
Influence of Human Age on the Prebiotic Effect of Pectin-Derived Oligosaccharides Obtained from Apple Pomace

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. High-Performance Anion-Exchange Chromatography
2.2.2. In Vitro Fecal Fermentation and Qualitative and Quantitative Assessment of the Microbiome
2.2.3. Koch’s Plating Method
2.2.4. Fluorescence In Situ Hybridization (FISH)
2.2.5. Short-Chain Fatty Acid (SCFA) Analysis
2.2.6. Lactic Acid Isomers Analysis
2.2.7. Statistical Analysis
3. Results
3.1. Quantitative and Qualitative Composition of POS
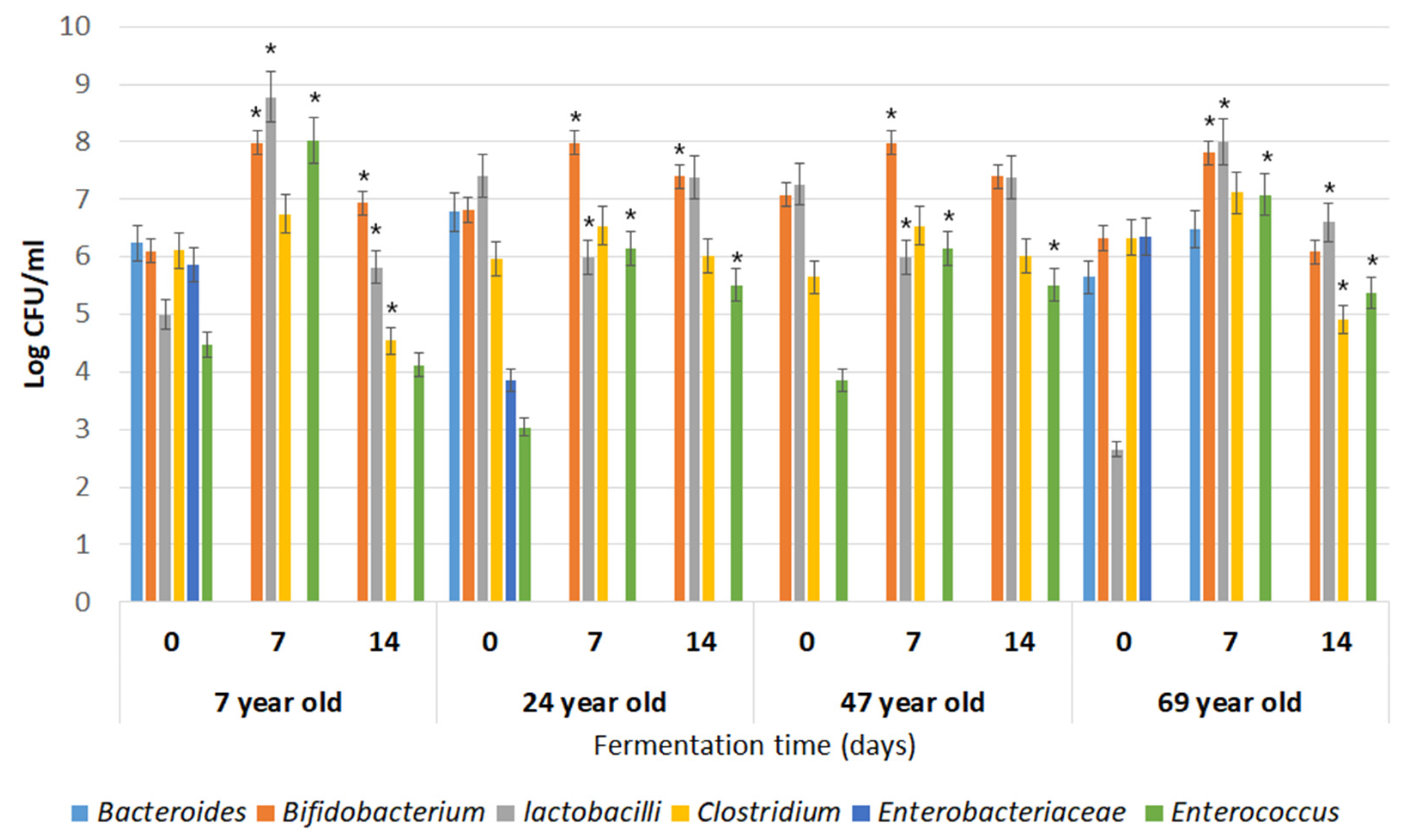
3.2. Growth Dynamics of Microorganisms Colonizing the Human Digestive Tract during In Vitro Fermentations in Apple Pomace Hydrolysate
3.3. Metabolism of Microorganisms Colonizing the Human Digestive Tract during In Vitro Fermentations in Apple Pomace Hydrolysate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGarr, S.E.; Ridlon, J.M.; Hylemon, P.B. Diet, anaerobic bacterial metabolism and colon cancer: A review of the literature. J. Clin. Gastroenterol. 2005, 39, 98–109. [Google Scholar] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- Biasucci, G.; Benenati, B.; Morelli, L.; Bessi, E.; Boehm, G. Cesarean delivery may affect the early biodiversity of intestinal bacteria. J. Nutr. 2008, 138, 1796–1800. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [Green Version]
- Bui, T.P.N.; Troise, A.D.; Nijsse, B.; Roviellof, G.N.; Foglianog, V.; de Vos, W.M. Intestinimonas-like bacteria are important butyrate producers that utilize Nε-fructosyllysine and lysine in formula-fed infants and adults. J. Funct. Foods 2020, 70, 103974. [Google Scholar] [CrossRef]
- Bezirtzoglou, E. The intestinal microflora during the first weeks of life. Anaerobe 1997, 3, 173–177. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Salazar, N.; Arboleya, S.; Valdes, L. The human intestinal microbiome at extreme ages of life. Dietary intervention as a way to counteract alterations. Front. Genet. 2014, 5, 1–9. [Google Scholar] [CrossRef]
- Biagi, E.; Nylund, L.; Candela, M.; Ostan, R.; Bucci, L.; Pini, E.; Nikkı, J.; Monti, D.; Satokari, R.; Franceschi, C.; et al. Through ageing, and beyond: Gut microbiota and inflammatory status in seniors and centenarians. PLoS ONE 2015, 5, e10667. [Google Scholar]
- Konturek, P.C.; Haziri, D.; Brzozowski, T.; Hess, T.; Heyman, S.; Kwiecien, S.; Konturek, S.J.; Koziel, J. Emerging role of fecalmicrobiota therapy in the treatment of gastrointestinal andextra-gastrointestinal diseases. J. Physiol. Pharmacol. 2015, 66, 483–491. [Google Scholar] [PubMed]
- Wegh, C.A.M.; Schoterman, M.H.C.; Vaughan, E.E.; Belzer, C.; Benning, M.A. The effect of fiber and prebiotics on children’s gastrointestinal disorders and microbiome. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 1031–1045. [Google Scholar] [CrossRef] [Green Version]
- Scheida, M.M.A.; Franco Moreno, Y.M.; Maróstica, M.R., Jr.; Pastorea, G.M. Effect of prebiotics on the health of the elderly. Food Res. Int. 2013, 53, 426–432. [Google Scholar] [CrossRef]
- Toward, R.; Montandon, S.; Walton, G.; Gibson, G.R. Effect of prebiotics on the human gut microbiota of elderly persons. Gut Microbes 2012, 3, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Gullón, B.; Garrote, G.; Alonso, J.L.; Parajo, J.C. Production of L-lactic acid and oligomeric compounds of apple pomace by simultaneous saccharification and fermentation: A response surface methodology assessment. J. Agric. Food Chem. 2007, 55, 5580–5587. [Google Scholar] [CrossRef]
- Hotchkiss, A.T., Jr.; Manderson, K.; Tuohy, K.M.; Widmer, W.W.; Nunez, A.; Gibson, G.R.; Rastall, R.A. Bioactive properties of pectic oligosaccharides from sugar beet and Valencia oranges. In Proceedings of the 233rd ACS National Meeting, Chicago, IL, USA, 25–29 March 2007. [Google Scholar]
- Rabetafika, H.N.; Bchir, B.; Blecker, C.H.; Richel, A. Fractionation of apple by-products as source of new ingredients: Current situation and perspectives. Trends Food Sci. Technol. 2014, 40, 99–114. [Google Scholar] [CrossRef]
- Gómez, B.; Gullón, B.; Yáñez, R.; Schols, H.; Alonso, J.L. Prebiotic potential of pectins and pectic oligosaccharides derived from lemon peel wastes and sugar beet pulp: A comparative evaluation. J. Funct. Foods 2016, 20, 108–121. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Wilkowska, A.; Nowak, A.; Antczak-Chrobot, A.; Motyl, I.; Czyżowska, A.; Paliwoda, A. Structurally different pectic oligosaccharides produced from apple pomace and their biological activity in vitro. Foods 2019, 8, 365. [Google Scholar] [CrossRef] [Green Version]
- Bonnin, E.; Clavurier, K.; Daniel, S.; Kauppinen, S.; Mikkelsen, J.D.M.; Thibault, J.-F. Pectin acetylesterases from Aspergillus are able to deacetylate homogalacturonan as well as rhamnogalacturonan. Carbohydr. Polym. 2008, 74, 411–418. [Google Scholar] [CrossRef]
- Gullon, B.; Gullon, P.; Sanz, Y.; Alonso, J.L.; Parajo, J.C. Prebiotic potential of a refined product containing pectic oligosaccharides. Food Sci. Tech. 2011, 44, 1687–1696. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, M.J.; Sharp, R.; Macfarlane, G.T. Variation in human intestinal microbiota with age. Digest Liver Dis. 2002, 34, 12–28. [Google Scholar] [CrossRef]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; de Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.; Fitzgerald, G.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2011, 108, 4586–4591. [Google Scholar] [CrossRef] [Green Version]
- Mack, D.R. D (–)-lactic acid-producing probiotics, D (–)-lactic acidosis and infants. Can. J. Gastroenterol. 2004, 18, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Cantarel, B.L.; Lombard, V.; Henrissat, B.; Hu, Y.; Walker, S.; Laine, R.; Varki, A.; Sharon, N.; Varki, A.; Hooper, L. Complex carbohydrate utilization by the healthy human microbiome. PLoS ONE 2012, 7, e28742. [Google Scholar] [CrossRef] [Green Version]
- El Kaoutari, A.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrateactive enzymes in the human gut microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, H.W.J.; Naylor, E.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieterich, W.; Schuppan, D.; Schink, M.; Schwappacher, R.; Wirtz, S.; Agaimy, A.; Neurath, M.F.; Zopf, Y. Influence of low FODMAP and gluten-free diets on disease activity and intestinal microbi;ota in patients with non-celiac gluten sensitivity. Clin. Nutr. 2018, 38, 697–707. [Google Scholar] [CrossRef]
- Dieterich, W.; Schink, M.; Zopf, Y. Microbiota in the Gastrointestinal Tract. Med. Sci. 2018, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S. Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell 2018, 174, 1388–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Jin, G.; Wang, G.; Liu, T.; Liu, X.; Wang, B.; Cao, H. Current Sampling Methods for Gut Microbiota: A Call for More Precise Devices. Front. Cell. Infect. Microbiol. 2020, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Bunesova, V.; Lacroix, C.; Schwab, C. Fucosyllactose and L-fucose utilization of infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol. 2016, 16, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asakuma, S.; Hatakeyama, E.; Urashima, T.; Yoshida, E.; Katayama, T.; Yamamoto, K.; Kumagai, H.; Ashida, H.; Hirose, J.; Kitaoka, M. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J. Biol. Chem. 2011, 286, 34583–34592. [Google Scholar] [CrossRef] [Green Version]
- Vandenplas, Y.; Berger, B.; Carnielli, V.P.; Ksiazyk, J.; Lagstrom, H.; Sanchez Luna, M.; Migacheva, N.; Mosselmans, J.-M.; Picaud, J.-C.; Possner, M.; et al. Human Milk Oligosaccharides: 2 -Fucosyllactose (2 -FL) and Lacto-N-Neotetraose (LNnT) in Infant Formula. Nutrients 2018, 10, 1161. [Google Scholar] [CrossRef] [Green Version]
- Hoeflinger, J.L.; Davis, S.R.; Chow, J.; Miller, M.J. In vitro impact of human milk oligosaccharides on Enterobacteriaceae growth. J. Agric. Food Chem. 2015, 63, 3295–3302. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.S.; Shin, S.Y.; Choi, Y.S.; Joo, W.; Cho, S.K.; Li, L.; Kang, J.-H.; Kim, T.-J.; Han, N.S. In vitro digestion and fermentation properties of linear sugar beet arabinan and its oligosaccharides. Carbohydr. Polym. 2015, 131, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Whisner, C.M.; Martin, B.R.; Schoterman, M.H.C.; Nakatsu, C.H.; McCabe, L.D.; McCabe, G.P.; Wastney, M.E.; van den Heuvel, E.G.H.M.; Weaver, C.M. Galacto-oligosaccharides increase calcium absorption and gut bifidobacteria in young girls: A double-blind cross-over trial. Br. J. Nutr. 2013, 110, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Vulevic, J.; Juric, A.; Walton, G.E.; Claus, S.P.; Tzortzis, G.; Toward, R.E.; Gibson, G.R. Influence of galacto-oligosaccharide mixture (B-GOS) on gut microbiota, immune parameters and metabonomics in elderly persons. Br. J. Nutr. 2015, 114, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Walton, G.E.; van den Heuvel, E.G.H.M.; Kosters, M.H.W.; Rastall, R.A.; Tuohy, K.M.; Gibson, G.R. A randomised crossover study investigating the effects of galacto-oligosaccharides on the faecal microbiota in men and women over 50 years of age. Br. J. Nutr. 2012, 107, 1466–1475. [Google Scholar] [CrossRef]
- Woodmansey, E.J.; McMurdo, M.E.T.; Macfarlane, G.T.; Macfarlane, S. Comparison of compositions and metabolic activities of fecal microbiotas in young adults and in antibiotic-treated and non-antibiotic-treated elderly subjects. Appl. Environ. Microbiol. 2004, 70, 6113–6122. [Google Scholar] [CrossRef] [Green Version]
- Rondanelli, M.; Giacosa, A.; Faliva, M.A.; Perna, S.; Allieri, F.; Castellazzi, A. Review on microbiota and effectiveness of probiotics use in older. World J. Clin. Cases 2015, 3, 156–162. [Google Scholar] [CrossRef]
- Nowak, A.; Czyżowska, A.; Huben, K.; Sójka, M.; Kuberski, S.; Otlewska, A.; Śliżewska, K. Prebiotics and age, but not probiotics affect transformation of 2-amino-3-methyl-3H-imidazo[4,5-f]quinoline (IQ) by faecal microbiota–An in vitro study. Anaerobe 2016, 39, 124–135. [Google Scholar] [CrossRef]
- Duncan, S.H.; Flint, H.J. Probiotics and prebiotics and health in ageing populations. Maturitas 2013, 75, 44–50. [Google Scholar] [CrossRef]
- Saffrey, M.J. Aging of the mammalian gastrointestinal tract: A complex organ system. Age 2014, 36, 1019–1032. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.; DuPont, H.L. New approaches for bacteriotheraphy: Prebiotics, new-generation probiotics, and synbiotics. Clinic. Inf. Dis. 2015, 60, 108–121. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Code | Target Microorganisms | Fluorescent Dye | Temperature [°C] | Time [h] |
|---|---|---|---|---|
| Bac 303 | Bacteroides/Prevotella sp. | 5′ Cy3 | 56 | 3 |
| Bif 164 | Bifidobacterium sp. | 5′ Cy3 | 58 | 18 |
| E. coli | Eschericha coli | 5′ Cy3 | 51.7 | 18 |
| Enter 1432 | Enterobacteriaceae | 5′ Cy3 | 43 | 18 |
| Erec 484 | Clostridium coccoides | 5′ Cy3 | 57 | 16 |
| Lab 158 | lactobacilli/Enterococcus sp. | 5′ Fluo | 45 | 24 |
| Host Age | Fermentation Period [days] | Lactic Acid Concentration [mg/100 g] | L (+)/D (−) |
|---|---|---|---|
| 7 years | 7 | 524.55 ± 26.93 a | 0.88 |
| 14 | 534.72 ± 32.40 a | 2.36 | |
| 24 years | 7 | 262.32 ± 15.72 b | 1.12 |
| 14 | 254.52 ± 14.11 b | 1.03 | |
| 47 years | 7 | 225.93 ± 11.65 bc | 0.85 |
| 14 | 210.64 ± 9.02 c | 1.15 | |
| 69 years | 7 | 213.89 ± 6.86 c | 2.09 |
| 14 | 257.14 ± 7.94 b | 2.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilkowska, A.; Motyl, I.; Antczak-Chrobot, A.; Wojtczak, M.; Nowak, A.; Czyżowska, A.; Motyl, W. Influence of Human Age on the Prebiotic Effect of Pectin-Derived Oligosaccharides Obtained from Apple Pomace. Fermentation 2021, 7, 224. https://doi.org/10.3390/fermentation7040224
Wilkowska A, Motyl I, Antczak-Chrobot A, Wojtczak M, Nowak A, Czyżowska A, Motyl W. Influence of Human Age on the Prebiotic Effect of Pectin-Derived Oligosaccharides Obtained from Apple Pomace. Fermentation. 2021; 7(4):224. https://doi.org/10.3390/fermentation7040224
Chicago/Turabian StyleWilkowska, Agnieszka, Ilona Motyl, Aneta Antczak-Chrobot, Maciej Wojtczak, Adriana Nowak, Agata Czyżowska, and Wojciech Motyl. 2021. "Influence of Human Age on the Prebiotic Effect of Pectin-Derived Oligosaccharides Obtained from Apple Pomace" Fermentation 7, no. 4: 224. https://doi.org/10.3390/fermentation7040224
APA StyleWilkowska, A., Motyl, I., Antczak-Chrobot, A., Wojtczak, M., Nowak, A., Czyżowska, A., & Motyl, W. (2021). Influence of Human Age on the Prebiotic Effect of Pectin-Derived Oligosaccharides Obtained from Apple Pomace. Fermentation, 7(4), 224. https://doi.org/10.3390/fermentation7040224