Abstract
Paraprobiotics are potential agents for improving animal health and performance. This experiment investigated the effect of dietary supplementation of yeast (Saccharomyces cerevisiae) culture (YC) on the growth performance, nutrient digestibility, rumen development and microbiome of fattening sheep. Ninety male Hu sheep weighed 38 ± 1.47 kg were randomly assigned to three treatments: CON diet (basal diet), LYC diet (basal diet supplied with 10 and 20 g/d yeast culture at the early and late stages, respectively), and HYC diet (basal diet supplied with 20 and 40 g/d yeast culture at the early and late stages, respectively). Treatments (LYC or HYC) were sprinkled on the feed surface according to the required dosage before feeding the basal diet to each sheep throughout the trial. The trial included early (60 days) and later (30 days) fattening periods. The results showed that average daily gain and feed efficiency were higher (p < 0.05) in the LYC group compared with CON in later and whole stages. Digestibility of DM, OM, CP, NDF and ADF were higher (p < 0.05) in LYC and HYC compared with CON. The retained N, the utilization efficiency of N and the biological value of N were higher (p < 0.05) in LYC compared with CON and HYC. Rumen NH3-N was higher (p < 0.05) in LYC and HYC. The papillary height of the rumen was higher (p < 0.05) in LYC when compared with CON and HYC, whereas rumen wall thickness and muscular layer thickness were higher (p < 0.05) in HYC compared with CON and LYC. The dressing percentage of LYC and HYC was higher (p < 0.05) compared with CON. The diversity, richness and structure of rumen microbiota showed no significant difference (p > 0.05); however, still observed remarkable increases in the relative abundance of several specific genera including Succiniclasticum and Fibrobacter with increasing doses of yeast culture. In addition, at the ASV level, ASV83, ASV123 (Succiniclasticum), and ASV148, ASV250 (Fibrobacter) were increased in YC groups. In conclusion, we confirmed that the supplementation of YC in diet could improve the growth and slaughter performance of fattening Hu sheep through improving nutrient digestion, especially nitrogen utilization, rumen microbial environment and the development of rumen epithelium, which proves the benefits of paraprobiotics in animal production.
1. Introduction
Statistics from the FAO showed that China’s mutton production reached 5.05 million tons in 2020, ranking first in the world and accounting for about 30% of the world’s total output [1]. With the intensification of the sheep industry, it is common to feed diets containing a high proportion of grains. However, with the increase of dietary concentrate ratio, the lactic acid, and volatile fatty acids (VFA) produced by massive fermentable carbohydrates in the rumen led to changes in rumen fermentation, such as pH drop and metabolic disorders, as well as metabolic diseases such as subacute rumen acidosis and nutritional diarrhea [2], which brought colossal economic losses to production. The indoor intensive feeding system enables feed additives to be extensively used for the manipulation of rumen fermentation and the improvement of animal performance. At the same time, since 1 July 2020, China has fully implemented the “anti-resistance order” (prohibiting the addition of growth-promoting additives in diets). It led to an increased number of studies exploring other feed additives as alternatives for growth promoters in sheep [3].
Paraprobiotics, defined as “inactivated microbial cells (non-viable) that confer a health benefit to the consumer”, hold the ability to improve the host condition. Yeast culture (YC) [4], one of the paraprobiotics, is a natural product produced from yeast fermentation. It contains, in addition to dead yeast cells, culture media and the products of yeast metabolism, including proteins, polysaccharides (mainly β-glucans and mannan-oligosaccharides), lipids, vitamins, peptides, amino acids, nucleotides, organic acids, and antioxidants [5]. Yeast culture supplementation in the animal diet has a range of beneficial effects, including increased structure and richness of the microbiome [6,7], feed intake [8], stabilization of rumen pH [9,10], increased feed digestion [11], enhancement of overall immunity [12,13], and consequently improved performance [14,15,16]. However, there are still different reports on the application effect of YC in mutton sheep. Malekkhahi et al. [17] found that there was no significant difference in dry matter intake, average daily gain or feed conversion ratio of lambs fed with 4 g/d of YC compared with the control treatment. Similarly, Tripathi et al. [18] showed that dietary YC supplementation had no significant effect on dry matter intake in lambs. As for the appropriate supplemental level of YC, studies in dairy cows [19,20] and beef cattle [21] showed that a high supplemental level of YC showed no dose effect. Ye et al. [22] added 0, 100, 200, and 300 mL YC per goat per day, respectively. The results showed that goats supplemented with YC (200 mL and 300 mL) had significantly lower ruminal pH and ammonia nitrogen compared with the control. The above studies indicate that there is a great difference in the studies on YC in fattening sheep, and the effect and appropriate supplemental level of YC is still unclear. Therefore, this experiment was conducted to study the effects of dietary yeast (Saccharomyces cerevisiae) culture on growth performance, carcass characteristics, nutrient digestibility, rumen fermentation and microbial communities of fattening Hu sheep under the condition of the same nutrient level, providing technical support for the application of YC in meat sheep production.
2. Materials and Methods
The experiment was conducted at a commercial farm in Jiangsu, China (latitude 32.01′ N, longitude 119.38′ E). The experimental procedure was approved by the Animal Ethics Committee of Chinese Academy of Agricultural Sciences (IFR-CAAS 20220614), and humane animal care and handling procedures were followed throughout the experiment.
2.1. Animals and Experimental Design
A total of 90 male fattening sheep (3- to 4-month-old, 38 ± 1.47 kg body weight) were used in a 97-day trial. All sheep were randomly assigned to 3 dietary treatments: CON diet (basal diet, of which the ratio of concentrate to roughage were 65:35 and 80:20 at early and later fattening stage, respectively), LYC diet (basal diet supplied with 10 and 20 g/d yeast culture at the early and late stages, respectively), and HYC diet (basal diet supplied with 20 and 40 g/d yeast culture at the early and late stages, respectively). Treatments (LYC or HYC) were sprinkled on the feed surface according to the required dosage before feeding the basal diet to each sheep throughout the trial. Each treatment had five replicates and each replicate included six sheep (Figure 1). Six sheep from each replicate were raised in one pen. The trial included a 7-day dietary transition period and a 90-day test period. Furthermore, the test period was divided into a 60-day early fattening period and a 30-day later fattening period.
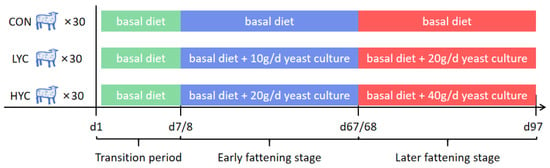
Figure 1.
The frame diagram of experimental design. CON, control (basal diet supplied with 0 g/d yeast culture at the whole stage); LYC, low dose yeast culture (basal diet supplied with 10 and 20 g/d yeast culture at the early and late stages, respectively); HYC, high dose yeast culture (basal diet supplied with 20 and 40 g/d yeast culture at the early and late stages, respectively).
2.2. YC Processing, Diets and Feeding Management
The yeast culture (YC) was a dried culture product that was produced through anaerobic fermentation of Saccharomyces cerevisiae. The inoculation of yeast was firstly on a liquid medium that contained molasses, urea, corn syrup, etc., with a set temperature of 30 °C. Then, the live bacteria cells in the liquid medium were cultured on a solid medium that was composed of corncob powder, corn flour, soybean meal, etc., with a set temperature of 30 °C. After dehydration, the YC products could be used for this experiment [23]. The ingredient analysis shows that YC contained: 18.0% crude protein, 8000 IU/g anti-microbial peptides, 300 IU/g digestive enzymes, 1.0% mannan, and 8% crude ash.
The basal diet was prepared according to the nutritional requirement of 250 g daily gain of mutton sheep in NRC (2007) [24]. The ingredients and chemical composition of the basic diet are presented in Table 1. The sheep were fed twice a day at 7:00 and 16:00 and had free access to water. Daily diet allotments to each pen were adjusted to allow minimal residuals (<5% of the total offered) in the feed trough just prior to the next morning feeding. The dietary offers and residuals were recorded daily. The sheep were weighed at 8, 67 and 97 days. The dry matter intake, average daily gain and feed efficiency were calculated accordingly. The floors, pens and feed troughs were disinfected, and all the sheep were earmarked, dewormed, and immunized according to normal immunization procedures before the trial.

Table 1.
Ingredients and chemical composition of the basal diets for the fattening sheep (%, DM basis).
2.3. Digestion and Metabolism Trial
The digestion and metabolism trials were carried out from d 15 before the end of the experiment with five sheep selected from each treatment, respectively. Sheep were moved into individual metabolism crates that could separate feces and urine. Sheep were allowed a 7-day adaptation in the metabolism crates followed by a 5-day collection period [25]. During the collection period, feed intake and refusals were recorded daily. Total feces were collected and weighed daily. After thorough and uniform mixing, 10% of total feces were collected and mixed with 10 mL 12 M HCL solution. Then, 5-day samples of each sheep were pooled together. Total urine was collected into a plastic container containing 50 mL of 12 M HCL solution to prevent ammonia loss [26]. The volume was measured and recorded daily. Then, 20% of the total urine was collected each day and pooled together after the 5-day collection period. Samples of feed, feces and urine were frozen in the refrigerator at −20 °C for further analysis. After the digestibility trial, the frozen feed, orts, and fecal subsamples were thawed overnight at room temperature, dried in an oven at 65 °C for 48 h, and then ground to pass through a 1 mm sieve. The chemical composition of the samples was analyzed according to AOAC [27] except for the neutral detergent fiber (NDF) and acid detergent fiber (ADF) whose measurement followed the method of Wang et al. [28].
The formula for calculating the apparent digestibility of nutrients in diet:
- Apparent digestibility of certain nutrients in the diet (%) = 100 × (feed intake × the content of this nutrient in the diet − defecation amount × nutrient content in feces)/(feed intake × the content of this nutrient in the diet).
- Total excreted nitrogen (g/d) = fecal nitrogen + urinary nitrogen.
- Apparent digestible nitrogen (g/d) = intake nitrogen − fecal nitrogen.
- Deposition nitrogen (g/d) = intake nitrogen − (fecal nitrogen + urinary nitrogen).
- Apparent digestibility of nitrogen (%) = 100 × apparent digestible nitrogen/intake nitrogen.
- Nitrogen utilization rate (%) = 100×deposited nitrogen/ingested nitrogen.
- The biological value of nitrogen (%) = 100 × [ingestion of nitrogen − (fecal nitrogen + urinary nitrogen)]/(ingestion of nitrogen − fecal nitrogen).
2.4. Slaughter Performance and Rumen Parameter Determination
At the end of the experiment, five fattening sheep in each group were slaughtered to determine the slaughtering performance. After slaughtering, the head, hoof and viscera were removed, and the carcass was weighed after peeling. The rumen was separated, and the contents were removed and washed. Rumen tissue sections was stored in 70% ethanol until further processing after 24 h of fixing in formalin. All samples were embedded in paraffin blocks and were stained with Yihong-haematoxylin (H.E.) at the Chinese Agriculture University (Beijing, China). The length and width of the rumen papillae and stratum corneum thickness were measured using the Axiovision software of image-pro express image analysis processing system (Zeiss, Oberkochen, Germany). The ruminal digesta sample was collected and the pH value was measured immediately (Testo 206-pH2, Germany). The rumen digesta was then filtered through four layers of cheesecloth, and a 10 mL subsample of each strained fluid was collected and stored at −20 °C for analysis of the volatile fatty acids (VFAs) and ammonia nitrogen (NH3-N) as described by Huang et al. [29] and Haro et al. [30]. The sulfuric acid paper was used to draw the outline of the superior ophthalmic muscle (longissimus dorsi muscle) of the spine between the penultimate first and second ribs, and the area of the ophthalmic muscle were measured. The tissue thickness between the 12th and 13th ribs and 11 cm away from the midline of the spine (i.e., GR value) was measured with vernier caliper.
2.5. Next-Generation Sequencing
Then, the rumen liquid for the analysis of rumen microbial communities were kept in 10 mL tubes, immediately placed in liquid nitrogen, and then stored at −80 °C. After the extraction of total genome DNA, bacterial 16S rRNA gene regions were amplified using PCR primers (341F:5′-CCTAYGGGRBGCASCAG-3′; 806R: 5′-GGACTACNNGGGTATCTAAT-3′). To prevent contamination from reagents, negative controls for DNA extraction and PCR amplification were included, and no PCR products of the negative controls were detected from agarose gel. Library quality was checked using a Thermo NanoDrop 2000 UV microphotometer and 2% agarose gel electrophoresis. A Qubit 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA) was used to quantify the library. Amplicon libraries were sequenced using an Illumina Miseq PE250 platform (Allwegene Technology Co., Ltd., Shanghai, China). The obtained raw data from the sequencer were imported into the QIIME2 platform (2021.4 released), and were processed by the Deblur program, which obtains single-nucleotide resolution based on error profiles within samples. Deblur denoised sequences are usually called amplicon sequence variants (ASVs). The number of reads from each sample was rarefied to 5556. The taxonomy of these features was assigned to the Greengenes reference database (13-8 version) classifier with 99% similarity. A feature table was generated using Qiime2′s qiime vsearch cluster-features-closed-reference command. Determination of alpha and beta diversities and analysis of similarity (ANOSIM) was also conducted in qiime2.
2.6. Statistical Analysis
The data on growth performance, nutrient utilization, organ weight and meat quality were subjected to the one-way ANOVA procedure of SAS 9.1 statistical software. If the difference was significant, Duncan’s method was used for multiple comparison tests. p < 0.05 was the judgment standard of significant difference.
Alpha diversity of the rumen microbial data between treatments was tested using the Kruskal–Wallis test and a post hoc Dunn Kruskal–Wallis multiple comparisons with a Bonferroni adjustment were used to evaluate differences between treatments with boxplots being made in R (‘ggpubr’ packages). Beta diversity was visualized with a PCoA plot. Featured bacteria among treatments were identified by the linear discriminant analysis (LDA) effect size (LEfSe) with an LDA score > 2 [31].
3. Results
3.1. Growth Performance
No differences in body weight were observed (p > 0.05) among groups (Table 2). In the early fattening stage, although the YC supplementation did not affect the DMI and ADG of Hu sheep (p > 0.05), the feed-to-gain ratio tended to be lower than that of CON (p = 0.082). In the later and whole fattening stage, ADG and feed-to-gain ratio were significantly higher (p < 0.05) in LYC compared with CON. In addition, in the later fattening stage, the DMI of LYC tended to be higher than that of CON (p = 0.076); There were no differences in DMI between groups in the early and whole fattening stage (p > 0.05).
Table 2.
Effect of dietary supplementation of YC on growth performance of fattening Hu sheep.
3.2. Nutrient Apparent Digestibility
DM, OM, CP, NDF and ADF were higher (p < 0.05) in LYC and HYC compared with CON (Table 3).
Table 3.
Effect of dietary supplementation of YC on nutrient apparent digestibility of fattening Hu sheep.
3.3. Nitrogen Metabolism
Urinary N and total N excretion of LYC were lower (p < 0.05) than CON and HYC, but there were no differences in HYC compared with CON (p > 0.05, Table 4). The Retained N, the Utilization efficiency of N and the biological value of N were higher (p < 0.05) in LYC compared with CON and HYC. While there were no differences among groups in the N intake, fecal N, apparent digestible N, and apparent digestibility of N (p > 0.05).
Table 4.
Effect of dietary different supplemental levels of YC on the nitrogen metabolism of fattening Hu sheep.
3.4. Rumen Fermentation Parameters
As shown in Table 5, NH3-N was higher (p < 0.05) in LYC and HYC compared with CON, although there were no differences in LYC and HYC (p > 0.05). There were no differences among groups in the pH, microbial protein, TVFA, acetate/TVFA, propionate/TVFA, butyrate/TVFA, and acetate/propionate (p > 0.05).
Table 5.
Effect of dietary different supplemental levels of YC on the rumen fermentation parameters of fattening Hu sheep.
3.5. Development of the Rumen Epithelium
The papillary height of the rumen was higher (p < 0.05) in LYC compared with CON and HYC, whereas rumen wall thickness and muscular layer thickness were higher (p < 0.05) in HYC compared with CON and LYC (Table 6).
Table 6.
Effect of dietary different supplemental levels of YC on morphology index of the rumen in fattening Hu sheep.
3.6. Carcass Characteristics
The dressing percentage was higher (p < 0.05) in LYC and HYC compared with CON. There were no differences in live weight before slaughter, carcass weight, eye area, and GR value among groups (p > 0.05) during dietary different supplemental levels of YC (Table 7).
Table 7.
Effect of dietary different supplemental levels of YC on carcass characteristics of fattening Hu sheep.
3.7. Rumen Microbial Communities
In this study, there were no significant differences in microbial richness (Chao1 index) and diversity (Shannon index) (Figure 2A,B) (p > 0.05). Similarly, we did not observe the distinct clusters of microbial community among groups according to the PCoA plot of Beta diversity (p > 0.05) (Figure 2C).
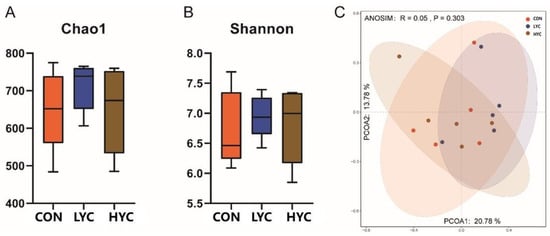
Figure 2.
Effect of yeast culture supplementation on microbial diversity and structure. CON, control (basal diet supplied with 0 g/d yeast culture at the whole stage); LYC, low dose yeast culture (basal diet supplied with 10 and 20 g/d yeast culture at the early and late stages, respectively); HYC, high dose yeast culture (basal diet supplied with 20 and 40 g/d yeast culture at the early and late stages, respectively). (A) the richness of rumen microbiota. (B) The diversity of rumen microbiota. (C) The principal coordinate analysis (PCoA) based on the bacterial Bray–Curtis in each sample.
In terms of the rumen composition, at the phylum level, Firmicutes and Bacteroidetes were the dominant phyla across the three groups (Figure 3A). At the genus level, Prevotella, Succiniclasticum, Treponema, Ruminococcus and Fibrobacter were the top 5 genera. The relative abundance of Prevotella decreased with yeast culture supplementation (33.72% vs. 30.17% vs. 29.51%), while the abundance of Succiniclasticum and Fibrobacter showed an opposite pattern (5.46% vs. 7.11% vs. 9.52%; 3.24% vs. 2.75% vs. 5.47%) (Figure 3B).
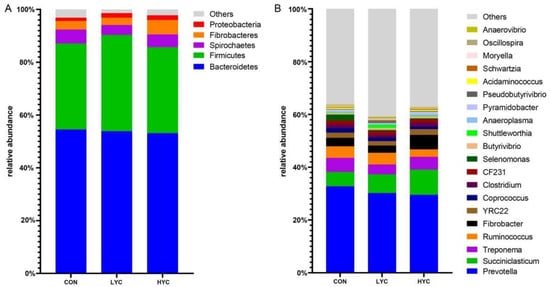
Figure 3.
The composition of rumen microbiota at phyla level (A) and genus level (B). CON, control (basal diet supplied with 0 g/d yeast culture at the whole stage); LYC, low dose yeast culture (basal diet supplied with 10 and 20 g/d yeast culture at the early and late stages, respectively); HYC, high dose yeast culture (basal diet supplied with 20 and 40 g/d yeast culture at the early and late stages, respectively).
To deeply understand the microbial composition affected by YC, linear discriminant analysis (LDA) effect size (LEfSe) analysis was performed to identify the featured microbiota at the ASVs level (Figure 4). Prevotella (ASV537) and Bacteroidales (ASV921) were identified as the featured bacteria in the CON group. Prevotella (ASV29), Shuttleworthia (ASV225), and Succiniclasticum (ASV83) were the featured bacteria in the LYC group. In the HYC group, the bacteria including S24_7 (ASV4), Succiniclasticum (ASV123), Succinogenes (ASV148, ASV250), and Prevotella (ASV1255), were higher.
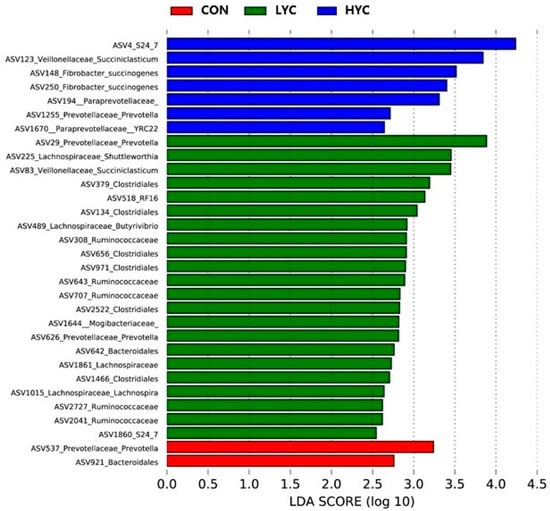
Figure 4.
The featured bacteria identified by LEfSe analyses. CON, control (basal diet supplied with 0 g/d yeast culture at the whole stage); LYC, low dose yeast culture (basal diet supplied with 10 and 20 g/d yeast culture at the early and late stages, respectively); HYC, high dose yeast culture (basal diet supplied with 20 and 40 g/d yeast culture at the early and late stages, respectively). ASVs in this figure was significant (p < 0.05) and had an LDA Score > 2, which was considered a significant effect size.
To detect the correlations between these featured bacteria and rumen phenotypes (VFAs, pH, NH3-N, rumen epithelial morphology) (Figure 5). Spearman correlation matrix was used to explore their relationships. Only Prevotella (ASV1255) and Succiniclasticum (ASV123) showed negative correlations with Acetate (p < 0.05, |r| > 0.5), while other bacteria did not construct significant correlations with VFAs. In contrast, many featured bacteria had associations with NH3-N and epithelial morphology. For instance, Prevotella (ASV29) and Shuttleworthia (ASV225) were positively correlated with NH3-N, papillae height, epithelial thickness and keratin layer thickness (p < 0.05, |r| > 0.5). Clostridiales (ASV379, ASV656, ASV1466), Ruminococcaceae (ASV308) showed positive association with NH3-N, epithelial thickness and cuticle thickness (p < 0.05, |r| > 0.5). In addition, Ruminococcaceae (ASV643) and Clostridiales (ASV1466) were negatively correlated with muscle thickness (p < 0.05, |r| > 0.5).

Figure 5.
The associations between signature bacteria and rumen phenotypes. The colors of the squares range from blue to purple to indicate negative (closer to −1) to positive (closer to 1) correlations. The degree of significance is indicated by the symbol *, 0.01 < * p < 0.05; 0.001< ** p < 0.01; *** p < 0.001.
4. Discussion
Yeast culture (YC) is a natural product produced from yeast fermentation [4]. Studies in ruminants showed that YC can improve feed digestibility, and promote feed intake and growth performance [10,15]. However, due to the different production processes of YC, the quality of products is also different. Currently, there is no unified supplemental amount in practical application. Different management modes and different dietary compositions lead to inconsistent application effects of YC in ruminants. In this experiment, although dietary YC did not improve the average daily gain and dry matter intake of early fattening stage Hu sheep, the feed-to-gain ratio was improved during the trial, which indicated the effects of YC on growth performance. Consistently, improved nutrient digestion, rumen NH3-N, and utilization efficiency of N in YC groups were observed. The development of the rumen epithelium was found in lambs supplemented with YC. Additionally, significant changes in rumen microbiota caused by YC supplementation were identified. Our results revealed that YC application in meat sheep production is important and can be an alternative to antibiotics.
The effect of YC on ruminant performance is controversial. It was found that YC had no significant effect on the average daily feed intake, average daily gain and the feed-to-gain ratio of beef cattle when 40 g/d YC was added to the diet with the ratio of concentrate to roughage of 60:40 [32]. Similarly, Bayat [33] and Kawas [34] found that dietary YC had no effect on the growth performance of dairy cows and lambs. In this experiment, similar results were observed. However, after the ratio of concentrate to roughage was adjusted from 65:35 to 80:20 in the later fattening stage, LYC trended to increase dry matter intake, increased the average daily gain, and decreased the feed to gain ratio significantly, which is consistent with Tripathi and Karim [35]. In the whole fattening period, low supplemental level of YC improved the growth performance of fattening sheep, but high supplemental level showed no significant effect, which was similar to the dose effect of high supplemental level of YC in cows [19,20] and beef cattle [21], which may be related to the β-glucan concentration in YC. It was reported that the active components of the cell wall in YC were mainly composed of β-glucan (30–34%), mannan oligosaccharides (30%), glycoproteins (22%) and chitin [36]. The content of mannan oligosaccharides in the YC used in this study was ≥1%. According to the dose estimation, the content of β-glucan in LYC and HYC was 55.6 and 111.1 mg/kg in the early fattening stage, and 111.1 and 222.2 mg/kg in the late fattening stage, respectively. Zhou [37] found that adding 75 mg/kg β-glucan promoted the growth performance of calves, while added 100 and 200 mg/kg β-glucan decreased the growth performance of calves. These results indicate that the high supplemental level of YC may lead to the high content of β-glucan, which may have adverse effects on animal health. This may be due to overfeeding YC, whose active ingredients may stimulate phagocytosis of white blood cells and increase the production of reactive oxygen species, inflammatory mediators and cytokines [38].
As a safe and environmentally friendly feed additive, YC plays a certain role in improving the apparent digestibility of nutrients in mutton sheep. Malekkhahi et al. [17] reported that the apparent digestibility of CP and NDF of fattening sheep was significantly increased by adding 4 g/d YC in the high concentrate diet. Song et al. [15] found that fiber digestibility increased with the supplementation of YC. Similarly, Lei [39] and Jiang [40] reported that YC increased the apparent digestibility of fiber in beef cattle and dairy cows. In this study, dietary YC supplementation increased the digestibility of DM, OM, CP, NDF and ADF in group LYC and HYC compared with the control group. The common finding is an increase in fiber digestibility with the supplementation of YC, which may be due to an increase in the number of cellulose-degrading bacteria or the metabolites in YC that stimulate ruminal cellulolytic bacteria [41,42,43]. Another possibility is that YC may stimulate anaerobic fungi colonizing fiber to make the fiber more accessible to fiber-degrading bacteria rather than having a direct effect on the proliferation of these bacteria [44].
Part of the nitrogen intake is mainly used for protein synthesis, and the other part is discharged with the metabolic products, which constitute the balanced metabolism of nitrogen and the dynamic balance system of protein. This study found that rumen NH3-N, the retained N, utilization efficiency of N and biological value of N were improved by YC. Similarly, Previous studies showed that YC supplementation in diet can increase the yield of rumen microbial protein, increase the available amino acids in the small intestine and promote protein deposition [45]. The increase in NH3-N concentration indicates that rumen microbes have enhanced utilization of protein, which may be one of the important reasons why YC improves the growth performance of animals. Malekkhahi [17] and Cole [46] also found that dietary YC supplementation can significantly improve retained N. The reason may be that the addition of YC reduces the circulation of microbial nitrogen in the rumen, accelerates the emptying speed of nutrients to the rear intestinal segment, increases the circulation of total nitrogen in the duodenum, improves the absorption and utilization of nitrogen-containing substances, reduces the discharge of nutrients from feces and urine [47], and promotes the absorption and deposition of nitrogen. Although YC is widely used in practical production, due to its complex composition, the nutritional regulation mechanism of YC on ruminants is not clear, which still needs to be further explored.
VFA is the product of carbohydrate degradation by rumen microorganisms in rumen and is an important energy source for ruminants. In this experiment, YC addition had no significant effect on TVFA and the proportion of acetic acid, propionic acid and butyric acid in TVFA in the rumen fluid of fattening sheep in each group. Previous studies on dairy cows [48,49], mutton sheep [17] and beef cattle [50] also obtained similar results. The addition of YC improved nutrient apparent digestibility, but rumen VFAs did not change significantly. This may be because there is a dynamic balance between rumen VFA production and absorption.
Rumen morphological development reflects the development of the animal body, and also affects the digestion and absorption of nutrients [51]. Our study found that the supplementation of yeast culture improved papillary height, rumen wall thickness, and muscular layer thickness of the rumen in fattening Hu Sheep. Lesmeister [52] found that the addition of yeast culture in a dairy calf starter at 2% slightly improves rumen development in dairy calves. β-glucan, an important component of YC, acts as a substrate for immune response. It may activate the humoral immune system to produce antibodies, which attach to the intestinal mucosa, protect intestinal villi from loss and promote villi development to a certain extent [53]. It is reported that yeast cell walls (β-glucan and α-mannose) have protective effects on the intestinal mucosa, which competitively inhibit the binding of pathogenic bacteria to villi and prevent antigen and other substances from contacting with villi, thus promoting the normal development of villi [36].
The rumen microbiota is essential for the digestion of plant fibers and contributes to the variation of production and health traits in ruminants [54,55]. In this study, the diversity, richness, and structure of rumen microbiota were not affected by YC. A previous study related to yeast culture also found that the α diversity and β diversity were not affected by the dietary treatments, which is consistent with our result [56]. This might be due to the rapid absorption of yeast culture in the rumen and the functional redundancy of rumen microbiota maintaining the steady state of ruminal microecology [57,58]. Moreover, the remarkable increases of several specific genera, including Succiniclasticum and Fibrobacter were caused by YC supplementation. Consistently, at the ASV level, Succiniclasticum (ASV83, ASV123) and the members under Fibrobacter (i.e., ASV148, ASV250) were increased in YC. These two genera are cellulolytic bacteria and short-chain fatty acid producers [59], which may explain the increased nutrient (especially for NDF and ADF) digestion in YC groups. Interestingly, the VFA in the rumen was not affected by YC, which might be due to the rapid absorption of VFA by developed rumen morphology in YC groups. Moreover, positive correlations between Succiniclasticum and NH3-N were observed in our study, which was also found in a previous study [54]. The strong correlation between microbiota and NH3-N represented that it is feasible to increase the utilization rate of rumen nitrogen by regulating the related microbiota. However, the roles of Succiniclasticum in nitrogen utilization were still unclear. A previous study assumed that Succiniclasticum restrains the nitrogen metabolism and absorption of the host. Future studies need to classify the effects of Succiniclasticum in rumen nitrogen. Taken together, we thought yeast culture supplementation increased the abundance of some nutrient decomposers without altering the rumen microbial community and these bacteria further promoted the rumen nitrogen cycle and rumen epithelium development (Figure 6).
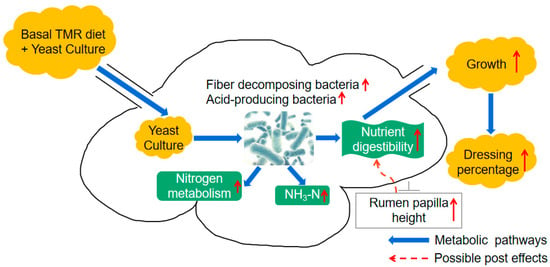
Figure 6.
Putative schematic illustration of supplementing yeast culture to promote rumen development and growth performance of fattening Hu sheep. ↑ increase.
5. Conclusions
In summary, the supplementation of yeast culture in diet could improve the growth and slaughter performance of fattening Hu sheep through improving nutrient digestion especially nitrogen utilization, rumen microbial environment and the development of rumen epithelium. Our findings provide insights that yeast (Saccharomyces cerevisiae) culture (one type of paraprobiotics) could be an alternative approach to improve ruminant performance.
Author Contributions
J.W. and G.Z. contributed to animal trial, data collection and evaluation, laboratory and statistical analysis, and writing; J.C. contributed statistical analysis and critical manuscript review; Y.Z. contributed to manuscript review and data visualization; N.Z. contributed to study design, feed formulation, and critical manuscript review. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially funded by the National Natural Science Foundation of China (31872385) and National Key R&D Program Projects (2017YFD0502001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw reads of 16S rRNA gene sequencing of ruminal microbiota are available at National Center for Biotechnology Information (NCBI, project number PRJNA897865).
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. FAOSTAT—FAO’S Online Statistical Database. Available online: http://www.fao.org/faostat/en/#home (accessed on 30 May 2022).
- Schwaiger, T.; Beauchemin, K.A.; Penner, G.B. The Duration of Time that Beef Cattle are Fed a High-Grain Diet Affects the Recovery from a Bout of Ruminal Acidosis: Dry Matter Intake and Ruminal Fermentation. J. Anim. Sci. 2013, 91, 5729–5742. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Yin, Y. Environmental Sustainability Analysis and Nutritional Strategies of Animal Production in China. Annu. Rev. Anim. Biosci. 2017, 5, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Z.; Lang, M.; Zhen, Y.G.; Chen, X.; Sun, Z.; Zhao, W.; Zhang, X.F.; Wang, T.; Qin, G.X. Effects of Yeast Culture Supplementation and the Ratio of Non-Structural Carbohydrate to Fat on Growth Performance, Carcass Traits and the Fatty Acid Profile of the Longissimus Dorsi Muscle in Lambs. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Alugongo, G.M.; Xiao, J.; Wu, Z.; Li, S.; Wang, Y.; Cao, Z. Review: Utilization of Yeast of Saccharomyces cerevisiae Origin in Artificially Raised Calves. J. Anim. Sci. Biotechnol. 2017, 8, 34. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, S.I.; Ricke, S.C. Microbial Populations in Naked Neck Chicken Ceca Raised on Pasture Flock Fed with Commercial Yeast Cell Wall Prebiotics via an Illumina MiSeq Platform. PLoS ONE 2016, 11, e0151944. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Guo, X.; Liang, Y.; Wang, W. Yeast Culture Dietary Supplementation Modulates Gut Microbiota, Growth and Biochemical Parameters of Grass Carp. Microb. Biotechnol. 2018, 11, 551–565. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Karim, S.A. Effect of Yeast Cultures Supplementation on Live Weight Change, Rumen Fermentation, Ciliate Protozoa Population, Microbial Hydrolytic Enzymes Status and Slaughtering Performance of Growing Lamb. Livest. Sci. 2011, 135, 17–25. [Google Scholar] [CrossRef]
- Li, S.; Yoon, I.; Scott, M.; Khafipour, E.; Plaizier, J.C. Impact of Saccharomyces cerevisiae Fermentation Product and Subacute Ruminal Acidosis on Production, Inflammation, and Fermentation in the Rumen and Hindgut of Dairy Cows. Anim. Feed. Sci. Technol. 2016, 211, 50–60. [Google Scholar] [CrossRef]
- Dias, A.L.G.; Freitas, J.A.; Micai, B.; Azevedo, R.A.; Greco, L.F.; Santos, J.E.P. Effects of Supplementing Yeast Culture to Diets Differing in Starch Content on Performance and Feeding Behavior of Dairy Cows. J. Dairy. Sci. 2018, 101, 186–200. [Google Scholar] [CrossRef]
- Dias, J.D.L.; Silva, R.B.; Fernandes, T.; Barbosa, E.F.; Graças, L.E.C.; Araujo, R.C.; Pereira, R.A.N.; Pereira, M.N. Yeast Culture Increased Plasma Niacin Concentration, Evaporative Heat Loss, and Feed Efficiency of Dairy Cows in a Hot Environment. J. Dairy. Sci. 2018, 101, 5924–5936. [Google Scholar] [CrossRef]
- Broadway, P.R.; Carroll, J.A.; Sanchez, N.C. Live Yeast and Yeast Cell Wall Supplements Enhance Immune Function and Performance in Food-Producing Livestock: A Review. Microorganisms 2015, 3, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.M.; Youssef, I.M.I.; Abd El-Tawab, M.M.; Bakr, H.A.; Eissa, N.A.; Hassan, M.S.; Giadinis, N.D.; Milewski, S.; Baumgartner, W.; Sobiech, P. Influence of Probiotic and Yeast Culture Supplementation on Selected Biochemical and Immunological Parameters of Growing Lambs. Pol. J. Vet. Sci. 2020, 23, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhou, M.; Tu, Y.; Zhang, N.F.; Deng, K.D.; Ma, T.; Diao, Q.Y. Effect of Oral Administration of Probiotics on Growth Performance, Apparent Nutrient Digestibility and Stress-Related Indicators in Holstein Calves. J. Anim. Physiol. Anim. Nutr. 2016, 100, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Wu, T.; You, P.; Wang, H.; Burke, J.L.; Kang, K.; Yu, W.; Wang, M.; Li, B.; He, Y.; et al. Dietary Supplementation of Yeast Culture Into Pelleted Total Mixed Rations Improves the Growth Performance of Fattening Lambs. Front. Vet. Sci. 2021, 8, 657816. [Google Scholar] [CrossRef]
- Mitchell, L.K.; Heinrichs, A.J. Feeding Various Forages and Live Yeast Culture on Weaned Dairy Calf Intake, Growth, Nutrient Digestibility, and Ruminal Fermentation. J. Dairy Sci. 2020, 103, 8880–8897. [Google Scholar] [CrossRef] [PubMed]
- Malekkhahi, M.; Tahmasbi, A.M.; Naserian, A.A.; Danesh Mesgaran, M.; Kleen, J.L.; Parand, A.A. Effects of Essential Oils, Yeast Culture and Malate on Rumen Fermentation, Blood Metabolites, Growth Performance and Nutrient Digestibility of Baluchi Lambs Fed High-Concentrate Diets. J. Anim. Physiol. Anim. Nutr. 2015, 99, 221–229. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Karim, S.A.; Chaturvedi, O.H.; Verma, D.L. Effect of Different Liquid Cultures of Live Yeast Strains on Performance, Ruminal Fermentation and Microbial Protein Synthesis in Lambs. J. Anim. Physiol. Anim. Nutr. 2008, 92, 631–639. [Google Scholar] [CrossRef]
- Arambel, M.J.; Kent, B.A. Effect of Yeast Culture on Nutrient Digestibility and Milk Yield Response in Early-to Midlactation Dairy Cows. J. Dairy. Sci. 1990, 73, 1560–1563. [Google Scholar] [CrossRef]
- Zaworski, E.M.; Shriver-Munsch, C.M.; Fadden, N.A.; Sanchez, W.K.; Yoon, I.; Bobe, G. Effects of Feeding Various Dosages of Saccharomyces cerevisiae Fermentation Product in Transition Dairy Cows. J. Dairy Sci. 2014, 97, 3081–3098. [Google Scholar] [CrossRef]
- Deters, E.L.; Stokes, R.S.; Genther-Schroeder, O.N.; Hansen, S.L. Effects of a Saccharomyces cerevisiae Fermentation Product in Receiving Diets of Newly Weaned Beef Steers. I. Growth Performance and Antioxidant Defense. J. Anim. Sci. 2018, 96, 3897–3905. [Google Scholar] [CrossRef]
- Ye, G.; Zhu, Y.; Liu, J.; Chen, X.; Huang, K. Preparation of Glycerol-Enriched Yeast Culture and Its Effect on Blood Metabolites and Ruminal Fermentation in Goats. PLoS ONE 2014, 9, e94410. [Google Scholar] [CrossRef] [PubMed]
- Ping, S.H.; Hao, Z.G. A Process Technology and Equipment for Preparing Yeast Culture by Further Fermentation Yeast. Chinese Patent ZL202010179292.5, 26 June 2020. [Google Scholar]
- NRC—National Research Council. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; National Academy of Science: Washintgton, DC, USA, 2007; 347p. [Google Scholar]
- Ma, T.; Wan, F.; Yang, D.; Deng, K.; Yang, K.; Diao, Q. Growth Performance, Nutrient Digestibility, and Slaughter Traits of Male Fattening Lambs Under Different Feeding Standards. Anim. Nutr. 2019, 5, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.M.; Diao, Q.Y.; Wang, H.; Tu, Y.; Tao, X.; Zhang, N.F. Effects of Weaning Age on Growth, Nutrient Digestibility and Metabolism, and Serum Parameters in Hu Lambs. Anim. Nutr. 2015, 1, 344–348. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 2010. [Google Scholar]
- Wang, S.; Ma, T.; Zhao, G.; Zhang, N.; Tu, Y.; Li, F.; Cui, K.; Bi, Y.; Ding, H.; Diao, Q. Effect of Age and Weaning on Growth Performance, Rumen Fermentation, and Serum Parameters in Lambs Fed Starter with Limited Ewe-Lamb Interaction. Animals 2019, 9, 825. [Google Scholar] [CrossRef]
- Huang, W.Q.; Cui, K.; Han, Y.; Chai, J.M.; Wang, S.Q.; Lv, X.K.; Diao, Q.Y.; Zhang, N.F. Long Term Effects of Artificial Rearing Before Weaning on the Growth Performance, Ruminal Microbiota and Fermentation of Fattening Lambs. J. Integr. Agric. 2022, 21, 1146–1160. [Google Scholar] [CrossRef]
- Haro, A.N.; Carro, M.D.; De Evan, T.; González, J. Influence of Feeding Sunflower Seed and Meal Protected Against Ruminal Fermentation on Ruminal Fermentation, Bacterial Composition and in Situ Degradability in Sheep. Arch. Anim. Nutr. 2020, 74, 380–396. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome. Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Geng, C.Y.; Ren, L.P.; Zhou, Z.M.; Chang, Y.; Meng, Q.X. Comparison of Active Dry Yeast (Saccharomyces cerevisiae) and Yeast Culture for Growth Performance, Carcass Traits, Meat Quality and Blood Indexes in Finishing Bulls. Anim. Sci. J. 2016, 87, 982–988. [Google Scholar] [CrossRef]
- Bayat, A.R.; Kairenius, P.; Stefański, T.; Leskinen, H.; Comtet-Marre, S.; Forano, E.; Chaucheyras-Durand, F.; Shingfield, K.J. Effect of Camelina Oil or Live Yeasts (Saccharomyces cerevisiae) on Ruminal Methane Production, Rumen Fermentation, and Milk Fatty Acid Composition in Lactating Cows Fed Grass Silage Diets. J. Dairy Sci. 2015, 98, 3166–3181. [Google Scholar] [CrossRef]
- Kawas, J.R.; García-Castillo, R.; Fimbres-Durazo, H.; Garza-Cazares, F.; Hernández-Vidal, J.F.G.; Olivares-Sáenz, E.; Lu, C.D. Effects of Sodium Bicarbonate and Yeast on Nutrient Intake, Digestibility, and Ruminal Fermentation of Light-Weight Lambs Fed Finishing Diets. Small. Rumin. Res. 2007, 67, 149–156. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Karim, S.A. Effect of Individual and Mixed Live Yeast Culture Feeding on Growth Performance, Nutrient Utilization and Microbial Crude Protein Synthesis in Lambs. Anim. Feed. Sci. Technol. 2010, 155, 163–171. [Google Scholar] [CrossRef]
- Kogan, G.; Kocher, A. Role of Yeast Cell Wall Polysaccharides in Pig Nutrition and Health Protection. Livest. Sci. 2007, 109, 161–165. [Google Scholar] [CrossRef]
- Zhou, Y.; Diao, Q.Y.; Tu, Y.; Yun, Q. Effects of Yeast β-glucan and Bacitracin Zinc on Growth Performance and Gastrointestinal Development of Early-Weaned Calves. Chin. J. Anim. Nutr. 2011, 23, 813–820. [Google Scholar]
- Li, Y.; Shen, Y.; Niu, J.; Guo, Y.; Pauline, M.; Zhao, X.; Li, Q.; Cao, Y.; Bi, C.; Zhang, X.; et al. Effect of Active Dry Yeast on Lactation Performance, Methane Production, and Ruminal Fermentation Patterns in Early-Lactating Holstein Cows. J. Dairy Sci. 2021, 104, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.L.; Dong, G.Z.; Jin, L.; Zhang, S.; Zhou, J. Effects of Dietary Supplementation of Montmorillonite and Yeast Cell Wall on Lipopolysaccharide Adsorption, Nutrient Digestibility and Growth Performance in Beef Cattle. Livest. Sci. 2013, 158, 57–63. [Google Scholar] [CrossRef]
- Jiang, Y.; Ogunade, I.M.; Arriola, K.G.; Qi, M.; Vyas, D.; Staples, C.R.; Adesogan, A.T. Effects of the Dose and Viability of Saccharomyces cerevisiae. 2. Ruminal Fermentation, Performance of Lactating Dairy Cows, and Correlations Between Ruminal Bacteria Abundance and Performance Measures. J. Dairy Sci. 2017, 100, 8102–8118. [Google Scholar] [CrossRef]
- Knollinger, S.E.; Poczynek, M.; Miller, B.; Mueller, I.; de Almeida, R.; Murphy, M.R.; Cardoso, F.C. Effects of Autolyzed Yeast Supplementation in a High-Starch Diet on Rumen Health, Apparent Digestibility, and Production Variables of Lactating Holstein Cows. Animals 2022, 12, 2445. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, X.; Wanapat, M.; Shah, A.M.; Luo, X.; Peng, Q.; Kang, K.; Hu, R.; Guan, J.; Wang, Z. Ruminal pH Pattern, Fermentation Characteristics and Related Bacteria in Response to Dietary Live Yeast (Saccharomyces cerevisiae) Supplementation in Beef Cattle. Anim. Biosci. 2022, 35, 184–195. [Google Scholar] [CrossRef]
- Zhou, X.; Ouyang, Z.; Zhang, X.; Wei, Y.; Tang, S.; Ma, Z.; Tan, Z.; Zhu, N.; Teklebrhan, T.; Han, X. Sweet Corn Stalk Treated with Saccharomyces cerevisiae Alone or in Combination with Lactobacillus Plantarum: Nutritional Composition, Fermentation Traits and Aerobic Stability. Animals 2019, 9, 598. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Rodriguez-Herrera, M.V.; Marroquin-Cardona, A.G.; Kawas, J.R. The Health Enhancer Yeast Saccharomyces cerevisiae in Two Types of Commercial Products for Animal Nutrition. Lett. Appl. Microbiol. 2019, 68, 472–478. [Google Scholar] [CrossRef]
- Hristov, A.N.; Varga, G.; Cassidy, T.; Long, M.; Heyler, K.; Karnati, S.K.; Corl, B.; Hovde, C.J.; Yoon, I. Effect of Saccharomyces cerevisiae Fermentation Product on Ruminal Fermentation and Nutrient Utilization in Dairy Cows. J. Dairy Sci. 2010, 93, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Cole, N.A.; Purdy, C.W.; Hutcheson, D.P. Influence of Yeast Culture on Feeder Calves and Lambs. J. Anim. Sci. 1992, 70, 1682–1690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, J.; Yi, X.; Li, Z.; Liang, S.; Fang, Z.; Lin, Y.; Xu, S.; Feng, B.; Zhuo, Y.; et al. Rhodotorula Benthica Culture as an Alternative to Antibiotics Improves Growth Performance by Improving Nutrients Digestibility and Intestinal Morphology, and Modulating Gut Microbiota of Weaned Piglets. Front. Microbiol. 2022, 13, 964531. [Google Scholar] [CrossRef]
- Dias, A.L.G.; Freitas, J.A.; Micai, B.; Azevedo, R.A.; Greco, L.F.; Santos, J.E.P. Effect of Supplemental Yeast Culture and Dietary Starch Content on Rumen Fermentation and Digestion in Dairy Cows. J. Dairy Sci. 2018, 101, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Aikman, P.C.; Henning, P.H.; Humphries, D.J.; Horn, C.H. Rumen pH and Fermentation Characteristics in Dairy Cows Supplemented with Megasphaera Elsdenii NCIMB 41125 in Early Lactation. J. Dairy Sci. 2011, 94, 2840–2849. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, H.; Ran, T.; Yoon, I.; Saleem, A.M.; Yang, W. Influence of Yeast Culture and Feed Antibiotics on Ruminal Fermentation and Site and Extent of Digestion in Beef Heifers Fed High Grain Rations1. J. Anim. Sci. 2018, 96, 3916–3927. [Google Scholar] [CrossRef]
- Wang, J.; Cui, K.; Ma, T.; Zhang, F.; Wang, S.Q.; Tu, Y.; Diao, Q.Y.; Zhang, N.F. Effects of Dietary Methionine Deficiency Followed by Replenishment on the Growth Performance and Carcass Characteristics of Lambs. Anim. Prod. Sci. 2018, 59, 243–248. [Google Scholar] [CrossRef]
- Lesmeister, K.E.; Heinrichs, A.J.; Gabler, M.T. Effects of Supplemental Yeast (Saccharomyces cerevisiae) Culture on Rumen Development, Growth Characteristics, and Blood Parameters in Neonatal Dairy Calves. J. Dairy Sci. 2004, 87, 1832–1839. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, Y.; Wang, F.; He, H.; Wan, K.; Liu, A. Effect of Yeast Culture Supplementation on Blood Characteristics, Body Development, Intestinal Morphology, and Enzyme Activities in Geese. J. Anim. Physiol. Anim. Nutr. 2022, 31, 1–9. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, H.; Li, C. Response of Sheep Rumen Fermentation and Microbial Communities to Feed Infected with the Endophyte Epichloë Gansuensis as Evaluated with Rumen-Simulating Technology. J. Microbiol. 2021, 59, 718–728. [Google Scholar] [CrossRef]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host Genetics Influence the Rumen Microbiota and Heritable Rumen Microbial Features Associate with Feed Efficiency in Cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ogunade, I.M.; Pech-Cervantes, A.A.; Fan, P.X.; Li, X.; Kim, D.H.; Arriola, K.G.; Poindexter, M.B.; Jeong, K.C.; Vyas, D.; et al. Effect of Sequestering Agents Based on a Saccharomyces cerevisiae Fermentation Product and Clay on the Ruminal Bacterial Community of Lactating Dairy Cows Challenged with Dietary Aflatoxin B1. J. Dairy Sci. 2020, 103, 1431–1447. [Google Scholar] [CrossRef]
- Moraïs, S.; Mizrahi, I. The Road Not Taken: The Rumen Microbiome, Functional Groups, and Community States. Trends Microbiol. 2019, 27, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, I.; Wallace, R.J.; Moraïs, S. The Rumen Microbiome: Balancing Food Security and Environmental Impacts. Nat. Rev. Microbiol. 2021, 19, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Guo, W.; Zhao, N.; Chen, F.; Fan, Q.; Zhang, W.; Huang, S.; Zhou, G.; Fu, T.; Wei, J. Effects of Diets with Various Levels of Forage Rape (Brassica Napus) on Growth Performance, Carcass Traits, Meat Quality and Rumen Microbiota of Hu Lambs. J. Sci. Food. Agric. 2022, 102, 1281–1291. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).