
Differentiation of Penicillium roqueforti from Closely Related Species Contaminating Cheeses and Dairy Environment


Abstract
1. Introduction
2. Materials and Methods
2.1. Strain Cultivation and Morphological Features
2.2. P. roqueforti-Specific Primer Design
2.2.1. DNA Extraction from Mycelial Samples
2.2.2. ITS and β-Tubulin Sequencing
2.3. ITS-Pr Primer Testing
3. Results
3.1. Morphological Features
3.1.1. Macromorphology
3.1.2. Micromorphology
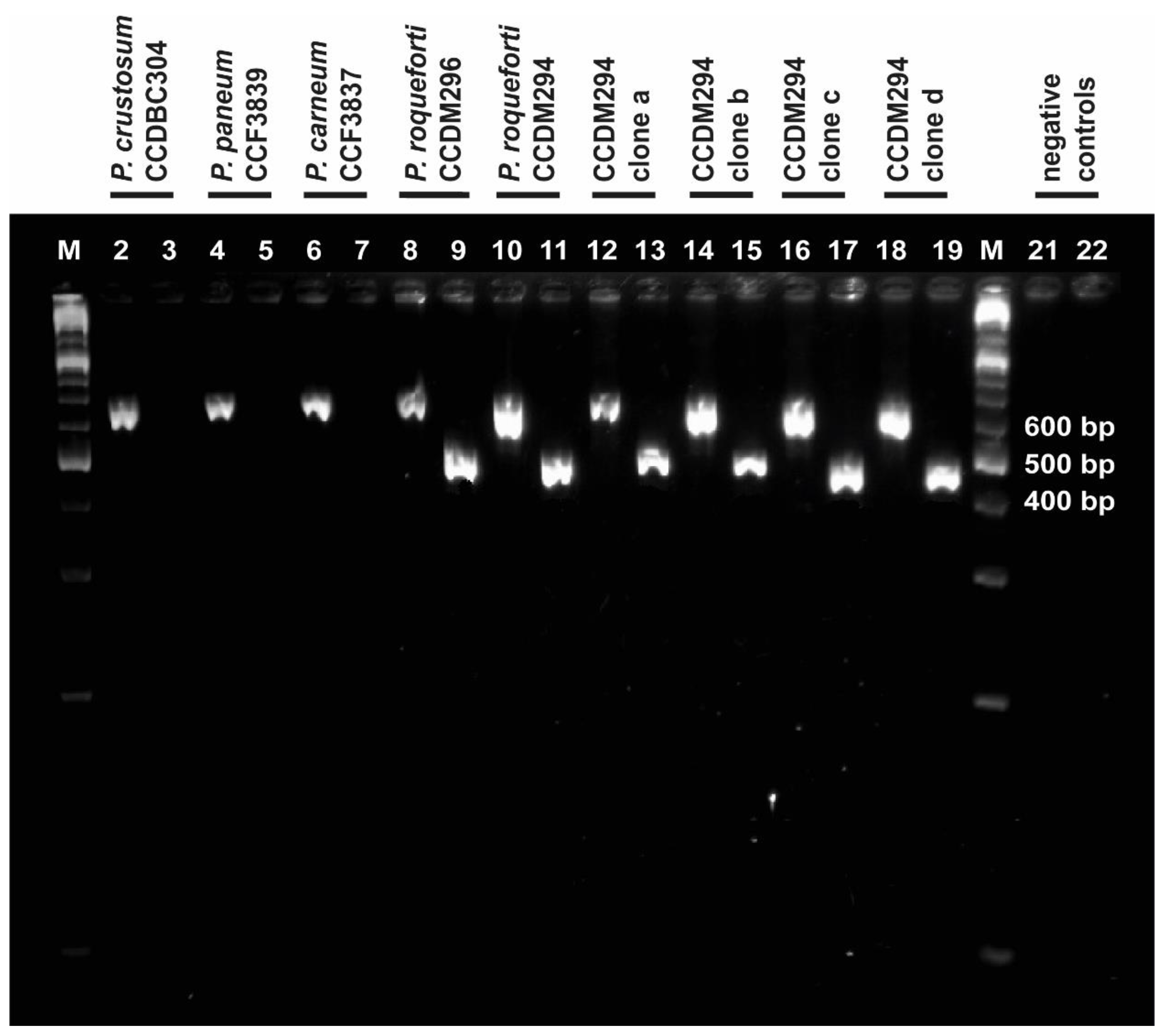
3.2. Verification of the Taxonomic Affiliation of Target Species and Strains via PCR
3.3. Application of ITS-Pr Primer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Strain/Clone | Diameter (mm) * | Figure Position | Colony Texture | Margin | Colour | |
|---|---|---|---|---|---|---|
| Line 1 | Munsell | |||||
| CCDBC 304 | 26.5 d ± 1.22 | A | Velutinous, irregular, flat | Arachnoid, creamy white, regular | Pale green, yellow green in the centre | 5G 7/2 |
| CCF 3839 | 25.6 d± 0.88 | B | Velutinous colony, rugose, partially wrinkled (2/3), irregular | Arachnoid, undulate | Pale blue | 5B 6/2 |
| CCF 3837 | 25.8 d± 1.15 | C | Velutinous colony, irregular, partially fasciculate in centre | Arachnoid, undulate | Pale blue | 5B 6/2 |
| CCDM 296 | 36.4 ab ± 0.46 | D | Velutinous colony | Arachnoid, creamy white, regular | Pale blue green | 5BG 7/2 |
| CCDM 294 | 38.3 a ± 0.72 | E | Velutinous partially fasciculate in centre, flat, | Arachnoid, creamy white, irregular | Pale blue green | 5BG 7/2 |
| Clone 294a | 42.1 a ± 1.15 | F | Velutinous flat | Arachnoid, creamy white, regular | Pale blue green | 5BG 7/2 |
| Clone 294b | 41.6 a ± 1.54 | G | Velutinous partially fasciculate, flat, | Arachnoid, creamy white, regular | Pale blue green | 5BG 7/2 |
| Clone 294c | 38.41 a± 0.81 | H | Velutinous partially fasciculate, flat, | Arachnoid, creamy white, regular | Pale blue green | 5BG 7/2 |
| Clone 294d | 27.8 d ± 0.32 | I | Velutinous, umbonate, irregular | Arachnoid, creamy white, irregular | Pale green | 10G 6/2 |
| Strain/Clone | Diameter * (mm) | Figure Position | Colony Texture | Margin | Colour | |||
|---|---|---|---|---|---|---|---|---|
| Line 2, 3 | Upper Side | Munsell | Reverse | Munsell | ||||
| CCDBC 304 | 36.98 e ± 0.38 | A | Velutinous, rugose, wrinkles from centre to edges, regular | Creamy white, regular | Greyish blue green | 5BC 5/2 | Yellow, slight wrinkles | 5Y 9/10 |
| CCF 3839 | 25.25 d ± 0.32 | B | Velutinous, fasciculate in the centre, exudate droplets, irregular | Irregular, white | Moderate blue | 5B 5/6 | Yellow, slight wrinkles | 5Y 9/14 |
| CCF 3837 | 36.70 e ± 0.34 | C | Velutinous, flat, fasciculate in the centre, wrinkled, exudates, irregular | Creamy white, regular | Pale blue green | 5BG 7/2 | Yellow with pale orange in centre, wrinkles, cracks | 7 YR 7/8 |
| CCDM 296 | 32.95 c ± 0.38 | D | Velutinous, partially filamentous, flat, irregular | White, irregular, arachnoid | Pale blue green | 5BG 7/2 | Dusky yellowish greenYellow margins | 10GY 3/2 |
| CCDM 294 | 31.72 c ± 0.32 | E | Velutinous, partially filamentous fasciculate in the centre | White, irregular, arachnoid | Pale blue green | 5BG 7/2 | Dusky yellowish greenYellow intercalary | 10GY 3/27.5Y 7/164 |
| Clone 294a | 34.76 ce ± 0.23 | F | Velvety, fasciculate in the centre | Creamy white, regular | Pale blue green | 5BG 7/2 | Dusky yellowish greenYellow intercalary | 10GY 3/27.5Y 7/164 |
| Clone 294b | 30.6 b ± 0.32 | G | Velutinous, partially filamentous, fasciculate, flat | White regular thin | Pale blue green | 5BG 7/2 | Moderate yellow | 5Y 7/6 |
| Clone 294c | 29.78 b ± 0.28 | H | Velutinous, partially filamentous, sparse, flat | White irregular thin | Pale blue green | 5BG 7/2 | Dusky yellowish greenYellow—only margins | 10GY 3/27.5Y, 7/164 |
| Strain/Clone | Diameter (mm) * | Figure Position | Colony Texture | Margin | Colour | |||
|---|---|---|---|---|---|---|---|---|
| Line 4, 5 | Upper Side | Munsell | Reverse | Munsell | ||||
| CCDBC 304 | 37.2 d± 0.8 | A | Velutinous, rugose, regular radial wrinkles | White, regular | Greyish green | 10GY 5/2 | Yellow, radial wrinkles | 7.5Y 9/10 |
| CCF 3839 | 29.7 e ± 0.8 | B | Velutinous, rugose, irregular wrinkles and cracks | White, regular | Pale blue | 5B 6/2 | Yellow, radial wrinkles, cracks | 5Y 8/16 |
| CCF 3837 | 37.8 d ± 0.8 | C | Velutinous, rugose, weaved wrinkles radial, dark exudates in the centre | White, regular | Pale blue green | 5BG 7/2 | Yellow, radial rippled wrinkles, orange centre | 5Y 8/16 7.5R 8/10 |
| CCDM 296 | 20.4 a ± 0.8 | D | Velutinous, rugose, weaved wrinkles radial and concentric | White, irregular, arachnoid | Pale blue green | 5BG 7/2 | Dusky yellowish green Yellow margin | 10GY 3/2 7.5Y 7/164 |
| CCDM 294 | 25.4 b± 0.8 | E | Velutinous, rugose, weaved wrinkles radial and concentric | White, irregular, arachnoid | Pale blue green | 5BG 7/2 | Yellow, radial wrinkles, cracks | 7.5Y 9/6 |
| Clone 294a | 32.3 ± 0.8 | F | Velutinous, rugose, weaved wrinkles radial | White, irregular, arachnoid | Pale blue green | 5BG 7/2 | Dusky yellowish green Yellow, radial wrinkles Mycelia intercalary | 7.5Y 9/8 |
| Clone 294b | 32.1 c ± 0.8 | G | Velutinous, rugose, weaved wrinkles radial | White, irregular, arachnoid | Pale blue green | 5BG 7/2 | Yellow, radial wrinkles, cracks, intergrow white mycelia | 7.5Y 9/6 |
| Clone 294c | 26.7 b ± 0.8 | H | Velutinous, rugose, weaved wrinkles radial, irregular, short | White, irregular, arachnoid | Pale blue green | 5BG 7/2 | Dusky yellowish green, yellow margin cracks radially | 10GY 3/2 7.5Y 7/164 |
| Clone 294d | 19.4 a ± 0.8 | I | Velutinous, rugose, weaved wrinkles, short cranks, concentric, irregular, short | White, irregular, arachnoid | Pale blue | 5B 6/2 | Dusky yellowish green margin, yellow weaved cracks irregular | 10GY 3/2 7.5Y 7/164 |
| Strain/Clone | Diameter (mm) * | Figure Position | Colony Characteristics | Margin | Colour | |
|---|---|---|---|---|---|---|
| Line 6 | Munsell | |||||
| CCDBC 304 | 22.4 b ± 0.6 | A | Velvety, circular form, small droplets, flat intensive discoloration of CREA | Irregular arachnoid white | Very pale green | 10G 5/2 |
| CCF 3839 | 13.4 a± 0.8 | B | Velvety, circular form, fasciculate in the centre, discoloration in the centre only | Irregular arachnoid white | Light blue green | 5BG 5/2 |
| CCF 3837 | 23.1 b ± 0.3 | C | Velvety, circular form, droplets in the centre, flat | Irregular arachnoid white | Light green to light blue green | 5G 7/4 5BG 5/2 |
| CCDM 296 | 17.4 a ± 1.2 | D | Velvety, circular form, fasciculate in the centre, flat | Irregular arachnoid white | Very pale green | 10G 5/2 |
| CCDM 294 | 21.6 b ± 0.5 | E | Velvety, circular form, fasciculate in the centre, flat | Irregular arachnoid white | Pale blue | 5B 6/2 |
| Clone 294a | 23.8 b ± 0.4 | F | Velvety, circular form, fasciculate in the centre, flat | Irregular arachnoid white | Light blue green | 5BG 6/6 |
| Clone 294b | 20.2 ab± 0.6 | G | Velvety, circular form, fasciculate in the centre, umbonate | Irregular arachnoid white | Pale green | 10G 6/2 |
| Clone 294c | 22.4 b ± 1.2 | H | Velvety, irregular form, fasciculate in the centre, flat | Irregular arachnoid white | Moderate yellowish green | 10GY 6/4 |
| Clone 294d | 17.5 a ± 0.2 | I | Velvety, circular form, fasciculate in the centre, flat | Irregular arachnoid white | Very pale green | 10G 5/2 |
| Strain/Clone | Diameter (mm) * | Figure Position | Colony Characteristics | Margin | Colour | |
|---|---|---|---|---|---|---|
| Line 8 | Munsell | |||||
| CCDBC 304 | 21.8 b ± 0.9 | A | Velutinous, regular form, raised | Irregular, blue green | Moderate blue green | 5BG 4/6 |
| CCF 3839 | 24.5 c± 0.3 | B | Velutinous, irregular form, wavy pattern | Irregular, blue green | Dusky blue green | 5BG 3/2 |
| CCF 3837 | 24.4 c± 0.5 | C | Velutinous, irregular form, rugose, flat | Irregular, green, indistinct | Dusky yellowish green | 10GY 3/2 |
| CCDM 296 | 22.5 bc ± 0.8 | D | Velutinous, irregular form, rugose, aerial mycelia | Irregular, blue-green, indistinct | Pale blue green | 5BG 7/2 |
| CCDM 294 | 20.8 b ± 0.8 | E | Velutinous, irregular form, rugose, secondary white mycelia | Irregular, greyish green, indistinct | Greyish green | 10G 4/2 |
| Clone 294a | 21.6 b ± 1.4 | F | Velutinous, irregular form, flat, exudate droplets | Irregular, green | Greyish green | 10G 4/2 |
| Clone 294b | 24.7 c ± 1.2 | G | Velutinous, irregular form, rugose, secondary aerial mycelia | Irregular, white-blue shaggy | Dusty blue green | 5 BG 3/2 |
| Clone 294c | 23.6 c± 0.5 | H | Velutinous, irregular form, umbonate | Irregular, arachnoid | Greyish green | 10G 4/2 |
| Clone 294d | 14.3 a ± 1.2 | I | Velutinous, irregular form, rugose | Irregular, white thin | Greyish blue | 5 PB 5/2′ |
| Strain/Clone | Diameter (mm) * | Figure Position | Colony Characteristics | Margin | Colour | |
|---|---|---|---|---|---|---|
| Line 7 | Munsell | |||||
| CCDBC 304 | 26.49 c ± 0.21 | A | Fasciculate, regular flat form | White regular | Moderate yellowish green | 10GY6/4 |
| CCF 3839 | 24.51 a ± 0.34 | B | Velutinous, regular flat form | White regular | Moderate blue green | 5BG 4/6 |
| CCF 3837 | 24.24 d ± 0.52 | C | Velutinous, powdery, regular flat form | White regular | Pale blue green | 5BG 7/2 |
| CCDM 296 | 25.21 a ± 0.21 | D | Powdery, irregular flat form | Yellow, arachnoid, irregular | Dark yellowish green | 10 GY 4/4 |
| CCDM 294 | 26.03 a ± 0.32 | E | Powdery, regular flat form | White irregular, 1/3 of whole colony diameter | Greyish green | 10G 5/2 |
| Clone 294a | 28.5 c ± 0.32 | F | Powdery, regular flat form | Creamy, arachnoid, irregular | Greyish blue green | 10G 5/2 |
| Clone 294b | 24.09 b ± 0.32 | G | Powdery, regular flat form | Regular, white, ½ of whole colony diameter | Dark yellowish green | 10GY 4/4 |
| Clone 294c | 24.67 a ± 0.32 | H | Powdery, irregular flat form | White irregular, 1/3 of whole colony diameter | Dark yellowish green | 10GY 4/4 |
| Clone 294d | 25.45 a ± 0.32 | I | Powdery, regular flat form, fasciculate in the centre | White regular | Yellowish gray | 5Y 8/1 |
| Strains | Accession No. |
|---|---|
| CCDM 296 Penicillium roqueforti | MZ520338 |
| CCDM 294 +clones a, b, c, d Penicillium roqueforti | MZ520339, MZ520340, MZ520341, MZ520342, MZ520343 |
| CCF 3837 Penicillium carneum | MZ520344 |
| CCF 3839 Penicillium paneum | MZ520345 |
| CCDBC 304 Penicillium crustosum | MZ520346 |
| CCDBC 331 Penicillium solitum | MZ520347 |
| CCDBC 311 Penicillium chrysogenum | MZ520348 |
| CCDBC 303 Penicillium rubens | MZ520349 |
| CCDBC 312 Penicillium glabrum | MZ520350 |
| CCDBC 307 Penicillium expansum | MZ520351 |
References
- Dumas, E.; Feurtey, A.; de la Vega, R.C.R.; Le Prieur, S.; Snirc, A.; Coton, M.; Thierry, A.; Coton, E.; Le Piver, M.; Roueyre, D.; et al. Independent domestication events in the blue-cheese fungus Penicillium roqueforti. Mol. Ecol. 2020, 29, 2639–2660. [Google Scholar] [CrossRef] [PubMed]
- Boysen, M.; Skouboe, P.; Frisvad, J.; Rossen, L. Reclassification of the Penicillium Roqueforti Group into Three Species on the Basis of Molecular Genetic and Biochemical Profiles. Microbiology 1996, 142, 541–549. [Google Scholar] [CrossRef]
- Boysen, M.E.; Jacobsson, K.-G.; Schnürer, J. Molecular Identification of Species from the Penicillium roqueforti Group Associated with Spoiled Animal Feed. Appl. Environ. Microbiol. 2000, 66, 1523–1526. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Sex in Penicillium series Roqueforti. IMA Fungus 2010, 1, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Suarez, M.; García, D.; Cano-Lira, J.; Guarro, J.; Gené, J. Species diversity in Penicillium and Talaromyces from herbivore dung, and the proposal of two new genera of penicillium-like fungi in Aspergillaceae. Fungal Syst. Evol. 2020, 5, 39–76. [Google Scholar] [CrossRef]
- O’Brien, M.; Egan, D.; O’Kiely, P.; Forristal, P.D.; Doohan, F.M.; Fuller, H.T. Morphological and molecular characterisation of Penicillium roqueforti and P. paneum isolated from baled grass silage. Mycol. Res. 2008, 112, 921–932. [Google Scholar] [CrossRef]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi, 1st ed.; Crous, P.W., Samson, R.A., Eds.; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010; p. 390. [Google Scholar]
- Gillot, G.; Jany, J.-L.; Coton, M.; Le Floch, G.; Debaets, S.; Ropars, J.; López-Villavicencio, M.; Dupont, J.; Branca, A.; Giraud, T.; et al. Insights into Penicillium roqueforti Morphological and Genetic Diversity. PLoS ONE 2015, 10, e0129849. [Google Scholar] [CrossRef] [PubMed]
- Cantor, M.; Tempel, T.V.D.; Hansen, T.; Ardö, Y. Blue cheese. In Cheese: Chemistry, Physics and Microbiology; Academic Press: Cambridge, MA, USA, 2004; Volume 2, pp. 175–198. [Google Scholar]
- Larsen, M.D.; Jensen, K. The effects of environmental conditions on the lipolytic activity of strains of Penicillium roqueforti. Int. J. Food Microbiol. 1999, 46, 159–166. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Kiss, L. Limits of nuclear ribosomal DNA internal transcribed spacer (ITS) sequences as species barcodes for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, E1811. [Google Scholar] [CrossRef] [PubMed]
- Seifert, K.A.; Samson, R.A.; Dewaard, J.R.; Houbraken, J.; Lévesque, C.A.; Moncalvo, J.-M.; Louis-Seize, G.; Hebert, P. Prospects for fungus identification using CO1 DNA barcodes, with Penicillium as a test case. Proc. Natl. Acad. Sci. USA 2007, 104, 3901–3906. [Google Scholar] [CrossRef] [PubMed]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.; Hong, S.-B.; Klaassen, C.; Perrone, G.; Seifert, K.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.; Nielsen, P. Development of a Semisynthetic Cheese Medium for Fungi Using Chemometric Methods. J. Dairy Sci. 1997, 80, 1237–1245. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2009, 27, 221–224. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 315–322. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Mounier, J. Diversity and Control of Spoilage Fungi in Dairy Products: An Update. Microorganisms 2017, 5, 42. [Google Scholar] [CrossRef]
- Ropars, J.; López-Villavicencio, M.; Snirc, A.; Lacoste, S.; Giraud, T. Blue cheese-making has shaped the population genetic structure of the mould Penicillium roqueforti. PLoS ONE 2017, 12, e0171387. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553. [Google Scholar] [CrossRef]
- Wang, X.-C.; Chen, K.; Zeng, Z.-Q.; Zhuang, W.-Y. Phylogeny and morphological analyses of Penicillium section Sclerotiora (Fungi) lead to the discovery of five new species. Sci. Rep. 2017, 7, 8233. [Google Scholar] [CrossRef][Green Version]
- Chen, K.; Tian, Z.; Jiang, F.; Long, C.-A. Development of Penicillium italicum-Specific Primers for Rapid Detection among Fungal Isolates in Citrus. J. Microbiol. Biotechnol. 2019, 29, 984–988. [Google Scholar] [CrossRef]
- Vanittanakom, N.; Vanittanakom, P.; Hay, R.J. Rapid Identification of Penicillium marneffei by PCR-Based Detection of Specific Sequences on the rRNA Gene. J. Clin. Microbiol. 2002, 40, 1739–1742. [Google Scholar] [CrossRef]
- Zhu, P.; Wu, L.; Liu, L.; Huang, L.; Wang, Y.; Tang, W.; Xu, L. Fusarium asiaticum: An Emerging Pathogen Jeopardizing Postharvest Asparagus Spears. J. Phytopathol. 2013, 161, 696–703. [Google Scholar] [CrossRef]
- Rychlik, W. Selection of primers for polymerase chain reaction. Mol. Biotechnol. 1995, 3, 129–134. [Google Scholar] [CrossRef]
- Simsek, M.; Adnan, H. Effect of single mismatches at 3’-end of primers on polymerase chain reaction. J. Sci. Res. Med. Sci. 2000, 2, 11–14. [Google Scholar]
- Bru, D.; Martin-Laurent, F.; Philippot, L. Quantification of the Detrimental Effect of a Single Primer-Template Mismatch by Real-Time PCR Using the 16S rRNA Gene as an Example. Appl. Environ. Microbiol. 2008, 74, 1660–1663. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Yang, X.; Zheng, F.; Long, C.-A. Genome sequencing and analysis of Kloeckera apiculata strain 34-9, a biocontrol agent against postharvest pathogens in citrus. Genes Genom. 2016, 39, 87–99. [Google Scholar] [CrossRef]



| Strains | Origin | Accession No. |
|---|---|---|
| CCDM 296 Penicillium roqueforti | Culture Collection of Dairy Microorganisms, Milcom Ltd., Czech Republic | MW600465 |
| CCDM 294 +clones a, b, c, d Penicillium roqueforti | Culture Collection of Dairy Microorganisms, Milcom Ltd., Czech Republic | MW600466, MW600467, MW600468, MW600469, MW600470 |
| CCF 3837 Penicillium carneum | Culture Collection of Fungi, Dpt. Botany, Fac. Biology, Charles University, Prague, Czech Republic | MW600471 |
| CCF 3839 Penicillium paneum | Culture Collection of Fungi, Dpt. Botany, Fac. Biology, Charles University, Prague, Czech Republic | MW600472 |
| CCDBC 304 Penicillium crustosum | Culture Collection of Dairy and Bakery Contaminants, Milcom Ltd., Czech Republic | MW600473 |
| CCDBC 331 Penicillium solitum | Culture Collection of Dairy and Bakery Contaminants, Milcom Ltd., Czech Republic | MW600474 |
| CCDBC 311 Penicillium chrysogenum | Culture Collection of Dairy and Bakery Contaminants, Milcom Ltd., Czech Republic | MW600475 |
| CCDBC 303 Penicillium rubens | Culture Collection of Dairy and Bakery Contaminants, Milcom Ltd., Czech Republic | MW600476 |
| CCDBC 312 Penicillium glabrum | Culture Collection of Dairy and Bakery Contaminants, Milcom Ltd., Czech Republic | MW600477 |
| CCDBC 307 Penicillium expansum | Culture Collection of Dairy and Bakery Contaminants, Milcom Ltd., Czech Republic | MW600478 |
| Medium Acronym | Medium Description | Origin | Cultivation | References |
|---|---|---|---|---|
| MEA | Malt Extract Agar | MERCK KGaA, Germany | 25 °C | Samson et al., 2010 [7] |
| CREA | Creatine Sucrose Agar | MILCOM a.s.; Czech Republic | 25 °C | |
| YES | Yeast Extract Sucrose Agar | MILCOM a.s.; Czech Republic | 25 °C | |
| CYA | Czapek Yeast extract Agar | MILCOM a.s.; Czech Republic | 30 °C | |
| Cheese-like media | ||||
| Edam-like | A total of 278 g grated Eidam cheese after salting, 630 g Cream 32% fat, 91.3 g Reconstituted skim milk,10 g NaCl, 10 g Trisodium citrate dihydrate, 10 g Tripotassium citrate monohydrate, 12 g agar, 250 mL UHT milk 3.5% fat, homogenization at 85 °C for 10 min in Thermomix (Vorwerk CS, Prague, Czech Republic) | 25 °C | Modified Hansen and Nielsen, 1996 [16] | |
| Roquefort-like | A total of 200 g grated Roquefort cheese after salting, 30 g NaCl, 10 g agar, tap water to 1000 mL, homogenization by Ultra-Turrax T25 at 9500 min−1 (Janke-Kunkel GmbH, Staufen, Germany), sterilization 102 °C for 20 min | 25 °C | ||
| Strains | Section | Accession No. |
|---|---|---|
| Penicillium roqueforti | Roquefortorum | EU427296 1, MITS3851 2, MITS3852 2, MITS3853 2 |
| Penicillium carneum | Roquefortorum | HQ442338 1 |
| Penicillium paneum | Roquefortorum | HQ442346 1 |
| Penicillium psychrosexualis | Roquefortorum | HQ442345 1 |
| Penicillium mediterraneum | Roquefortorum | NR172393 1 |
| Penicillium crustosum | Fasciculata | AF033472 1 |
| Penicillium solitum | Fasciculata | AY373932 1 |
| Penicillium chrysogenum | Chrysogena | AF033465 1 |
| Penicillium rubens | Chrysogena | JX997057 1 |
| Penicillium glabrum | Aspergiloides | GU981567 1 |
| Penicillium expansum | Penicillium | AY373912 1 |
| Source database | 1 NCBI http://www.ncbi.nlm.nih.gov/ (accessed on 7 October 2021) 2 ISHAM http://its.mycologylab.org/ (accessed on 7 October 2021) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kavková, M.; Cihlář, J.; Dráb, V.; Bár, L. Differentiation of Penicillium roqueforti from Closely Related Species Contaminating Cheeses and Dairy Environment. Fermentation 2021, 7, 222. https://doi.org/10.3390/fermentation7040222
Kavková M, Cihlář J, Dráb V, Bár L. Differentiation of Penicillium roqueforti from Closely Related Species Contaminating Cheeses and Dairy Environment. Fermentation. 2021; 7(4):222. https://doi.org/10.3390/fermentation7040222
Chicago/Turabian StyleKavková, Miloslava, Jaromír Cihlář, Vladimír Dráb, and Ladislav Bár. 2021. "Differentiation of Penicillium roqueforti from Closely Related Species Contaminating Cheeses and Dairy Environment" Fermentation 7, no. 4: 222. https://doi.org/10.3390/fermentation7040222
APA StyleKavková, M., Cihlář, J., Dráb, V., & Bár, L. (2021). Differentiation of Penicillium roqueforti from Closely Related Species Contaminating Cheeses and Dairy Environment. Fermentation, 7(4), 222. https://doi.org/10.3390/fermentation7040222