several advantages as a producer of 2,3-BD; it is non-pathogenic, has a broad substrate spectrum, possesses a high consumption rate and high yield, regardless of the substrate [
18]. The present study establishes the strain as a super-producer of 2,3-BD with a future industrial application by employing complex optimization via series of Plackett–Burman design and CCD experiments.
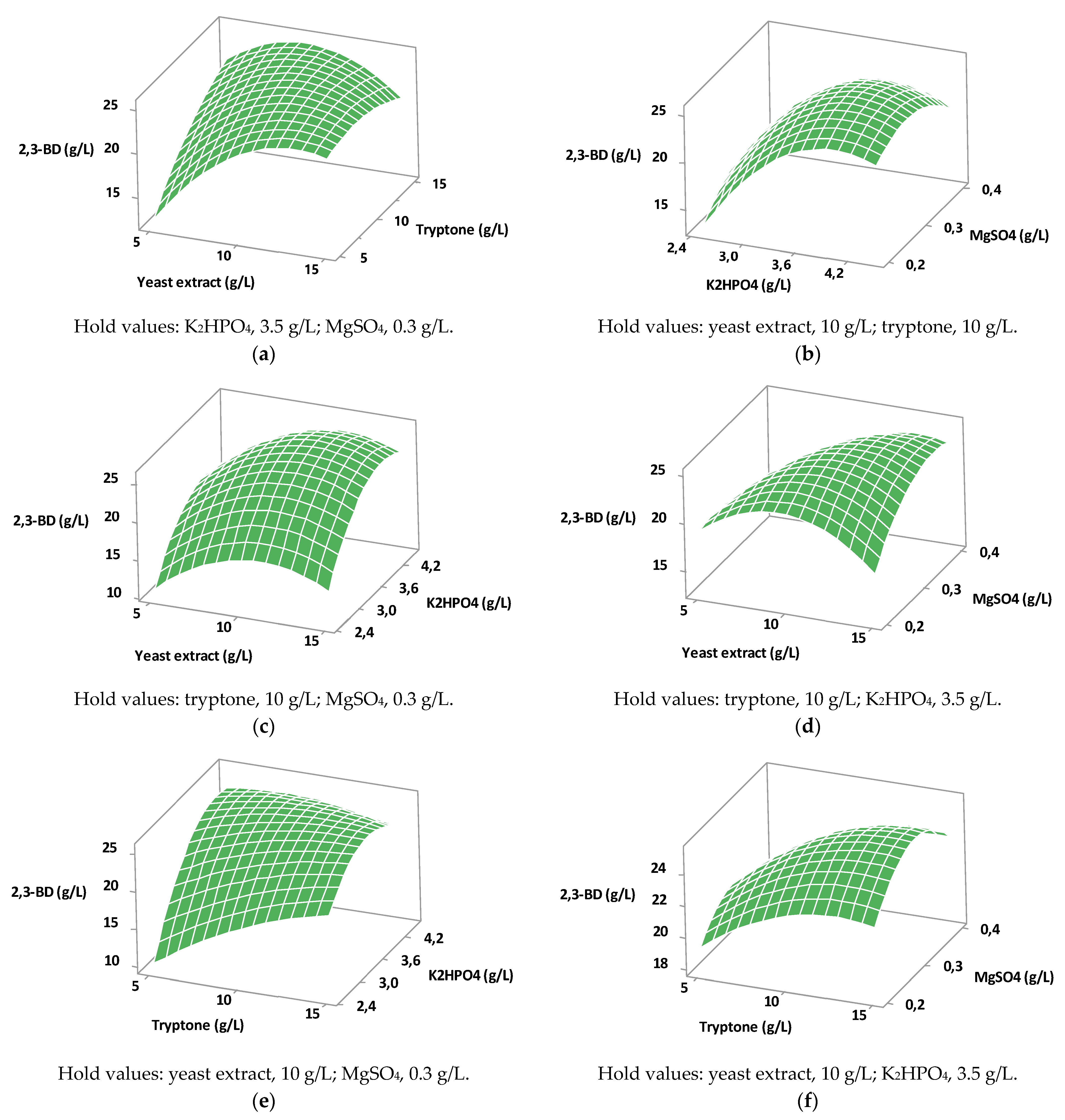
The application of the Plackett–Burman design showed that among the components of the nutrient medium with significant influence, the yeast extract, K
2HPO
4, MgSO
4 and tryptone had the most pronounced positive effect on 2,3-BD production. The addition of corn steep liquor and MOPS had a negative impact on 2,3-BD production and these two compounds were excluded from the media. This observation is in agreement with Song et al. [
16], who revealed that corn steep liquor is less suitable than yeast extract as a nitrogen source for
B. licheniformis. Unlike
P. polymyxa [
32],
B. licheniformis does not need MOPS, which makes the nutrient medium cheaper. The establishment of the optimal values of the components with the greatest impact on 2,3-BD production was achieved by CCD experiments. The computed optimal values were: yeast extract, 13.38 g/L; K
2HPO
4, 4.20 g/L; tryptone, 6.41 g/L; MgSO
4, 0.32 g/L. In this experiment, as revealed by the screening design, yeast extract and K
2HPO
4 had the highest impact on 2,3-BD production. However, MgSO
4 did not affect the response in the second experimental range. The response surface graph revealed that the yeast extract is the preferred nitrogen source, but a total concentration of yeast extract and tryptone above 20 g/L led to a decrease in 2,3-BD formation (
Figure 1a).
A specific characteristic feature of
B. licheniformis 24 is the production of glycerol as the main by-product of glucose fermentation. This finding distinguishes
B. licheniformis 24 from most of the 2,3-BD-producing bacilli. Importantly, due to glycerol formation, the acidity of the medium changes slightly during the process and the fermentation could be performed without any pH control. Unlike other strains of the species,
B. licheniformis 24 does not produce formic acid at all. The accumulation of the toxic formic acid [
33] remains a serious problem against the industrial application of
B. licheniformis, as its amount could reach between 29.1 g/L and 42 g/L at the end of the process [
16,
19].
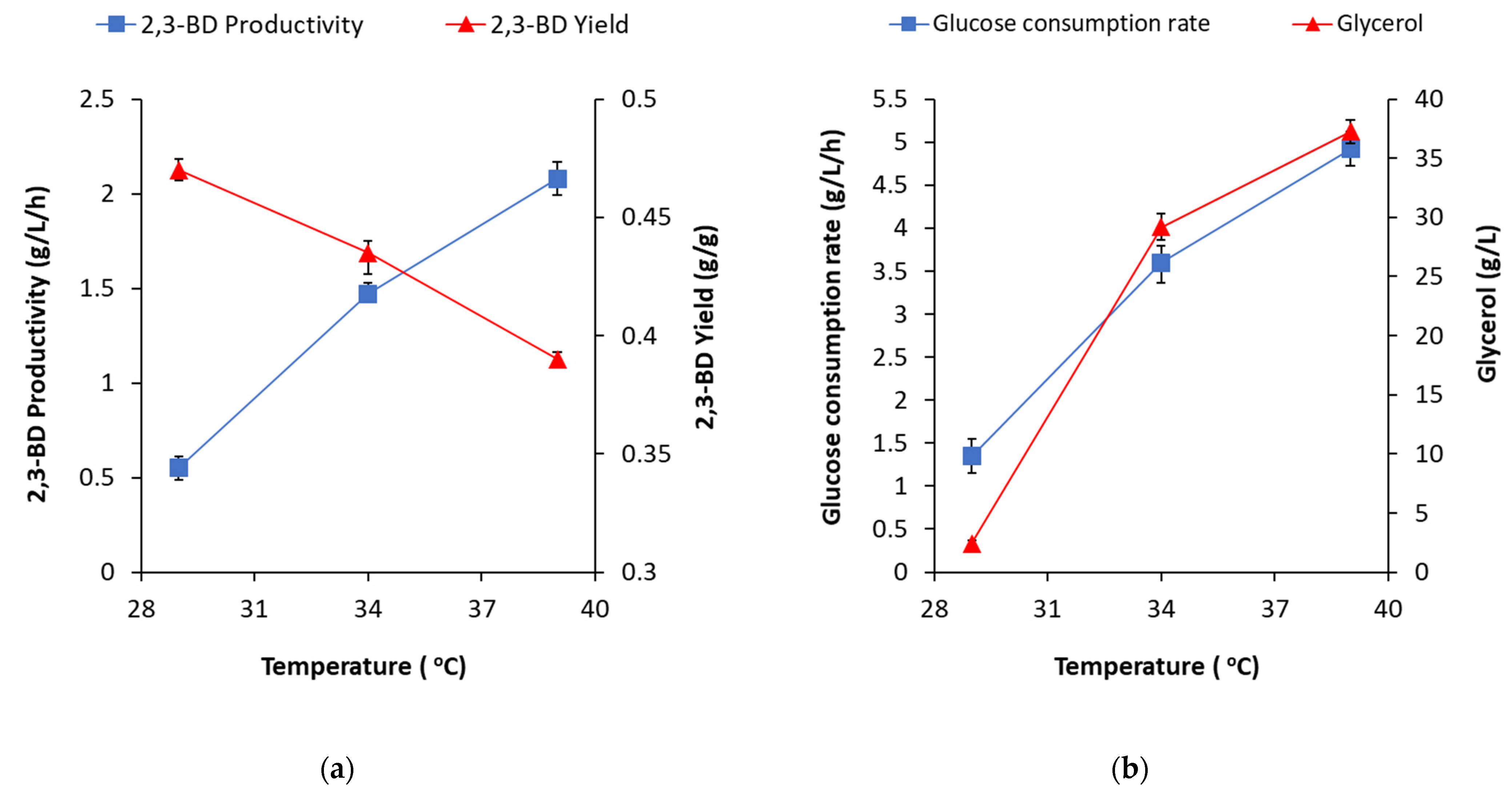
Therefore, an important achievement of the present study is the clarification of the process parameters that are suitable for maximum production of 2,3-BD by a strain, producing glycerol as a main by-product. To obtain the highest 2,3-BD concentration, according to maximized regression equation (4), the following process parameters were determined: temperature, 37.82 °C; pH, 6.23; airflow rate, 3.68 vvm. The temperature was found as the parameter with the highest impact, but the estimated optimal values were quite different for 2,3-BD concentration, 2,3-BD productivity and 2,3-BD yield. Indeed, the higher temperatures accelerate glucose consumption, which results in higher 2,3-BD productivity. For example, at a constant pH of 6.0 and aeration rate of 2 vvm, an increase in temperature from 29 °C to 39 °C raised 2,3-BD productivity from 0.55 to 2.08 g/L/h. However, due to higher glycerol formation, 2,3-BD yield decreased from 0.47 to 0.39 g/g (
Figure 4a). With a glucose consumption rate increase (from 1.35 to 4.92 g/L/h), glycerol accumulation rose from 2.42 to 37.25 g/L (
Figure 4b); therefore, at higher temperatures and low aeration, glycerol production appears to be favored.
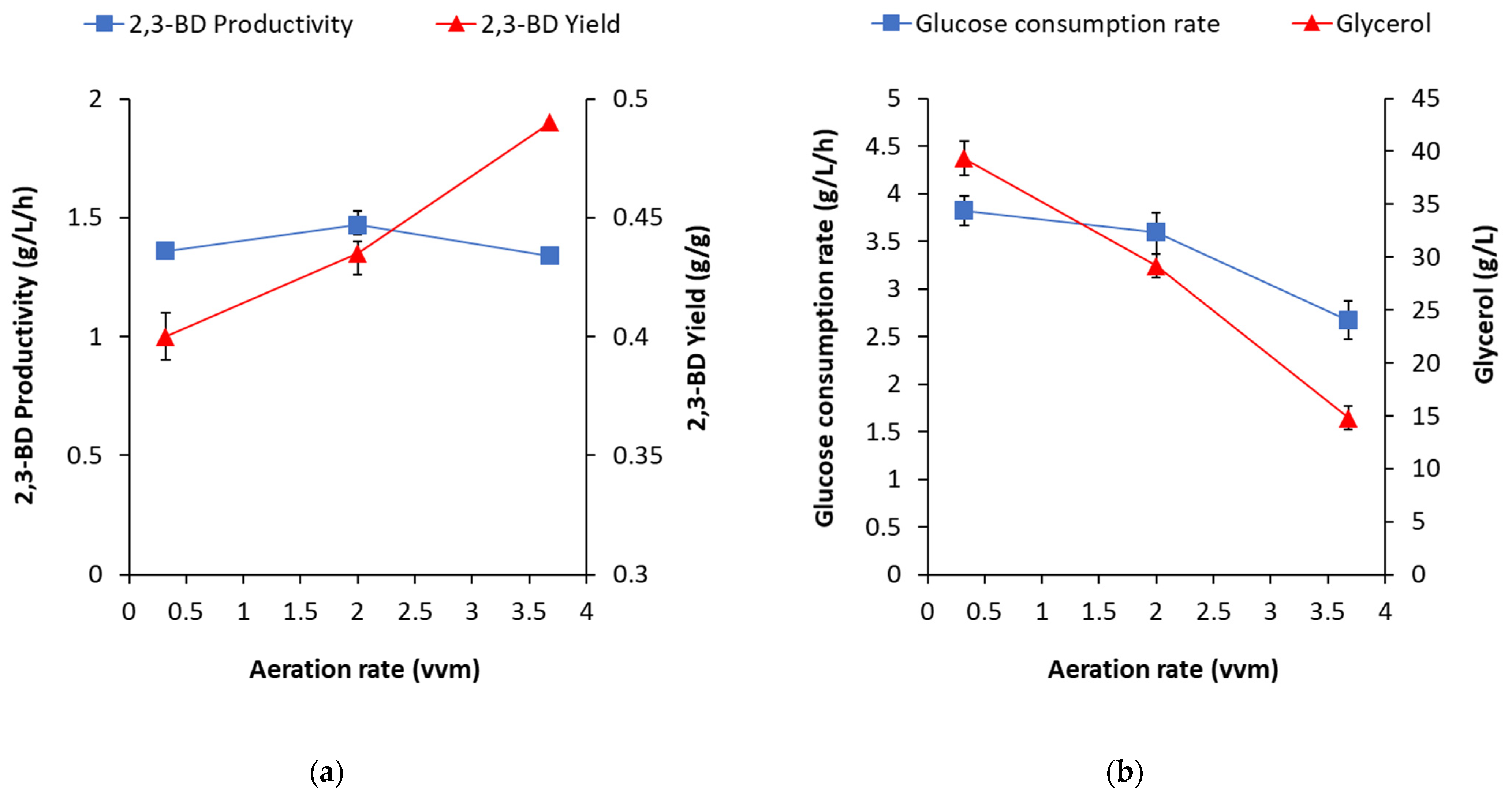
Conversely, the higher aeration levels favored 2,3-BD concentration increase and 2,3-BD yield and did not possess any significant effect on 2,3-BD productivity (
Figure 5a). Regarding glucose consumption and glycerol accumulation, the increase of aeration definitely affected the process in the opposite way with respect to the temperature rise (
Figure 5b).
According to Li et al. [
19] and Rebecchi et al. [
34], acetoin synthesis is strongly influenced by oxygen availability. Under fully aerobic conditions, the oxygen is the electron acceptor for NAD+ regeneration and acetoin reduction to 2,3-BD does not occur. The observation that acetoin production is greater at highly aerobic conditions determined the development of complex two-stage aeration regimes in the processes for obtaining 2,3-BD by
B. licheniformis [
16,
19,
34,
35]. On the contrary, when the oxygen supply is insufficient, the appropriate regeneration of co-factors such as NAD+ cannot be achieved by electron transfer onto oxygen, but, alternatively, NAD+ regeneration could be achieved by the reduction of dihydroxyacetone phosphate to glycerol-3-phosphate, which is then dephosphorylated yielding glycerol.
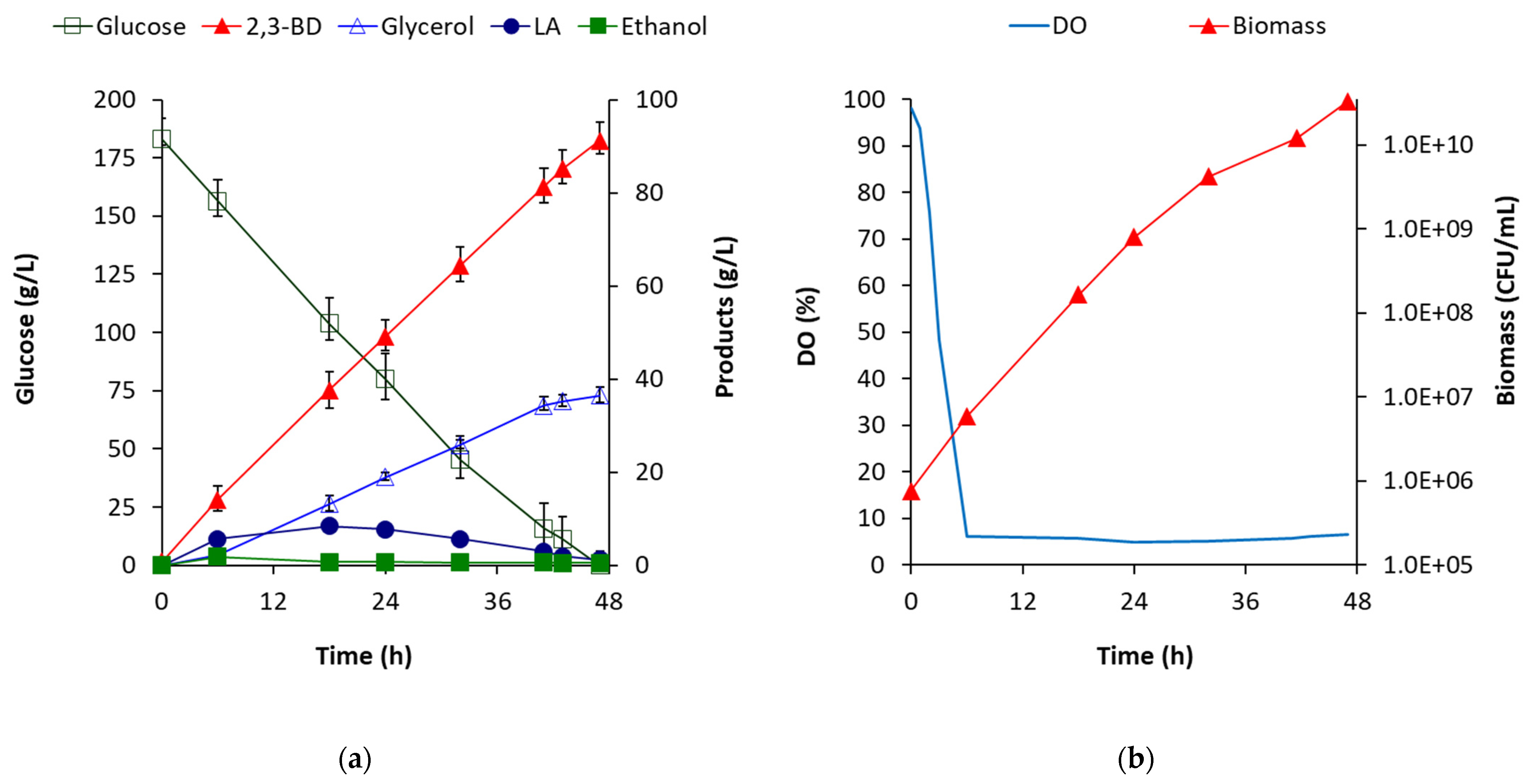
It turns out that
B. licheniformis strain 24 is an extremely aerobic 2,3-BD producer and is capable to consume huge oxygen amounts. As it was shown in
Figure 2b, the retention of dissolved oxygen in the medium remained constantly low, after the 6th hour, till the end of fermentation. This high oxygen consumption prevented acetoin formation even at the highest tested aeration levels. However, glycerol synthesis cannot be avoided even at high aeration levels.
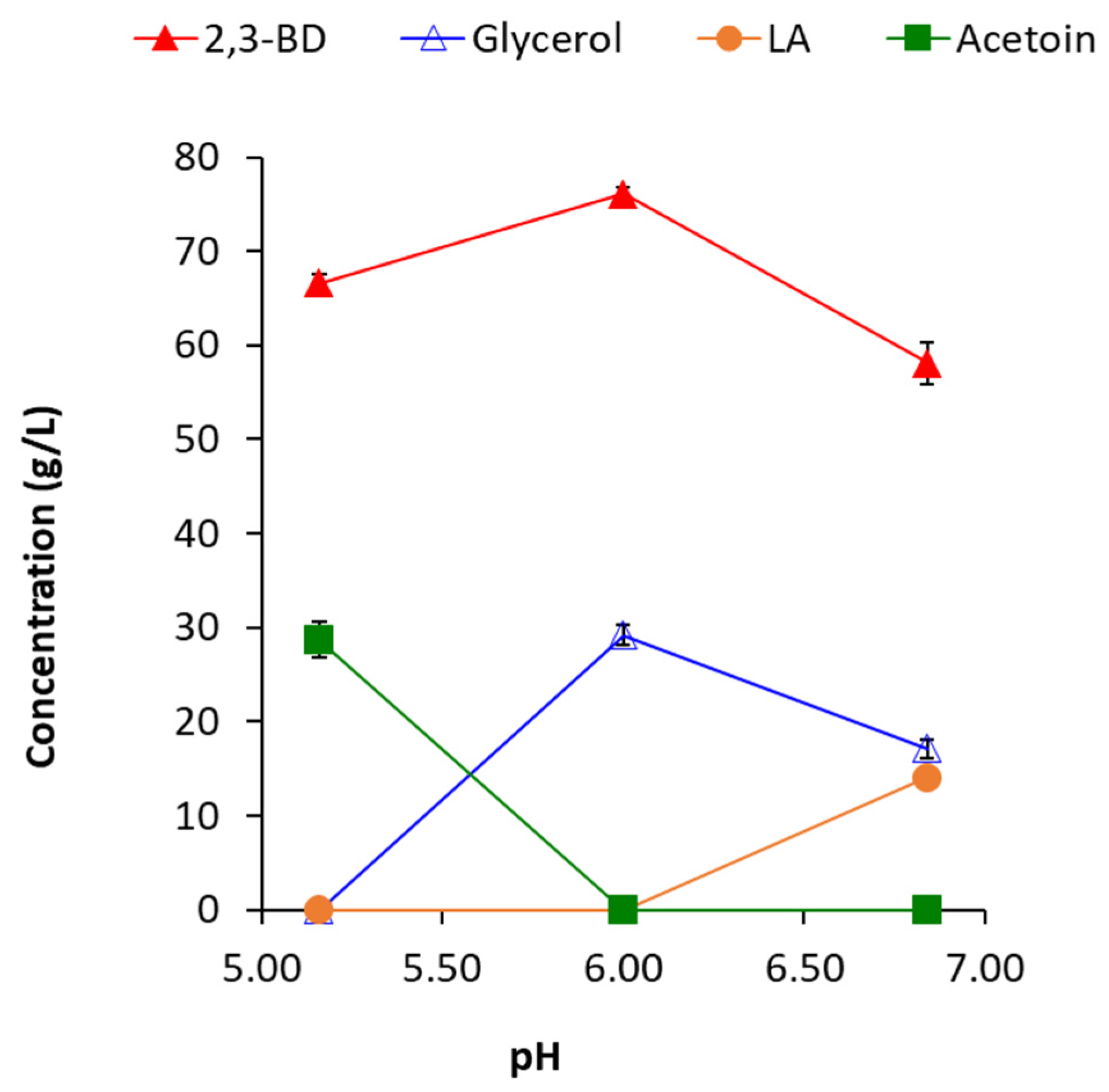
pH changes affect 2,3-BD production to a lesser extent. At lower pH (5.16), acetoin appeared as the main by-product, reaching concentrations of 28.64 g/L. Acetoin is accumulated at a low glucose consumption rate, in this case, at conditions combining lower pH (≤5.5) and lower temperature (≤31 °C). This positive effect of the high consumption rates of sugars (glucose, cellobiose and mannose) on 2,3-BD production at the expense of acetoin was always observed in
B. licheniformis 24 [
18].
The other significant by-product, glycerol, was not formed at lower pH (5.16), but increased at a pH of 6.0. However, Raspoet et al. [
27] showed that the effect of pH on glycerol produced is strain specific. Expectedly, lactic acid is accumulated in the highest amounts, 14.11 g/L, in the process with a pH of 6.84 (
Figure 6).
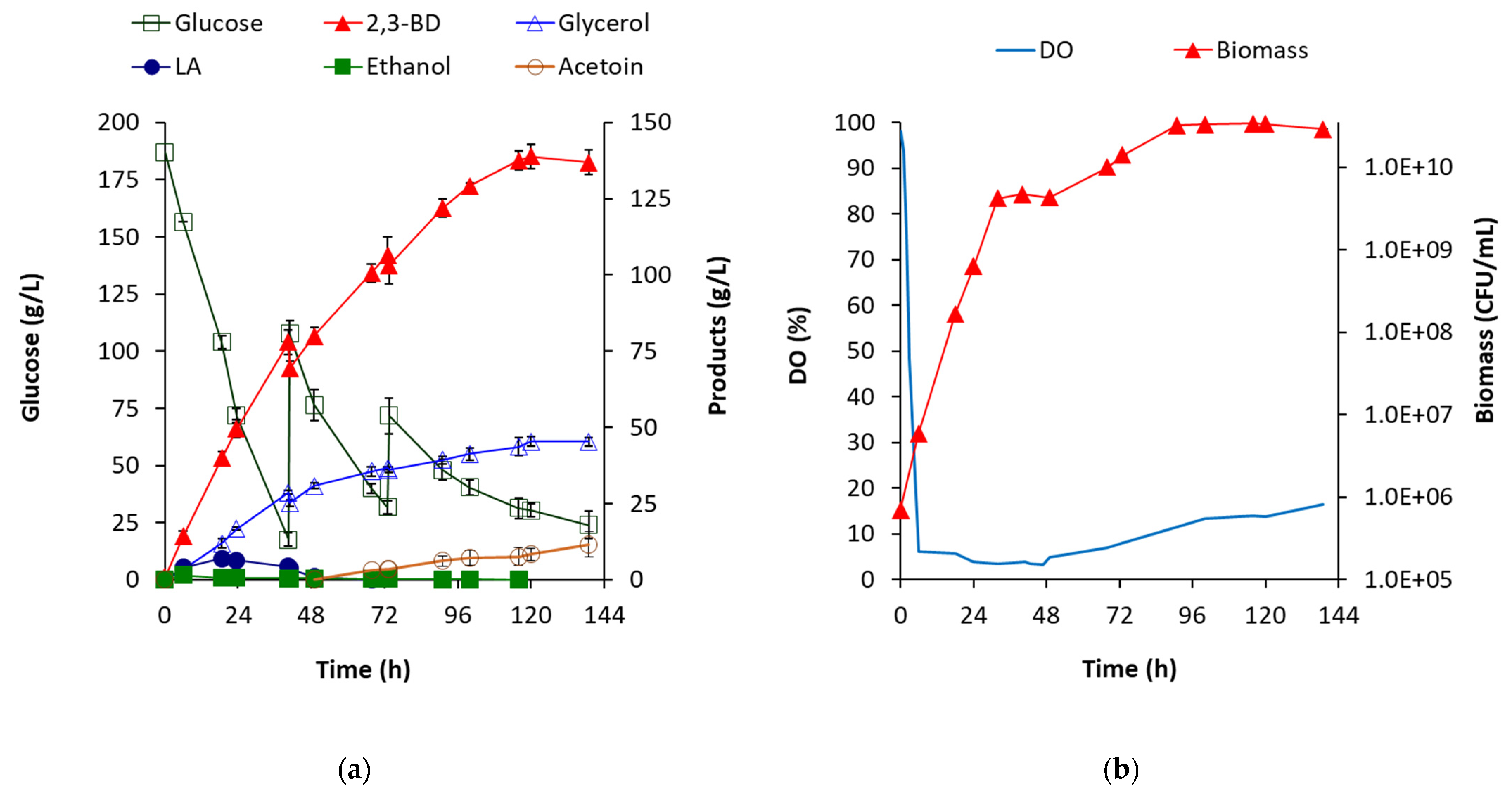
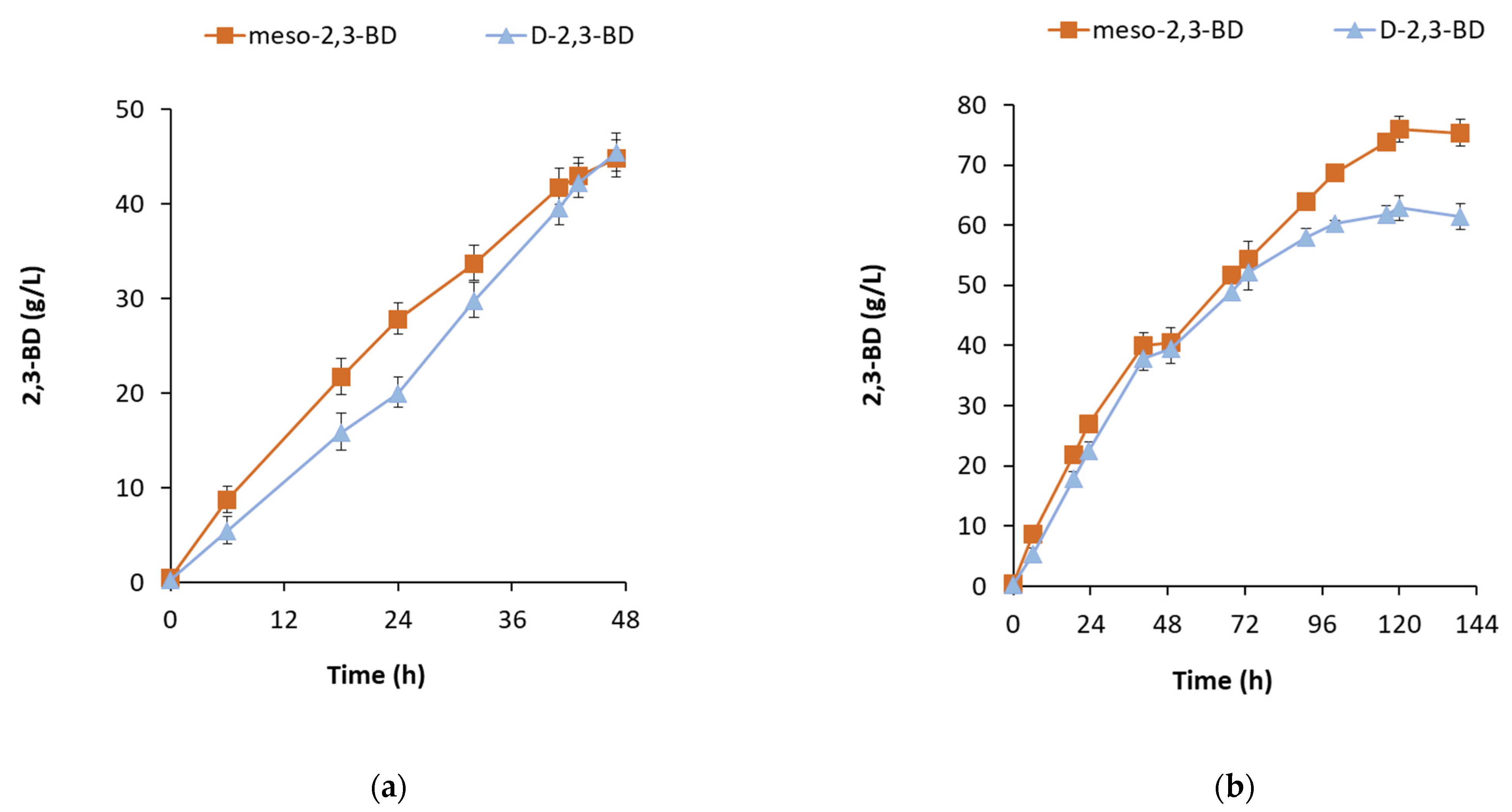
B. licheniformis 24 produces 2,3-BD in two isomeric forms, meso-2,3-BD and D-2,3-BD, in a ratio of 1.6:1–1:1. It has not been observed that the ratio depends on the substrate used, but it seems to depend on the total concentration of 2,3-BD produced. For example, at the beginning of each fermentation, the meso-form slightly prevailed, followed by equalization of the ratio at a total concentration of 80–90 g/L of 2,3-BD (
Figure 7a). This occurrence was observed in all batch processes. However, in the fed-batch process with optimized parameters, after the 70th h, the ratio changed again in favor of the meso-form and, by the end of the process, from 1.05:1, it became 1.23:1 (
Figure 7b). A possible explanation is the production of acetoin during this period (
Figure 3a), which was not observed in any of the batch processes when the carbon source was available. On the other hand, upon the complete depletion of the substrate, under the action of butanediol dehydrogenase BDH, the D-form was rapidly converted to acetoin, while the meso-form decreased slowly and was converted to acetoin only partially.
The effect of the presented complex optimization of medium composition and process parameters is the most obvious when the batch processes are compared before and after optimizations. The application of optimized parameters increased the maximal concentration of 2,3-BD by 28.9%, from 70.8 g/L, obtained without optimization [
18], to 91.23 g/L. The yield increased by 6.8%, from 0.457 g/g to 0.488 g/g substrate. Notably, the productivity increased more than 5-fold, from 0.38 g/L/h to 1.94 g/L/h.
After performing all the steps of optimization and validation, we conducted a fed-batch process, which aimed to reveal the full potential, or the upper limit, of the strain as a producer of 2,3-BD. The obtained 2,3-BD concentration of 138.8 ± 4.3 g/L, the productivity of 1.16 g/L/h and the yield, close to the theoretical (0.479 g/g), ranks
B. licheniformis 24 among the best non-pathogenic producers. Our results are very similar to the results presented by Jurchescu et al. [
20] and obtained by the use of
B. licheniformis DSM 8785: 2,3-BD concentration of 144.7 g/L and productivity of 1.14 g/L/h. A comparison of these results reveals that these values are probably the highest that can be obtained using the species
B. licheniformis and comprise its maximum capabilities as a 2,3-BD producer. Compared to the best pathogenic producers as
Klebsiella pneumoniae or
K. oxytoca, it is obvious that they have comparable tolerance to 2,3-BD and produce relatively equal quantities of 2,3-BD, reaching 150 g/L. However,
Klebsiella strains achieve this concentration up to four times faster [
36,
37,
38]. Therefore, the low productivity continues to be an obstacle for industrial application of even the most successful non-pathogenic 2,3-BD producers of the species
B. licheniformis.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}