
Hydrogen-Rich Water Can Restrict the Formation of Biogenic Amines in Red Beet Pickles


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Red Beet Pickle Preparation
2.2. pH and Oxidoreduction Potential (Eh) Measurement
2.3. Microbiological Counts
2.4. Biogenic Amine Analysis
2.5. Statistical Analysis
3. Results
3.1. pH and Oxidoreduction Potential (Eh)
3.1.1. pH Measurement
3.1.2. Eh Measurement
3.2. Microbiological Counts
3.3. Biogenic Amines
3.3.1. Aromatic Amines
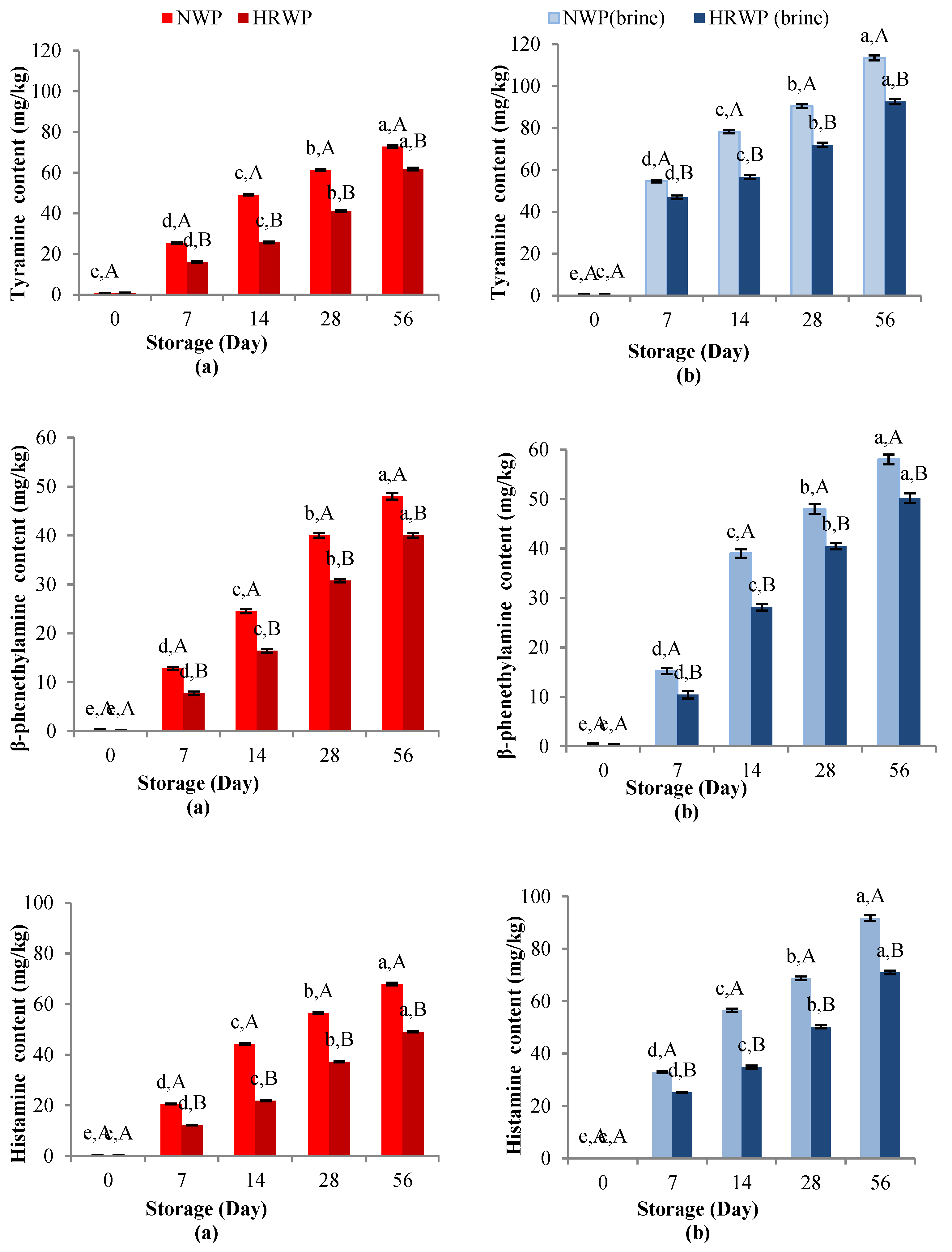
Tyramine
Phenylethylamine
3.3.2. Heterocyclic Amines
Histamine
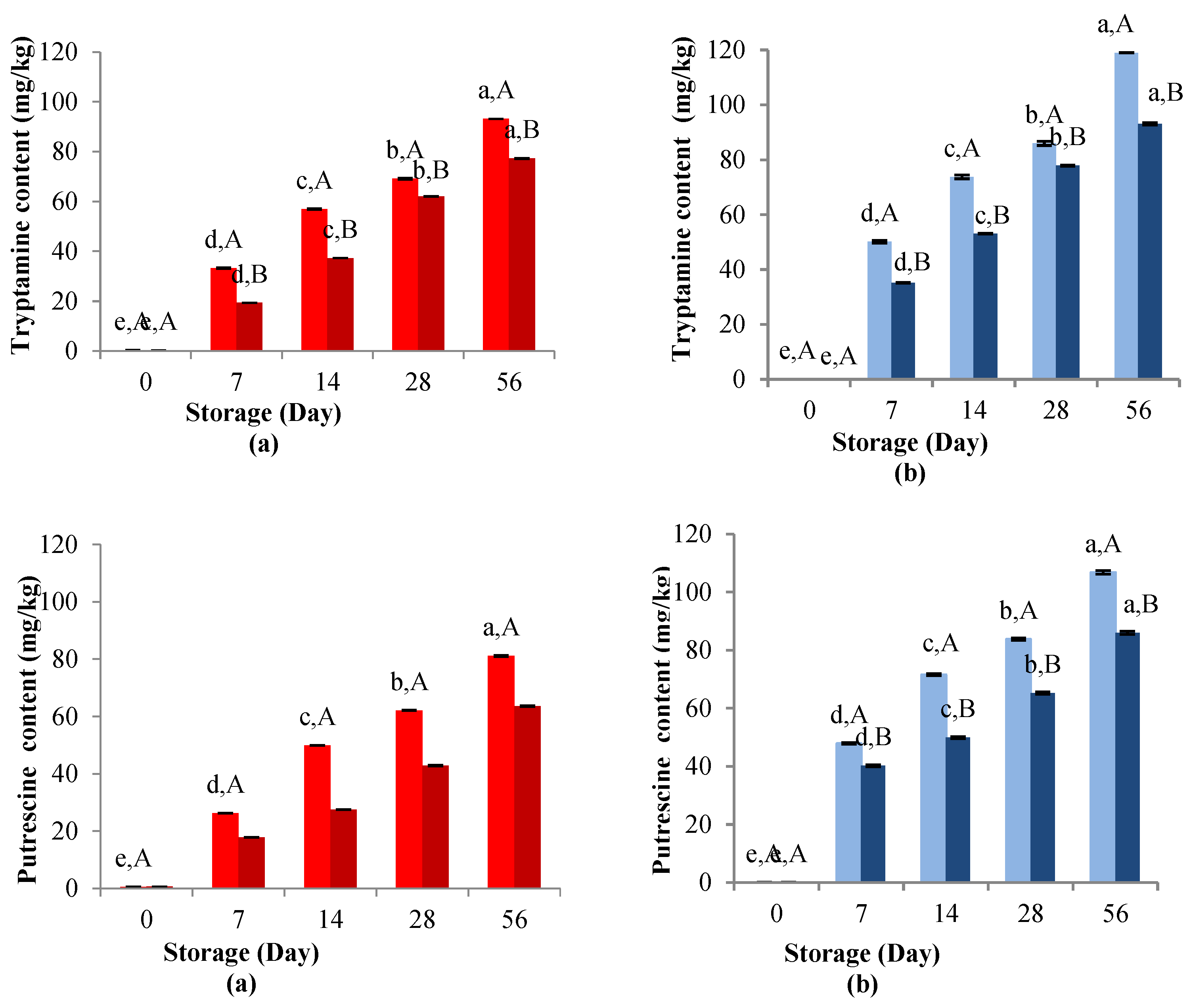
Tryptamine
3.3.3. Aliphatic Di-Amines
Putrescine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baráth, Á.; Halász, A.; Németh, E.; Zalán, Z. Selection of LAB strains for fermented red beet juice production. Eur. Food Res. Technol. 2004, 218, 184–187. [Google Scholar] [CrossRef]
- Akan, S.; Tuna Gunes, N.; Erkan, M. Red beetroot: Health benefits, production techniques, and quality maintaining for food industry. J. Food Process. Preserv. 2021, 45, e15781. [Google Scholar] [CrossRef]
- Sarkadi, L.S. Biogenic amines in fermented foods and health implications. In Fermented Foods in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2017; pp. 625–651. [Google Scholar]
- Moret, S.; Smela, D.; Populin, T.; Conte, L.S. A survey on free biogenic amine content of fresh and preserved vegetables. Food Chem. 2005, 89, 355–361. [Google Scholar] [CrossRef]
- Pessione, A.; Lamberti, C.; Pessione, E. Proteomics as a tool for studying energy metabolism in lactic acid bacteria. Mol. BioSyst. 2010, 6, 1419–1430. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mah, J.-H.; Park, Y.K.; Jin, Y.H.; Lee, J.-H.; Hwang, H.-J. Bacterial production and control of biogenic amines in Asian fermented soybean foods. Foods 2019, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Alwazeer, D. Reducing Atmosphere Packaging Technique for Extending the Shelf-life of Food Products. J. Inst. Sci. Technol. 2019, 9, 2117–2123. [Google Scholar] [CrossRef] [Green Version]
- Alwazeer, D.; Liu, F.F.-C.; Wu, X.Y.; LeBaron, W.T. Combating oxidative stress and inflammation in COVID−19 by molecular hydrogen therapy: Mechanisms and perspectives. Oxidative Med. Cell. Longev. 2021, 2021, 5513868. [Google Scholar] [CrossRef]
- Jafta, N.; Magagula, S.; Lebelo, K.; Nkokha, D.; Mochane, M.J. The Production and Role of Hydrogen-Rich Water in Medical Applications. In Applied Water Science Volume 1: Fundamentals and Applications; Scrivener Publishing LLC: Beverly, MA, USA, 2021; pp. 273–298. [Google Scholar] [CrossRef]
- Hu, H.; Li, P.; Wang, Y.; Gu, R. Hydrogen-rich water delays postharvest ripening and senescence of kiwifruit. Food Chem. 2014, 156, 100–109. [Google Scholar] [CrossRef]
- Alwazeer, D.; Örs, B. Reducing atmosphere drying as a novel drying technique for preserving the sensorial and nutritional notes of foods. J. Food Sci. Technol. 2019, 56, 3790–3800. [Google Scholar] [CrossRef]
- Alwazeer, D.; Örs, B.; Tan, K. Reducing atmosphere packaging as a novel alternative technique for extending shelf life of fresh cheese. J. Food Sci. Technol. 2020, 57, 3013–3023. [Google Scholar] [CrossRef] [PubMed]
- Alwazeer, D.; Özkan, N. Incorporation of hydrogen into the packaging atmosphere protects the nutritional, textural and sensorial freshness notes of strawberries and extends shelf life. J. Food Sci. Technol. 2022, 59, 3951–3964. [Google Scholar] [CrossRef] [PubMed]
- Giroux, H.J.; Acteau, G.; Sabik, H.; Britten, M. Influence of dissolved gases and heat treatments on the oxidative degradation of polyunsaturated fatty acids enriched dairy beverage. J. Agric. Food Chem. 2008, 56, 5710–5716. [Google Scholar] [CrossRef]
- Ceylan, M.M.; Bulut, M.; Alwazeer, D.; Koyuncu, M. Evaluation of the Impact of Hydrogen-Rich Water on the Quality Attribute Notes of Butter. J. Dairy Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Alwazeer, D.; Ceylan, M.M.; Bulut, M.; Koyuncu, M. Evaluation of the impact of hydrogen-rich water on the deaccumulation of heavy metals in butter. J. Food Saf. 2022, 42, e13005. [Google Scholar] [CrossRef]
- Bulut, M.; Çelebi Sezer, Y.; Ceylan, M.M.; Alwazeer, D. Hydrogen-rich water can reduce the formation of biogenic amines in butter. Food Chem. 2022, 384, 132613. [Google Scholar] [CrossRef]
- Sezer, Y.Ç.; Bulut, M.; Boran, G.; Alwazeer, D. The effects of hydrogen incorporation in modified atmosphere packaging on the formation of biogenic amines in cold stored rainbow trout and horse mackerel. J. Food Compos. Anal. 2022, 112, 104688. [Google Scholar] [CrossRef]
- Olsen, M.J. Redox Potential Trends of Cucumber Fermentation as Influenced by Microbial Growth and Gas Purging; North Carolina State University: Raleigh, NC, USA, 2008. [Google Scholar]
- Olsen, M.J.; Pérez-Díaz, I.M. Influence of microbial growth on the redox potential of fermented cucumbers. J. Food Sci. 2009, 74, M149–M153. [Google Scholar] [CrossRef]
- Alwazeer, D. Importance of consideration of oxidoreduction potential as a critical quality parameter in food industries. Food Res. Int. 2020, 132, 109108. [Google Scholar] [CrossRef]
- Pacl, H.T.; Reddy, V.P.; Saini, V.; Chinta, K.C.; Steyn, A.J.C. Host-pathogen redox dynamics modulate Mycobacterium tuberculosis pathogenesis. Pathog. Dis. 2018, 76, fty036. [Google Scholar] [CrossRef]
- Wu, C.; Zheng, J.; Huang, J.; Zhou, R. Reduced nitrite and biogenic amine concentrations and improved flavor components of Chinese sauerkraut via co-culture of Lactobacillus plantarum and Zygosaccharomyces rouxii. Ann. Microbiol. 2014, 64, 847–857. [Google Scholar] [CrossRef]
- Kieronczyk, A.; Cachon, R.; Feron, G.; Yvon, M. Addition of oxidizing or reducing agents to the reaction medium influences amino acid conversion to aroma compounds by Lactococcus lactis. J. Appl. Microbiol. 2006, 101, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, D.; Bosscher, J.S.; ten Brink, B.; Driessen, A.J.; Konings, W.N. Generation of a proton motive force by histidine decarboxylation and electrogenic histidine/histamine antiport in Lactobacillus buchneri. J. Bacteriol. 1993, 175, 2864–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagong, H.-Y.; Kim, K.-J. Lysine decarboxylase with an enhanced affinity for pyridoxal 5-phosphate by disulfide bond-mediated spatial reconstitution. PLoS ONE 2017, 12, e0170163. [Google Scholar] [CrossRef] [Green Version]
- Bjornsdottir-Butler, K.; Bencsath, F.A.; McCarthy, S.; Benner, R.A., Jr. Heat resistance of histidine decarboxylase from gram-negative histamine-producing bacteria in seafood. J. Food Prot. 2017, 80, 1273–1279. [Google Scholar] [CrossRef]
- Capuani, A.; Werner, S.; Behr, J.; Vogel, R.F. Effect of controlled extracellular oxidation–reduction potential on microbial metabolism and proteolysis in buckwheat sourdough. Eur. Food Res. Technol. 2014, 238, 425–434. [Google Scholar] [CrossRef]
- Wu, C.Y.; Hsu, W.L.; Tsai, M.H.; Liang, J.L.; Lu, J.H.; Yen, C.J.; Yu, H.S.; Noda, M.; Lu, C.Y.; Chen, C.H.; et al. Hydrogen gas protects IP3Rs by reducing disulfide bridges in human keratinocytes under oxidative stress. Sci. Rep. 2017, 7, 3606. [Google Scholar] [CrossRef]
- Dong, W.; Hoseney, R.C. Effects of certain breadmaking oxidants and reducing agents on dough rheological properties. Cereal Chem. 1995, 72, 58–63. [Google Scholar]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.I.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688. [Google Scholar] [CrossRef]
- Keten, S.; Chou, C.-C.; van Duin, A.C.T.; Buehler, M.J. Tunable nanomechanics of protein disulfide bonds in redox microenvironments. J. Mech. Behav. Biomed. Mater. 2012, 5, 32–40. [Google Scholar] [CrossRef]
- Lin, Z.; Wu, Z.; Zhang, W. Bioinformatics analysis of amino acid decarboxylases related to four major biogenic amines in pickles. Food Chem. 2022, 393, 133339. [Google Scholar] [CrossRef] [PubMed]
- Bover Cid, S.; Miguélez-Arrizado, M.J.; Becker, B.; Holzapfel, W.H.; Vidal-Carou, M.C. Amino acid decarboxylation by Lactobacillus curvatus CTC273 affected by the pH and glucose availability. Food Microbiol. 2008, 25, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Buňková, L.; Buňka, F.; Pollaková, E.; Podešvová, T.; Dráb, V. The effect of lactose, NaCl and an aero/anaerobic environment on the tyrosine decarboxylase activity of Lactococcus lactis subsp. cremoris and Lactococcus lactis subsp. lactis. Int. J. Food Microbiol. 2011, 147, 112–119. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| 0th Day | 7th Day | 14th Day | 28th Day | 56th Day | ||
|---|---|---|---|---|---|---|
| Eh7 (mV) | NWP | −18.48 ± 2.17 eA | −7.87 ± 2.20 dA | +74.10 ± 4.42 cA | +143.25 ± 5.95 aA | +132.02 ± 7.19 bA |
| HRWP | −198.95 ± 9.90 eB | −57.28 ± 4.57 bB | +70.84 ± 2.83 cA | +121.15 ± 3.43 aA | +82.43 ± 6.44 bB | |
| pH | NWP | 3.37 ± 0.06 cB | 3.07 ± 0.03 dA | 3.91 ± 0.02 aA | 3.75 ± 0.02 bB | 3.78 ± 0.02 bA |
| HRWP | 3.95 ± 0.03 aA | 3.08 ± 0.02 dA | 3.76 ± 0.01 cB | 3.85 ± 0.02 bA | 3.77 ± 0.01 cA |
| 0th Day | 7th Day | 14th Day | 28th Day | 56th Day | ||
|---|---|---|---|---|---|---|
| TMAB | NWP | 4.86 ± 0.03 eA | 8.28 ± 0.04 cB | 10.05 ± 0.07 aB | 8.94 ± 0.03 bB | 8.11 ± 0.01 dB |
| HRWP | 4.63 ± 0.04 eB | 8.47 ± 0.04 dA | 11.19 ± 0.01 aA | 9.18 ± 0.03 cA | 9.90 ± 0.01 bA | |
| Yeast–Mold | NWP | 4.91 ± 0.01 eA | 8.44 ± 0.01 cB | 8.98 ± 0.03 bB | 9.20 ± 0.03 aA | 8.36 ± 0.03 dB |
| HRWP | 4.38 ± 0.04 eB | 9.75 ± 0.06 cA | 10.00 ± 0.06 bA | 9.36 ± 0.06 dA | 10.70 ± 0.07 aA | |
| LAB | NWP | 0.00 ± 0.00 eA | 8.62 ± 0.06 cB | 9.78 ± 0.04 aB | 9.17 ± 0.08 bA | 8.03 ± 0.18 dB |
| HRWP | 0.00 ± 0.00 dA | 10.01 ± 0.01 bA | 11.00 ± 0.14 aA | 9.30 ± 0.42 cA | 9.00 ± 0.14 cA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alwazeer, D.; Bulut, M.; Çelebi, Y. Hydrogen-Rich Water Can Restrict the Formation of Biogenic Amines in Red Beet Pickles. Fermentation 2022, 8, 741. https://doi.org/10.3390/fermentation8120741
Alwazeer D, Bulut M, Çelebi Y. Hydrogen-Rich Water Can Restrict the Formation of Biogenic Amines in Red Beet Pickles. Fermentation. 2022; 8(12):741. https://doi.org/10.3390/fermentation8120741
Chicago/Turabian StyleAlwazeer, Duried, Menekşe Bulut, and Yasemin Çelebi. 2022. "Hydrogen-Rich Water Can Restrict the Formation of Biogenic Amines in Red Beet Pickles" Fermentation 8, no. 12: 741. https://doi.org/10.3390/fermentation8120741