Abstract
Pomegranate-seed residue (PSR) was used in a new strategy for the simultaneous production of Yarrowia lipolytica lipase by submerged fermentation and its immobilization by adsorption. This biocatalyst—the fermented solid residue containing the adsorbed lipase (fermPSR)—was evaluated in hydrolysis reactions and in structured lipid synthesis. In shake flasks, yeast extract and urea were the best nitrogen sources for lipase production with PSR and their simultaneous use increased the lipase production even further. This result was confirmed in a 3.5-liter bioreactor, with lipase activity in an extracellular medium of 40 U/mL. A maximum reaction rate (Vmax) of 49.5 µmol/min/g, a Michaelis–Menten constant (Km) of 207 µmol/L, and a turnover number (Kcat) of 130 s−1 were determined for the new biocatalyst, fermPSR, for the hydrolysis of p-nitrophenyl laurate (p-NPL) into p-nitrophenol. The conversion of p-NPL into p-nitrophenol in subsequent reactions confirmed fermPSR’s potential for industrial hydrolytic reactions. The production of structured lipids from vegetable oil and free fatty acids by fermPSR evidences the versatility of this new biocatalyst.
1. Introduction
Pomegranate (Punica granatum L.) is a berry-fruit-bearing plant species cultivated in several regions of the world, including tropical Africa, South America, and Europe. During its processing, about 60% of residues are produced, including seeds (10%) [1]. These seeds contain 12 to 20% oil [2,3,4], with a unique fatty-acid profile, especially punicic acid, which represents 70 to 76% of total fatty acids. Unsaturated fatty acids, such as oleic and linoleic acids, also contribute to this profile [5]. Considering the importance of the rich fatty-acid composition of pomegranate oil for human health, many studies have focused on its extraction [6,7]. However, some of these compounds still remain in the residual solid, which can be used as raw material for microbial growth and to obtain high-value products [8], such as lipases [9].
Lipases can catalyze the hydrolysis of triglycerides, and in low amounts of water, they can also catalyze the synthesis of various esters [10,11]. These enzymes can be produced in fermentation processes by several microorganisms, such as the yeast Yarrowia lipolytica. This species is non-pathogenic and, therefore, GRAS (generally regarded as safe) products can be obtained from its metabolism [12].
Lipases can be used in free or immobilized form. The latter form is preferable because of several advantages, such as higher enzyme stability and potential reuse, which reduces the enzymatic-process cost [9,13]. An interesting approach to immobilization is the spontaneous adsorption of enzymes into solid residue during the fermentation process, which has been reported in recent years with promising results [9,13,14,15]. The solid residues of the culture medium also work as supports that keep the enzyme adsorbed [13]. Thus, during the fermentation of agro-industrial waste using submerged fermentation, two products with added value are obtained with this new strategy, the crude enzymatic extract (free enzyme) and the fermented solid containing the adsorbed enzyme (immobilized enzyme), which is often discarded after fermentation.
Therefore, this work aimed to explore this new strategy for the simultaneous production (submerged fermentation) and immobilization (by adsorption) of Y. lipolytica lipase using pomegranate-seed residue and to evaluate the potential use of the fermented solid residue containing the adsorbed lipase as a biocatalyst in hydrolysis reactions and structured lipid synthesis. Subsequent reactions with the biocatalyst were also tested to determine its reuse.
2. Materials and Methods
2.1. Materials
Pomegranate-seed residue (PSR) used in the fermentations was provided by the Food and Nutritional Biochemistry Laboratory of the Federal University of Rio de Janeiro, after a study of oil extraction [16]. PSR was previously dried in a Fabbe-Primar drying oven (model 219) at 65 °C and then ground in a laboratory mill. The flour was sieved in a 0.80-millimeter-mesh-size sieve and stored in polyethylene bags until its use. Peptone and yeast extract used was from Kasvi (Paraná, Brazil) and urea, ammonium chloride, ammonium sulfate, and potassium phosphate were from Sigma Aldrich Brazil. The tryptone was from Himedia (Mumbai, India) and corn-seep liquor was obtained from Ingredion Brazil. The p-nitrophenyllaurate (p-NFL) was obtained from Sigma Aldrich Brazil and dimethyl sulfoxide was obtained from Isofar (Rio de Janeiro, Brazil).
2.2. Physicochemical Characterization of PSR
Proximate composition analysis of PSR was performed according to the methods described by Institute Adolfo Lutz [17] and Official Methods of Analysis of AOAC International [18]. The following parameters were determined: moisture, lipids, ash, proteins, and total carbohydrates by difference (Nifext). All analyses were performed in triplicate.
2.3. Lipase Production
2.3.1. Strain and Inoculum Preparation
A wild-type strain of Yarrowia lipolytica (IMUFRJ 50682), isolated from Guanabara Bay, Rio de Janeiro (Brazil), was used to produce the lipase [19]. Cells were pre-cultivated in YPD medium (w/v: yeast extract 1%; peptone, 2%; glucose, 2%), at 28 °C, 160 rpm, for 72 h. The pre-cultured cells were used in sufficient amounts to start lipase production with 1 g of cells (dry weight)/L.
2.3.2. Lipase Production in Erlenmeyer Flasks
Lipase production in Erlenmeyer flasks was performed in 250-milliliter flasks containing 60 mL of production medium. In preliminary experiments, lipase-production medium was prepared with distilled water and PSR, 25 g/L, with and without different organic and inorganic nitrogen sources (urea, ammonium chloride, ammonium sulfate, corn-steep liquor, yeast extract, tryptone, soy tryptone, peptone, peptone/extract yeast, and urea/yeast extract) in a fixed concentration of 2 g/L. The combination of the best sources was preliminarily tested in increasing concentrations (until 4 g/L). Next, a central composite rotatable design (CCD) 22, with three replications at the central point, was used to evaluate the ideal concentrations of the best nitrogen sources (urea and yeast extract) for lipase production with PSR (Table 1).

Table 1.
Matrix of experimental runs for the central composite rotatable design for lipase production by Yarrowia lipolytica cultivated in Erlenmeyer flasks for 16 h in pomegranate-seed residue (PSR) supplemented with yeast extract and urea.
All these experimental samples were inoculated with 1 g dry weight of cells/L and incubated at 28 °C in a rotary shaker at 250 rpm for 70 h. Samples were collected during the bioprocess to determine extracellular lipolytic activity.
2.3.3. Lipase Production in Bioreactor
The best culture medium to produce lipase, determined in Erlenmeyer flasks, was used in larger-scale production process, with a New Brunswick Microferm MF-114 bioreactor (effective volume of 3.5 L). Lipase production was carried out with mechanical agitation of 600 rpm, aeration rate of 1.5 L.min−1 and temperature of 28 °C, for 90 h. Samples taken every 6 h were centrifuged at 4 °C, 2000× g for 5 min, and the supernatant was collected and frozen (−20 °C) for further evaluation of extracellular lipase activity.
Once the time for maximum extracellular lipase activity was determined in experiments that lasted 90 h with sampling amount of 5 mL, lipase production was performed again in the same conditions, but without taking samples and stocking all the cell-free medium and the fermented solid residue after the fixed time. This procedure was followed to obtain a larger volume of crude lipase extract with high activity and the fermented pomegranate seed residue (fermPSR) was freeze-dried (LC 1500, Terroni, São Carlos, Brazil) for evaluation as an immobilized biocatalyst.
2.4. Freeze-Dried Fermented-Pomegranate-Seed Residue (fermPSR) Characterization
2.4.1. Physicochemical Characterization of Freeze-Dried fermPSR
Freeze-dried fermPSR was evaluated for protein content (described in Section 2.2) and microscopic morphology in an optical microscopic (Nikon, model Eclipse E100LED) to check the possible adhesion of cells on the surface of the material.
2.4.2. Kinetic-Biocatalyst Parameters of Freeze-Dried fermPSR
Kinetic parameters, Michaelis–Menten constant (Km), and maximum reaction rate (Vmax) of freeze-dried fermPSR were determined from hydrolysis reactions of p-nitrophenyl laurate (p-NPL), with substrate concentrations ranging from 20 to 3000 μM. The extracellular crude extract was also tested for comparison. The hydrolysis reaction was carried out in a 1-milliliter cuvette in a spectrophotometer (Shimadzu model UV-1800) at 37 °C for 100 s and the absorbance at 410 nm was recorded to monitor p-nitrophenol production. Km and Vmax were obtained using a nonlinear regression of reaction rate versus substrate-concentration data using the Michaelis–Menten kinetic model. The substrate’s binding free energy (ΔGE-S), the renewal number (Kcat), and the transition-state free energy (ΔGE-T) were also determined, as shown in Equations (1)–(3) [20].
where Et is the protein concentration (μmol/mL), R is the universal ideal gas constant (8.314 × 10−3 kJ/mol.K), and T is the experimental temperature (310 K).
2.4.3. Hydrolysis and Synthesis Reactions with Freeze-Dried fermPSR as Biocatalyst
For the evaluation of freeze-dried fermPSR as an immobilized biocatalyst, experimental hydrolysis and synthesis reactions were performed. The hydrolysis experiments were performed using p-nitrophenyl laurate (p-NPL) as substrate, following the procedure described for lipase-activity analysis (Section 2.6.2). For the synthetic experiments, the production of structured lipids by acidolysis reaction with olive oil and lauric acid (12:0) was determined following the methodology described by Akil et al. [21]. The reactions were initiated by the addition of 200 mg of freeze-dried fermPSR in 50-milliliter mini-reactors containing 3.56 g of TAGs (olive oil) and 1.6 g of lauric acid (2:1 molar ratio: lauric acid: olive oil). A control experiment was performed with this same proportion of TAG and lauric acid, but without the enzyme, which was subjected to the same conditions as described here for the reactions with enzyme. Mini-reactors were kept at 37 °C for 15 min with magnetic stirring at 500 rpm. Next, the reacted substrate was collected and 3.0 mL of 5% Na2CO3 (w/v) and 7.0 mL of methanol (99.99%) were added to stop the reaction and encourage the separation of structured lipids (SL) of free fatty acids. The reaction medium was centrifuged at 3000× g for 10 min at 25 °C and the upper phase containing the SL was stored. The fatty-acid composition of the structured lipid formed was determined by gas chromatography. The amount of lauric acid obtained in the control experiment was deducted from the results with the enzyme.
2.5. Reuse of Freeze-Dried fermPSR in Biocatalysis Reactions
The reuse of freeze-dried fermPSR as a biocatalyst was performed by determining product concentration (p-nitrophenol) after successive reactions (4 cycles). These reactions were performed according to Section 2.4.3, and at the end of each cycle, the reaction system (substrate + biocatalyst) was transferred to a 50-milliliter falcon tube and centrifuged at 2000× g for 10 min. Next, the supernatant was collected, and the precipitate was used in a new cycle. The amount of product formed in the first cycle was defined as 100%. To evaluate desorption of the enzyme during reuse, fermPSR was subjected to magnetic stirring (500 rpm) in mini reactors with phosphate buffer at 37 °C for 60 min. Protein concentration was determined every 5 min.
2.6. Analytical Procedures
2.6.1. Cell Concentration
Cell concentration was determined by measuring the optical density in a spectrophotometer, with subsequent conversion to cell dry weight using a standard curve.
2.6.2. Lipolytic Activity
Lipase activity of the extracellular crude-enzyme extract was determined by monitoring the absorbance of p-nitrophenol production at 410 nm in a spectrophotometer (Shimadzu model UV-1800) at 37 °C for 100 s as a result of the hydrolysis of p-nitrophenyl laurate (p-NPL) [22]. For the freeze-dried fermented-pomegranate-seed residue (fermPSR), lipase activity was examined by adding 25 mL of p-NPL (560 μM) to 5 mg of freeze-dried fermPSR. This system was kept under magnetic stirring at 37 °C for 5 min and the reaction monitored by checking the absorbance (410 nm) at intervals of 30 s for 5 min. One unit of lipase activity is equivalent to the amount of enzyme required to produce 1 μmol p-nitrophenol/min.
2.6.3. Determination of Fatty-Acid Composition by Gas Chromatography (GC)
The fatty-acid composition of the structured lipid produced during synthesis reactions was determined by GC in a GC-2010 gas chromatograph (Shimadzu, Japan) equipped with a split/splitless inlet, in an Omegawax GC column (30 m×0.32 mm ID and 0.25 µm film thickness) and a flame-ionization detector (FID). Fatty-acid methyl esters (FAMEs) were prepared according to Lepage et al. [23] and injected into the GC. The chromatographic peak identities of the samples were determined based on retention time from commercial FAME standards (Supelco 37 FAME mix, Sigma-Aldrich, Brazil). The efficiency of incorporation (%) of lauric acid into the structured lipids produced was calculated from the peak area.
2.7. Statistical Analysis
The experimental data obtained were analyzed statistically using the software STATISTICA 7.1 (StatSoft, Inc., Tulsa, OK, USA). The obtained models were statistically verified by analysis of variance (ANOVA) and the significance determined by Fisher’s statistical test (Test F), using a 5% significance level. The variance explained by the models was observed by the coefficient of determination (R2).
3. Results and Discussion
3.1. Proximate Composition of Pomegranate Seed Residue (PSR)
Figure 1 shows the image of the PSR after oil extraction and after drying and milling processes. This flour (Figure 1b) was used for lipase production in a submerged culture. The proximate composition of PSR flour revealed a higher carbohydrate content in relation to lipid and protein (Table 2). Still, lipid and protein contents were significant and could help provide nutrients and inducers for the lipase-production process.

Figure 1.
PSR after oil extraction (a) and milling for use in fermentations (b).
Table 2.
Proximate composition of PSR.
The content of pomegranate-seed oil (12 to 20%) reduced to 3.54 in the PSR, as expected, since the PSR was obtained through oil extraction [16]; however, some oil remained in this residue, which was potentially good for microbial lipase production.
Guzmán-Lorite et al. [24] extracted and characterized protein from PSR using two different methods: pressurized liquid extraction and high-intensity focused ultrasound. High-intensity focused ultrasound yielded 11.4 ± 0.9 g protein/100 g, which was higher than that obtained with the pressurized liquid extraction (8.1 ± 0.3 g protein/100 g). The PSR of the present study was obtained after supercritical extraction [16] and presented a higher concentration of protein (12.57%), which is also important for lipase production.
3.2. Lipase Production with Pomegranate-Seed Residue
A culture medium composed simply of distilled water and PSR was ineffective for lipase production (Figure 2), possibly due to deficiencies in some of the important elements for yeast growth and enzyme production, mainly vitamins and nitrogen. Similar results were also reported by Pereira et al. [9] when studying lipase production using mango residue. Thus, PSR was supplemented with different nitrogen sources. The results of lipase production are also shown in Figure 2.
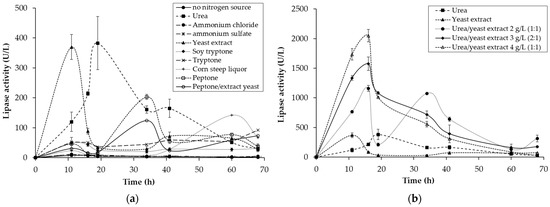
Figure 2.
Kinetic profile of extracellular lipase activity during submerged cultivation of Yarrowia lipolytica in Erlenmeyer flasks with pomegranate-seed residue (PSR) with and without supplementation of nitrogen sources: (a) testing of several nitrogen sources with PSR; (b) the best nitrogen sources (urea and yeast extract) with PSR and their combination.
The best sources of nitrogen (used individually) for the lipase production were yeast extract and urea, with a maximum production after 11 h for the yeast extract (369 U/L) and after 19 h for the urea (383 U/L) (Figure 2a). Yeast extract is effective at supplementing culture medium because it is a good nitrogen source and also provides vitamin components [25]. Galvagno et al. [26] and Novotný et al. [27] have also shown that yeast extract and urea significantly increase lipase production. The different maximum production times detected for the nitrogen sources (Figure 2a) were probably due to the secretion of lipases into the extracellular medium, which usually occur when the carbon source is scarce, and the nitrogen source exerts a direct influence on this process [28]. Ammonium chloride, ammonium sulfate, corn-steep liquor, tryptone, soy tryptone, and peptone were not effective at extracellular lipase production from PSR in the conditions studied. Even the combination of peptone and yeast extract, which are usually found in lipase-production media [28], was not as effective as the yeast extract or the urea. Lipase production with yeast extract and without PSR was carried out to verify whether this nitrogen source, which is also a carbon source, is sufficient for enzyme production. However, low lipolytic activity was detected (maximum 90 U/L after 20 h), evidencing that PSR is fundamental for lipase production.
As the yeast extract and urea were the nitrogen sources that led to the best lipase-production results, these compounds were tested simultaneously (Figure 2b). The results confirmed the importance of both nitrogen sources together (urea/yeast extract, 2 g/L (1:1); urea/yeast extract, 3 g/L (2:1); and urea/yeast extract, 4 g/L (1:1)), with the values of lipase activity increasing in line with their concentration (1160 U/L, 2 g/L; 1576 U/L, 3 g/L; 2051 U/L, 4 g/L). It seems that the lipase production was highest in the intersection (after 16 h) between the individual maximum production values (11 and 19 h), showing the synergistic effect of the compounds. In this sense, a central composite rotatable design was proposed to assess the influence of this synergistic effect on lipase production, as shown in Table 1.
The extracellular lipase activity ranged from 285.55 (5.9 g/L of nitrogen source) to 14,161.17 U/L (3.1 g/L of nitrogen source), which was significantly superior to the variation in the central points (7615.04 to 9990.86) (Table 1), indicating that the different concentrations of nitrogen sources influenced the lipase production. It is also possible to observe that when the urea concentration was higher than the yeast-extract concentration, the lipase activity reduced significantly (at least 16 times). The analysis of variance and the importance of the effects were verified using the Fisher test (F test), with a significance level of 10%. The model with the linear and quadratic effects of the variables was not significant and, therefore, non-significant terms were eliminated. This new ANOVA is presented in Table 3.
Table 3.
Analysis of variance (ANOVA) for the central composite rotatable design for extracellular lipase production by Y. lipolytica cultivated in Erlenmeyer flask for 16 h with pomegranate-seed residue (PSR) supplemented with yeast extract and urea.
The reduced model had no significant lack of fit (p > 0.10), which indicates that it adequately described the process (Table 3). The high value of R2 (0.913) also revealed a good correlation between the experimental and predicted data. Equation 4 represents the mathematical model with real variables (yeast extract and urea concentration) for the lipase activity.
where LA is the lipase activity in 16 h of fermentation, and YE and U represent the values of yeast extract and urea concentrations, respectively.
LA (U/L) = 19790.55 − 1470.61YE − 5424.52U
Due to the non-significance of the model’s quadratic terms, the optimum value for lipase production was not determined (Figure 3). However, there was a tendency towards an increase in lipase activity when the urea concentration tended to lower values (<0.5 g/L) and the yeast-extract concentration reached average values (±3.0 g/L). In these conditions, the lipase production was approximately 17,000 U/L.
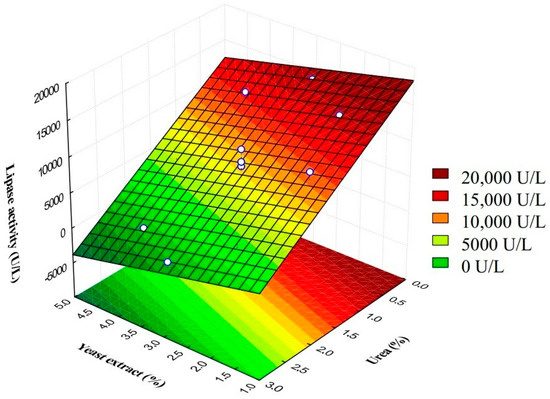
Figure 3.
Response surface of the central composite rotatable design (CCD) for extracellular lipase production by Y. lipolytica cultivated in Erlenmeyer flask for 16 h with PSR supplemented with yeast extract and urea.
The best combination of yeast extract and urea (3.0 g/L and 0.1 g/L, respectively) was used for the submerged cultivation of Y. lipolytica, a bioreactor with PSR, to produce lipase (Figure 4). An extracellular lipase activity of approximately 40,000 U/L was detected after 21 h. The second possible lipase peak (50–60 h) might have been related to the other lipases produced by this yeast, such as cell-wall-associated lipases, which can be released to extracellular medium when the carbon source is scarce [28]. However, the productivity would have been lower, which led to our choice of the peak at 21 h. The solid residue obtained after 21 h of cultivation in these conditions was centrifuged and separated from the supernatant containing the extracellular lipase, and from the cells that formed a layer between the supernatant and the solid. Subsequently, it was freeze-dried; the product, which is referred to in this study as freeze-dried-fermented-pomegranate-seed residue (freeze-dried fermPSR), was tested for lipase activity. A high hydrolytic activity in p-NPL was detected for this solid (69.83 ± 8.84 U/g) and, therefore, it was used as a biocatalyst in subsequent tests. A control test was performed with PSR and no activity was detected.
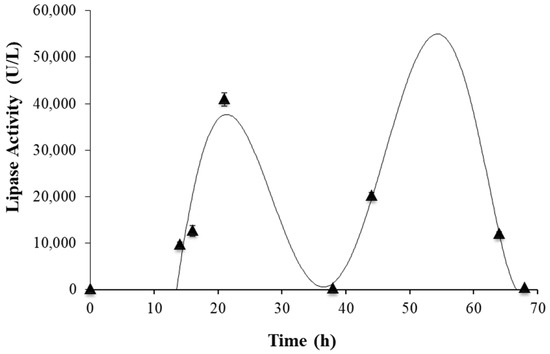
Figure 4.
Kinetic profile of extracellular lipase activity during submerged cultivation of Yarrowia lipolytica in 3.5-liter bioreactor with PSR with yeast extract (3 g/L) and urea (0.1 g/L).
3.3. Freeze-Dried Fermented-Pomegranate-Seed Residue (fermPSR) as a Biocatalyst
The protein concentration of the freeze-dried fermPSR was determined to verify whether protein enrichment from the adsorption of enzymes or cells occurred, but no significant difference was observed between the protein concentration before and after the fermentation process, probably because of the detection capacity of the method. An optical microscopic visualization of the PSR and fermPSR revealed no significant alteration in surface morphology after Y. lipolytica’s growth, not even the presence of adherent cells (data not shown). The enzyme activity associated with the cells separated from the solid showed no lipase. These results reinforce the argument that the enzyme activity found in the solid residue after the fermentation process was due to the physical adsorption of lipases on this support and not to the Y. lipolytica cells.
The influence of the substrate concentration (p-NPL) on the lipase activity of the fermPSR (immobilized biocatalyst) was determined. The Michaelis–Menten nonlinear kinetic model was used to obtain the apparent kinetic parameters Vmax and Km (Table 4). High values for the determination coefficients were found for the adjustment of this model (R2 = 0.978). The turnover number (Kcat), free-energy-substrate binding (ΔGE-S), and free energy of transition-state binding (ΔGE-T) were also calculated (Table 4).
Table 4.
Kinetic and thermodynamic parameters for the immobilized lipase (freeze-dried fermPSR).
The Vmax calculated per gram of biocatalyst (fermPSR) was low in comparison to the other Y. lipolytica immobilized lipases (175 U/mg of protein of lipase immobilized in chitosan–alginate beads [27], 731 U/g of lipase immobilized in cell debris [29], and 2904 U/g of lipase immobilized on magnetic nanoparticles [30]). However, for Km, a lower value (0.21 mM) was observed for the fermPSR in relation to the other immobilized Y. lipolytica lipases (2.52 mM, [29]; 0.93 mM, [30], which reveals the higher affinity of this biocatalyst towards the substrate. Pereira et al. [31] observed a reduction in enzyme-substrate affinity with the encapsulation of the Y. lipolytica lipase in the alginate and chitosan beads. Nevertheless, the Km obtained for their immobilized biocatalyst (0.04 mM) was lower than the fermPSR. The turnover number (Kcat) detected for the fermPSR was low, considering that for all the enzymes it ranged from 1 to 104 s−1, but it was not very far from the Y. lipolytica lipase encapsulated in the alginate and chitosan beads (606 s−1). The free-energy-substrate binding (ΔGE-S) revealed that the free lipase (8.7 kJ/μmol, [27]) needed a smaller amount of energy for the formation of the enzyme–substrate complex than the fermPSR. The free energy of the transition-state binding (ΔGE-T) indicated grater reaction spontaneity for the free lipase (−18.9 kJ/μmol, [27]). Nevertheless, the possibility of reusing the immobilized catalyst favors its choice.
3.4. Hydrolysis and Synthesis Reactions with Freeze-Dried fermPSR
The freeze-dried fermPSR was used in the p-NPL hydrolysis and in the structured lipids synthesis (olive oil + lauric acid). For the hydrolysis reaction, the production of approximately 120.0 ± 22.3 µmol/L of p-nitrophenol was achieved in 10 min of reaction, which corresponded to 24.4% of the possible theoretical yield.
Structured lipids (SLs) are triacylglycerols that have been restructured to change the position and composition of fatty acids from their original form [21]. In this test, we chose to produce SLs with a medium-chain fatty acid (lauric acid, 12:0) esterified at the sn-1 and sn-3 positions and a long-chain unsaturated fatty acid (oleic acid, 18:1n-9) esterified at the sn-2 position of the glycerol backbone. In this sense, we used olive oil—mostly composed of triacylglycerols with oleic acid in all positions—and lauric acid, a medium-chain fatty acid. By adding a strictly 1,3-regioselective lipase (Y. lipolytica lipase) [21], the lauric acid would probably have been incorporated exclusively in SL sn-1 and sn-3. For the structured-lipids synthesis, an incorporation efficiency of approximately 9.0 ± 1.9 % was observed after 15 min of reaction. Akil et al. [21] found an optimal incorporation efficiency of 30% when producing structured lipids using Y. lipolytica lipase immobilized in chitosan–alginate beads as a biocatalyst. Although the incorporation efficiency was not as high with the fermPSR, these experiments showed that it is possible to use this biocatalyst in structured-lipids synthesis. In addition, it is important to mention that an optimization of the reaction conditions was not performed for the fermPSR, which could have improved these results.
3.5. Reuse of Freeze-Dried fermPSR in Hydrolysis
To explore the reuse of the new biocatalyst, four reaction cycles were tested in the hydrolysis reactions. Figure 5 shows the reusability of the fermPSR in the p-NPL hydrolysis.
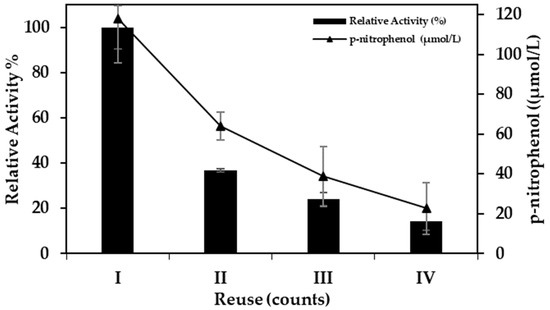
Figure 5.
Reusability of fermPSR in hydrolysis reaction with p-nitrophenyl laurate. The enzymatic activity of the first round was set to 100%.
The activity of the new biocatalyst reduced by more than 50% in the second reaction cycle and retained approximately 20% of its initial activity after the fourth reaction cycle. This is probably because the natural immobilization of the enzyme during fermentation occurred through physical adsorption, which resulted in a weak bond between the enzyme and the support. With a lower amount of catalyst in the support because of desorption, the increase in reaction time could have helped to achieve the product concentration of the first cycle. However, the p-nitrophenol concentration also reduced in the second cycle (approximately 60%) after 10 min. In successive reactions, it decreases to approximately 20%. A test was performed using magnetic-stirring mini-reactors with phosphate buffer and fermPSR at 37 °C (Figure 6). The increase in the protein concentration detected in the buffer evidences the desorption of the enzyme during the reactions. Despite the considerable reduction in enzymatic activity and product formation, the use of fermPSR in the hydrolysis reaction of the p-nitrophenyl laurate could have been advantageous, since it had already been used for microbial growth and was again used as a biocatalyst.
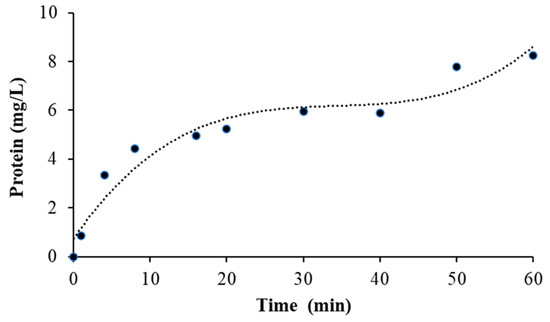
Figure 6.
Desorption test for the fermPSR in phosphate buffer at 37 °C with magnetic stirring of 500 rpm.
4. Conclusions
PSR without nitrogen supplementation was not sufficient to produce extracellular lipase. Yeast extract and urea were the best nitrogen sources tested to supplement PSR, with a lipase production of 369 U/L and 383 U/L, respectively. When these sources were combined, an activity of approximately 17,000 U/L and 40,000 U/L was obtained for Erlenmeyer flask and bioreactor cultures, respectively. A high hydrolytic activity in the p-NPL was detected in the solid residue after fermentation—fermPSR (69.83 U/g). The determination of the kinetic parameters of the fermPSR showed low Vmax and good affinity for the substrate (low Km) in comparison to other immobilized Y. lipolytica lipases. The use of the fermPSR in the hydrolysis of p-nitrophenyl laurate showed good results and the possibility of reusing this biocatalyst in these reactions. The production of structured lipids using the fermPSR was low, but possible. Therefore, the results presented here show that PSR has significant potential for use as a culture medium for extracellular lipase production and also as a support for the physical adsorption of enzymes and the formation of new biocatalysts.
Author Contributions
Conceptualization, A.d.S.P. and P.F.F.A.; methodology, A.d.S.P., P.F.F.A., M.M.D. and G.A.; software, A.d.S.P.; validation, A.d.S.P. and P.F.F.A.; formal analysis, A.d.S.P., M.M.D. and G.A.; investigation, A.d.S.P., P.F.F.A., M.M.D. and G.A; resources, A.d.S.P., P.F.F.A., M.M.D. and G.A.; data curation, A.d.S.P. and P.F.F.A.; writing—original draft preparation, A.d.S.P., M.M.D. and G.A.; writing—review and editing, A.d.S.P. and P.F.F.A.; visualization, A.d.S.P. and P.F.F.A.; supervision, P.F.F.A.; project administration, P.F.F.A.; funding acquisition, A.d.S.P. and P.F.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ—grant number E-26/201.915/2020), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Bolsa D.Sc.), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq—Bolsa PQ: 308626/2019-2).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aviram, M.; Dornfeld, L.; Rosenblat, M.; Volkova, N.; Kaplan, M.; Coleman, R.; Hayek, T.; Presser, D.; Fuhrman, B. Pomegranate juice consumption reduces oxidative stress, atherogenic modifications to LDL, and platelet aggregation: Studies in humans and in atherosclerotic apolipoprotein E-deficient mice. Am. J. Clin. Nutr. 2000, 71, 1062–1076. [Google Scholar] [CrossRef]
- Lansky, E.P.; Newman, R.A. Punica granatum (pomegranate) and its potential for prevention and treatment of inflammation and cancer. J. Ethnopharmacol. 2007, 109, 177–206. [Google Scholar] [CrossRef]
- Elfalleh, W.; Ying, M.; Nasri, N.; Sheng-Hua, H.; Guasmi, F.; Ferchichi, A. Fatty acids from Tunisian and Chinese pomegranate (Punica granatum L.) seeds. Int. J. Food Sci. Nutr. 2011, 62, 200–206. [Google Scholar] [CrossRef]
- Fernandes, L.; Pereira, J.A.; Lopéz-Cortés, I.; Salazar, D.M.; Ramalhosa, E.; Casal, S. Fatty acid, vitamin E and sterols composition of seed oils from nine different pomegranate (Punica granatum L.) cultivars grown in Spain. J. Food Compos. Anal. 2015, 39, 13–22. [Google Scholar] [CrossRef]
- Viladomiu, M.; Hontecillas, R.; Lu, P.; Bassaganya-Riera, J. Preventive and prophylactic mechanisms of action of pomegranate bioactive constituents. Evid. Based. Complement. Alternat. Med. 2013, 2013, 789764. [Google Scholar] [CrossRef]
- Goula, A.M. Ultrasound-assisted extraction of pomegranate seed oil—Kinetic modeling. J. Food Eng. 2013, 117, 492–498. [Google Scholar] [CrossRef]
- Đurđević, S.; Milovanović, S.; Šavikin, K.; Ristić, M.; Menković, N.; Pljevljakušić, D.; Petrović, S.; Bogdanović, A. Improvement of supercritical CO2 and n-hexane extraction of wild growing pomegranate seed oil by microwave pretreatment. Ind. Crops Prod. 2017, 104, 21–27. [Google Scholar] [CrossRef]
- Laufenberg, G.; Kunz, B.; Nystroem, M. Transformation of vegetable waste into value added products: (A) the upgrading concept; (B) practical implementations. Bioresour. Technol. 2003, 87, 167–198. [Google Scholar] [CrossRef]
- Da Pereira, A.S.; Fontes-Sant’Ana, G.C.; Amaral, P.F.F. Mango agro-industrial wastes for lipase production from Yarrowia lipolytica and the potential of the fermented solid as a biocatalyst. Food Bioprod. Process. 2019, 115, 68–77. [Google Scholar] [CrossRef]
- Treichel, H.; de Oliveira, D.; Mazutti, M.A.; Di Luccio, M.; Oliveira, J.V. A review on microbial lipases production. Food Bioprocess Technol. 2010, 3, 182–196. [Google Scholar] [CrossRef]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial lipases: An overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol. 2004, 64, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.H.; Ji, X.J.; Huang, H. Biotechnological applications of Yarrowia lipolytica: Past, present and future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef] [PubMed]
- Aguieiras, E.C.G.; Cavalcanti-Oliveira, E.D.; De Castro, A.M.; Langone, M.A.P.; Freire, D.M.G. Biodiesel production from Acrocomia aculeata acid oil by (enzyme/enzyme) hydroesterification process: Use of vegetable lipase and fermented solid as low-cost biocatalysts. Fuel 2014, 135, 315–321. [Google Scholar] [CrossRef]
- Todeschini, J.K.P.; Aguieiras, E.C.G.; de Castro, A.M.; Langone, M.A.P.; Freire, D.M.G.; Rodrigues, R.C. Synthesis of butyl esters via ultrasound-assisted transesterification of macaúba (Acrocomia aculeata) acid oil using a biomass-derived fermented solid as biocatalyst. J. Mol. Catal. B Enzym. 2016, 133, S213–S219. [Google Scholar] [CrossRef]
- Aguieiras, E.C.G.; de Barros, D.S.N.; Fernandez-Lafuente, R.; Freire, D.M.G. Production of lipases in cottonseed meal and application of the fermented solid as biocatalyst in esterification and transesterification reactions. Renew. Energy 2019, 130, 574–581. [Google Scholar] [CrossRef]
- De Silva, L.O.; Ranquine, L.G.; Monteiro, M.; Torres, A.G. Pomegranate (Punica granatum L.) seed oil enriched with conjugated linolenic acid (cLnA), phenolic compounds and tocopherols: Improved extraction of a specialty oil by supercritical CO2. J. Supercrit. Fluids 2019, 147, 126–137. [Google Scholar] [CrossRef]
- Zenebon, O.; Pascuet, N.S.; Tiglea, P. (Eds.) Métodos Físico-Químicos para Análise de Alimentos; Instituto Adolfo Lutz: São Paulo, Brazil, 2008. [Google Scholar]
- Horwitz, W.; George, W. Latimer Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Hagler, A.N.; Mendonça-Hagler, L.C. Yeasts from Marine and Estuarine Waters with Different Levels of Pollution in the State of Rio de Janeiro, Brazil. Appl. Environ. Microbiol. 1981, 41, 173. [Google Scholar] [CrossRef]
- Saqib, A.A.N.; Hassan, M.; Khan, N.F.; Baig, S. Thermostability of crude endoglucanase from Aspergillus fumigatus grown under solid state fermentation (SSF) and submerged fermentation (SmF). Process Biochem. 2010, 45, 641–646. [Google Scholar] [CrossRef]
- Akil, E.; da, S. Pereira, A.; El-Bacha, T.; Amaral, P.F.F.; Torres, A.G. Efficient production of bioactive structured lipids by fast acidolysis catalyzed by Yarrowia lipolytica lipase, free and immobilized in chitosan-alginate beads, in solvent-free medium. Int. J. Biol. Macromol. 2020, 163, 910–918. [Google Scholar] [CrossRef]
- Pereira-Meirelles, F.V.; Rocha-LeãO, M.H.M.; Santt’ Anna, G.L. A stable lipase from Candida lipolytica: Cultivation conditions and crude enzyme characteristics. Appl. Biochem. Biotechnol. 1997, 63–65, 73–85. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Direct transesterification of all classes of lipids in a one-step reaction. J. Lipid Res. 1986, 27, 114–120. [Google Scholar] [CrossRef]
- Guzmán-Lorite, M.; Marina, M.L.; García, M.C. Pressurized liquids vs. high intensity focused ultrasounds for the extraction of proteins from a pomegranate seed waste. Innov. Food Sci. Emerg. Technol. 2022, 77, 102958. [Google Scholar] [CrossRef]
- Li, C.Y.; Cheng, C.Y.; Chen, T.L. Fed-batch production of lipase by Acinetobacter radioresistens using Tween 80 as the carbon source. Biochem. Eng. J. 2004, 19, 25–31. [Google Scholar] [CrossRef]
- Galvagno, M.A.; Iannone, L.J.; Bianchi, J.; Kronberg, F.; Rost, E.; Carstens, M.R.; Cerrutti, P. Optimization of biomass production of a mutant of Yarrowia lipolytica with an increased lipase activity using raw glycerol. Rev. Argent. Microbiol. 2011, 43, 218–225. [Google Scholar] [CrossRef]
- Novotný, Č.; Doležalová, L.; Musil, P.; Novák, M. The production of lipases by some Candida and Yarrowia yeasts. J. Basic Microbiol. 1988, 28, 221–227. [Google Scholar] [CrossRef]
- Nunes, P.M.B.; Fraga, J.L.; Ratier, R.B.; Rocha-Leão, M.H.M.; Brígida, A.I.S.; Fickers, P.; Amaral, P.F.F. Waste soybean frying oil for the production, extraction, and characterization of cell-wall-associated lipases from Yarrowia lipolytica. Bioprocess Biosyst. Eng. 2021, 44, 809–818. [Google Scholar] [CrossRef]
- Fraga, J.L.; da Penha, A.C.B.; Akil, E.; Silva, K.A.; Amaral, P.F.F. Catalytic and physical features of a naturally immobilized Yarrowia lipolytica lipase in cell debris (LipImDebri) displaying high thermostability. 3 Biotech 2020, 10, 454. [Google Scholar] [CrossRef]
- Carvalho, T.; da, S. Pereira, A.; Bonomo, R.C.F.; Franco, M.; Finotelli, P.V.; Amaral, P.F.F. Simple physical adsorption technique to immobilize Yarrowia lipolytica lipase purified by different methods on magnetic nanoparticles: Adsorption isotherms and thermodynamic approach. Int. J. Biol. Macromol. 2020, 160, 889–902. [Google Scholar] [CrossRef]
- Da Pereira, A.S.; Diniz, M.M.; De Jong, G.; Gama Filho, H.S.; dos Anjos, M.J.; Finotelli, P.V.; Fontes-Sant’Ana, G.C.; Amaral, P.F.F. Chitosan-alginate beads as encapsulating agents for Yarrowia lipolytica lipase: Morphological, physico-chemical and kinetic characteristics. Int. J. Biol. Macromol. 2019, 139, 621–630. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).