
The Evaluation of γ-Zein Reduction Using Mass Spectrometry—The Influence of Proteolysis Type in Relation to Starch Degradability in Silages
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fresh and High-Moisture Corn Samples
2.2. Degradability Determination
2.3. Data Analysis
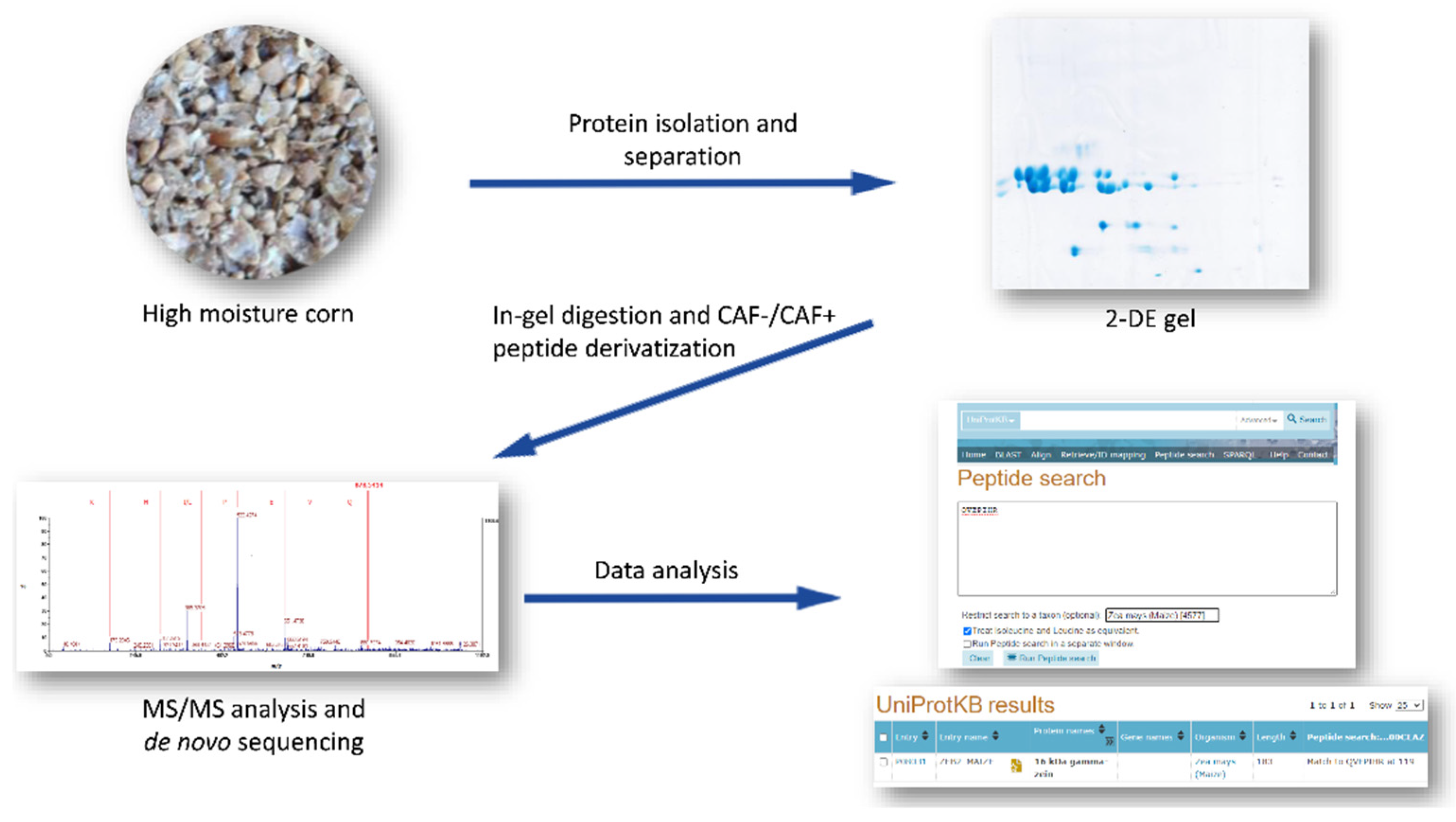
2.4. Gamma Zein Evaluation
2.4.1. Zein Extract Preparation
2.4.2. Two-Dimensional (2-D) Gel Electrophoresis with Zein Quantification
2.4.3. Sample Preparation for Mass Spectrometry
2.4.4. Mass Spectrometry and Protein Identification
2.5. Scanning Electron Microscopy Imaging of Ground Corn Samples
3. Results
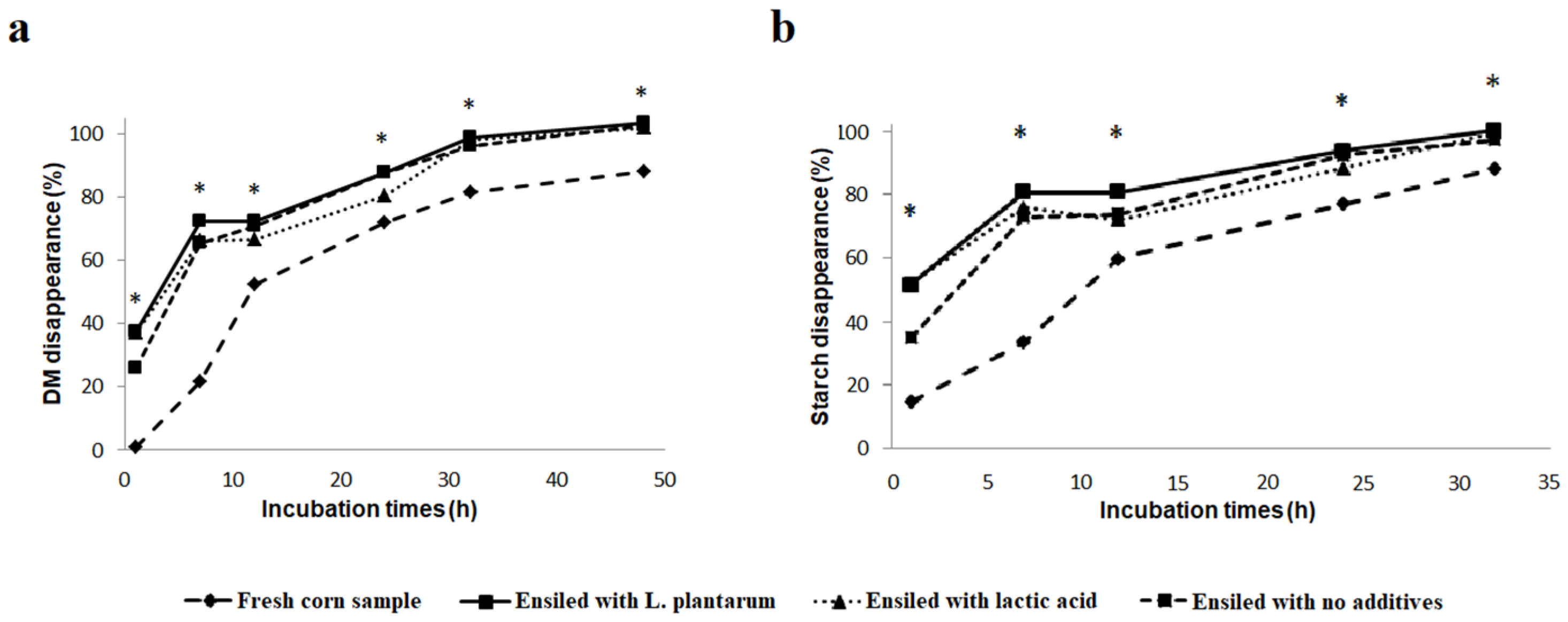
3.1. In Vitro Degradability Results
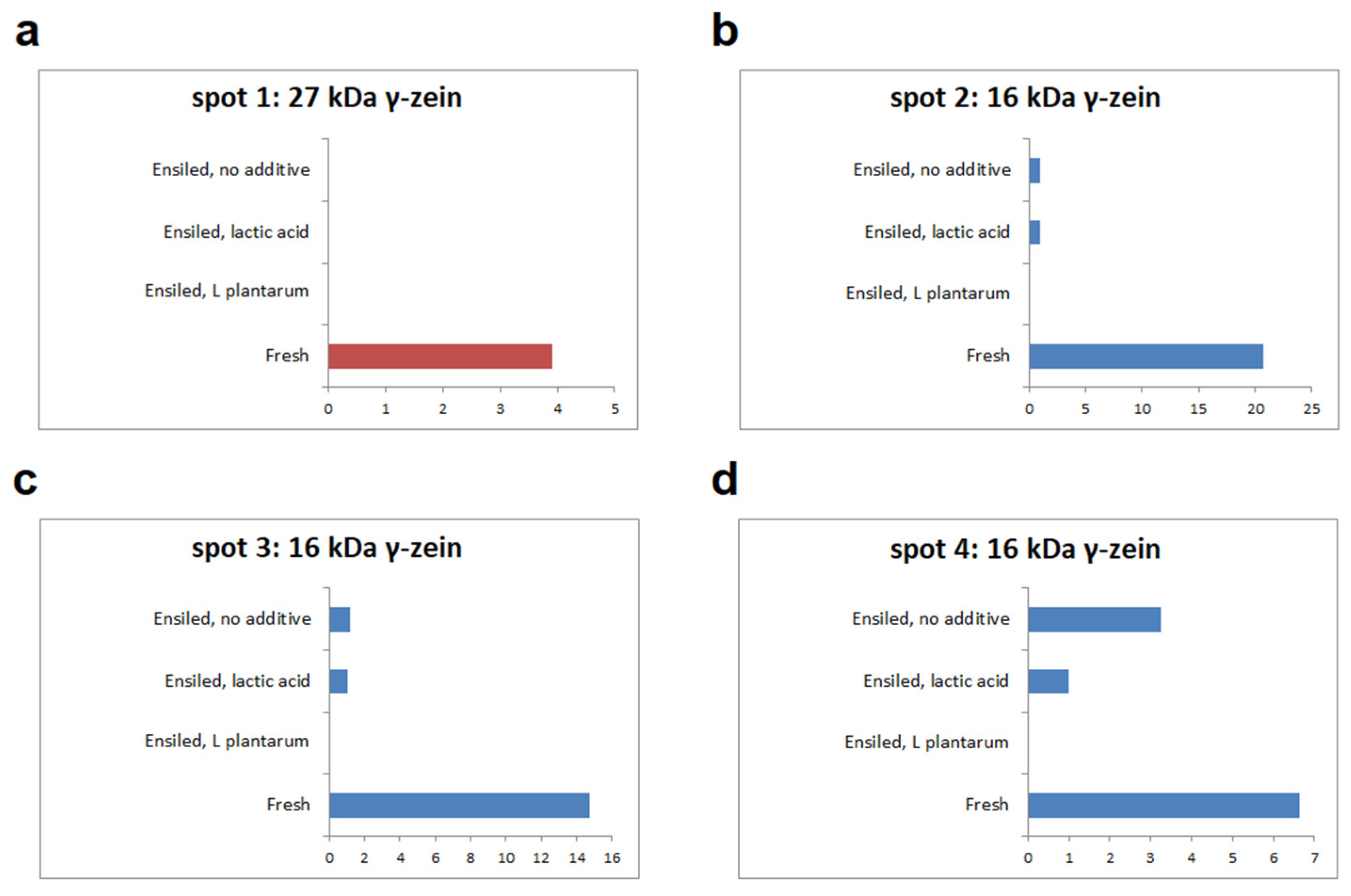
3.2. Zein Protein Pattern
3.3. γ-Zein Reduction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. Statistical Database. Available online: http://www.fao.org/faostat/en/#home (accessed on 30 July 2022).
- Holding, D.R.; Larkins, B.A. The development and importance of zein protein bodies in maize endosperm. Maydica 2006, 51, 243–254. [Google Scholar]
- Lawton, J.W. Zein: A history of processing and use. Cereal Chem. 2002, 79, 1–18. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Tapia-Hernández, J.A.; Del-Toro-Sánchez, C.L.; Cinco-Moroyoqui, F.J.; Juárez-Onofre, J.E.; Ruiz-Cruz, S.; Carvajal-Millan, E.; López-Ahumada, G.A.; Castro-Enriquez, D.D.; Barreras-Urbina, C.G.; Rodríguez-Felix, F. Prolamins from cereal by-products: Classification, extraction, characterization and its applications in micro-and nanofabrication. Trends Food Sci. Technol. 2019, 90, 111–132. [Google Scholar] [CrossRef]
- Song, J.; Sun, C.; Gul, K.; Mata, A.; Fang, Y. Prolamin-based complexes: Structure design and food-related applications. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1120–1149. [Google Scholar] [CrossRef]
- Giuberti, G.; Gallo, A.; Masoero, F.; Ferraretto, L.F.; Hoffman, P.C.; Shaver, R.D. Factors affecting starch utilization in large animal food production system: A review. Starch-Stärke 2014, 66, 72–90. [Google Scholar] [CrossRef]
- Kljak, K.; Duvnjak, M.; Grbeša, D. Effect of starch properties and zein content of commercial maize hybrids on kinetics of starch digestibility in an in vitro poultry model. J. Sci. Food Agric. 2019, 99, 6372–6379. [Google Scholar] [CrossRef]
- Shukla, R.; Cheryan, M. Zein: The industrial protein from corn. Ind. Crops Prod. 2001, 13, 171–192. [Google Scholar] [CrossRef]
- Ai, Y.; Jane, J.L. Macronutrients in corn and human nutrition. Compr. Rev. Food Sci. Food Saf. 2016, 15, 581–598. [Google Scholar] [CrossRef] [Green Version]
- Gibbon, B.C.; Larkins, B.A. Molecular genetic approaches to developing quality protein maize. Trends Genet. 2005, 21, 227–233. [Google Scholar] [CrossRef]
- Wu, Y.; Holding, D.R.; Messing, J. γ-Zeins are essential for endosperm modification in quality protein maize. Proc. Natl. Acad. Sci. USA 2010, 107, 12810–12815. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Messing, J. RNA interference-mediated change in protein body morphology and seed opacity through loss of different zein proteins. Plant Physiol. 2010, 153, 337–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holding, D.R. Recent advances in the study of prolamin storage protein organization and function. Front. Plant Sci. 2014, 5, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, Y.M.; Hu, D.W.N.; Larkins, B.A.; Jung, R. Genomics analysis of genes expressed in maize endosperm identifies novel seed proteins and clarifies patterns of zein gene expression. Plant Cell 2001, 13, 2297–2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, P.; Henderson, N.; Heron, S. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Muck, R.E.; Nadeau, E.M.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Philippeau, C.; Michalet-Doreau, B. Influence of genotype and ensiling of corn grain on in situ degradation of starch in the rumen. J. Dairy Sci. 1998, 81, 2178–2184. [Google Scholar] [CrossRef]
- Hoffman, P.C.; Esser, N.M.; Shaver, R.D.; Coblentz, W.; Scott, M.P.; Bodnar, A.L.; Schmidt, R.J.; Charley, R.C. Influence of ensiling time and inoculation on alteration of the starch-protein matrix in high-moisture corn. J. Dairy Sci. 2011, 94, 2465–2474. [Google Scholar] [CrossRef] [Green Version]
- Duvnjak, M.; Kljak, K.; Grbeša, D. Effect of hybrid, inoculant and storage time on whole plant nitrogen compounds and grain total zein content in maize silage. J. Anim. Feed Sci. 2016, 25, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.C.; Lopes, M.A.; Paiva, E.; Larkins, B.A. New methods for extraction and quantitation of zeins reveal a high content of γ-zein in modified opaque-2 maize. Plant Physiol. 1990, 92, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Shi, J.; Sun, C.; Gong, H.; Fan, X.; Qiu, F.; Huang, X.; Feng, Q.; Zheng, X.; Yuan, N.; et al. Gene duplication confers enhanced expression of 27-kDa γ-zein for endosperm modification in quality protein maize. Proc. Natl. Acad. Sci. USA 2016, 113, 4964–4969. [Google Scholar] [CrossRef] [Green Version]
- Postu, P.A.; Ion, L.; Drochioiu, G.; Petre, B.A.; Glocker, M.O. Mass spectrometric characterization of the zein protein composition in maize flour extracts upon protein separation by SDS-PAGE and 2D gel electrophoresis. Electrophoresis 2019, 40, 2747–2758. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.R.; Huang, S.; Kriz, A.L.; Luethy, M.H. Matrix-assisted laser desorption ionization time-of-flight mass spectrometry analysis of zeins in mature maize kernels. J. Agric. Food Chem. 2004, 52, 1842–1849. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zheng, H.; Lin, M.; Zhu, W.; Zhang, J. Characterization of the protein and peptide of excipient zein by the multi-enzyme digestion coupled with nano-LC-MS/MS. Food Chem. 2020, 321, 126712. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.; Mertens, D.R. Chemical and physical characteristics of corn silages and their effects on in vitro disappearance. J. Dairy Sci. 2005, 88, 4414–4425. [Google Scholar] [CrossRef] [Green Version]
- Goering, H.K.; Van Soest, P.J. Forage Fiber Analyses: Apparatus, Reagents, Procedures, and Some Applications; Agricultural Handbook no. 379; USDA Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1970.
- Ørskov, E.R.; McDonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Dhanoa, M.S. On the analysis of dacron bag data for low degradability feeds. Grass Forage Sci. 1988, 43, 441–444. [Google Scholar] [CrossRef]
- Peyrat, J.; Nozière, P.; Le Morvan, A.; Férard, A.; Protin, P.V.; Baumont, R. Effects of ensiling maize and sample conditioning on in situ rumen degradation of dry matter, starch and fibre. Anim. Feed Sci. Technol. 2014, 196, 12–21. [Google Scholar] [CrossRef]
- Saxton, A.M. A macro for converting mean separation output to letter groupings in Proc Mixed. In Proceedings of the 23rd SAS Users Group International, Nashville, TN, USA, 22–25 March 1998; SAS Institute Incorporated: Cary, NC, USA, 1998; pp. 1243–1246. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Butorac, A.; Dodig, I.; Bačun-Družina, V.; Tishbee, A.; Mrvčić, J.; Hock, K.; Diminić, J.; Cindrić, M. The effect of starvation stress on Lactobacillus brevis L62 protein profile determined by de novo sequencing in positive and negative mass spectrometry ion mode. Rapid Commu. Mass Spectrom. 2013, 27, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, A.T.; Salmi, J.M.; Aittokallio, T.; Westerholm, J.; Lahesmaa, R.; Nyman, T.A.; Nevalainen, O.S. Comparison of PDQuest and Progenesis software packages in the analysis of two-dimensional electrophoresis gels. Proteomics 2003, 10, 1936–1946. [Google Scholar] [CrossRef]
- Kotarski, S.F.; Waniska, R.D.; Thurn, K.K. Starch hydrolysis by the ruminal microflora. J. Nutr. 1992, 122, 178–190. [Google Scholar] [CrossRef]
- Philippeau, C.; Landry, J.; Michalet-Doreau, B. Influence of the protein distribution of maize endosperm on ruminal starch degradability. J. Sci. Food Agric. 2000, 80, 404–408. [Google Scholar] [CrossRef]
- Tsai, C.Y. Note on the effect of reducing agent on zein preparation. Cereal Chem. 1980, 57, 288–290. [Google Scholar]
- Guo, X.; Yuan, L.; Chen, H.; Sato, S.J.; Clemente, T.E.; Holding, D.R. Nonredundant function of zeins and their correct stoichiometric ratio drive protein body formation in maize endosperm. Plant Physiol. 2013, 162, 1359–1369. [Google Scholar] [CrossRef] [Green Version]
- Junges, D.; Morais, G.; Spoto, M.H.; Santos, P.S.; Adesogan, A.T.; Nussio, L.G.; Daniel, J.L.P. Influence of various proteolytic sources during fermentation of reconstituted corn grain silages. J. Dairy Sci. 2017, 100, 9048–9051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaji, M.; Matsuyama, H.; Nonaka, K. Effect of ensiling process of total mixed ration on fermentation profile, nutrient loss and in situ ruminal degradation characteristics of diet. Anim. Sci. J. 2017, 88, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Opsi, F.; Fortina, R.; Borreani, G.; Tabacco, E.; López, S. Influence of cultivar, sowing date and maturity at harvest on yield, digestibility, rumen fermentation kinetics and estimated feeding value of maize silage. J. Agric. Sci. 2013, 151, 740–753. [Google Scholar] [CrossRef] [Green Version]
- McAllister, T.A.; Cheng, K.J. Microbial strategies in the ruminal digestion of cereal grains. Anim. Feed Sci. Technol. 1996, 62, 29–36. [Google Scholar] [CrossRef]
- Nozière, P.; Michalet-Doreau, B. Validation of in sacco method: Influence of sampling site, nylon bag or rumen contents, on fibrolytic activity of solid-associated microorganisms. Anim. Feed Sci. Technol. 1996, 57, 203–210. [Google Scholar] [CrossRef]
- Van Milgen, J.; Baumont, R. Models based on variable fractional digestion rates to describe ruminal in situ digestion. Br. J. Nutr. 1995, 73, 793–807. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| High-Moisture Corn | p | ||||||
|---|---|---|---|---|---|---|---|
| Kinetics Parameters | Fresh Corn | Inoculant L. plantarum | Lactic Acid | Control | Ensiling (E) | Additive (A) | E×A |
| DM 6 degradability parameters | |||||||
| ED 1 | 0.59 b | 0.83 a | 0.76 a | 0.80 a | <0.001 | 0.302 | 0.302 |
| a 2 | 0.28 b | 0.58 a | 0.52 a | 0.58 a | <0.001 | 0.579 | 0.579 |
| b 3 | 0.69 a | 0.47 b | 0.47 b | 0.46 b | <0.001 | 0.995 | 0.995 |
| c (h−1) 4 | 0.0561 b | 0.0892 a | 0.0453 b | 0.0782 a | 0.002 | 0.003 | 0.001 |
| lag (h) 5 | 7.28 a | 4.01 b | 3.33 b | 6.02 a | <0.001 | 0.142 | 0.142 |
| Starch degradability parameters | |||||||
| ED 1 | 0.56 b | 0.83 a | 0.78 a | 0.79 a | <0.001 | 0.529 | 0.530 |
| a 2 | 0.36 c | 0.64 ab | 0.60 b | 0.68 a | <0.001 | 0.214 | 0.214 |
| b 3 | 0.63 a | 0.37 b | 0.41 b | 0.33 b | <0.001 | 0.693 | 0.693 |
| c (h−1) 4 | 0.0527 b | 0.1368 a | 0.0638 b | 0.0835 b | <0.001 | 0.035 | 0.024 |
| lag (h) 5 | 6.54 a | 3.03 b | 2.34 b | 5.44 a | 0.003 | 0.002 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duvnjak, M.; Butorac, A.; Kljak, K.; Nišavić, M.; Cindrić, M.; Grbeša, D. The Evaluation of γ-Zein Reduction Using Mass Spectrometry—The Influence of Proteolysis Type in Relation to Starch Degradability in Silages. Fermentation 2022, 8, 551. https://doi.org/10.3390/fermentation8100551
Duvnjak M, Butorac A, Kljak K, Nišavić M, Cindrić M, Grbeša D. The Evaluation of γ-Zein Reduction Using Mass Spectrometry—The Influence of Proteolysis Type in Relation to Starch Degradability in Silages. Fermentation. 2022; 8(10):551. https://doi.org/10.3390/fermentation8100551
Chicago/Turabian StyleDuvnjak, Marija, Ana Butorac, Kristina Kljak, Marija Nišavić, Mario Cindrić, and Darko Grbeša. 2022. "The Evaluation of γ-Zein Reduction Using Mass Spectrometry—The Influence of Proteolysis Type in Relation to Starch Degradability in Silages" Fermentation 8, no. 10: 551. https://doi.org/10.3390/fermentation8100551