
Regulation of β-Disaccharide Accumulation by β-Glucosidase Inhibitors to Enhance Cellulase Production in Trichoderma reesei


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Microorganisms
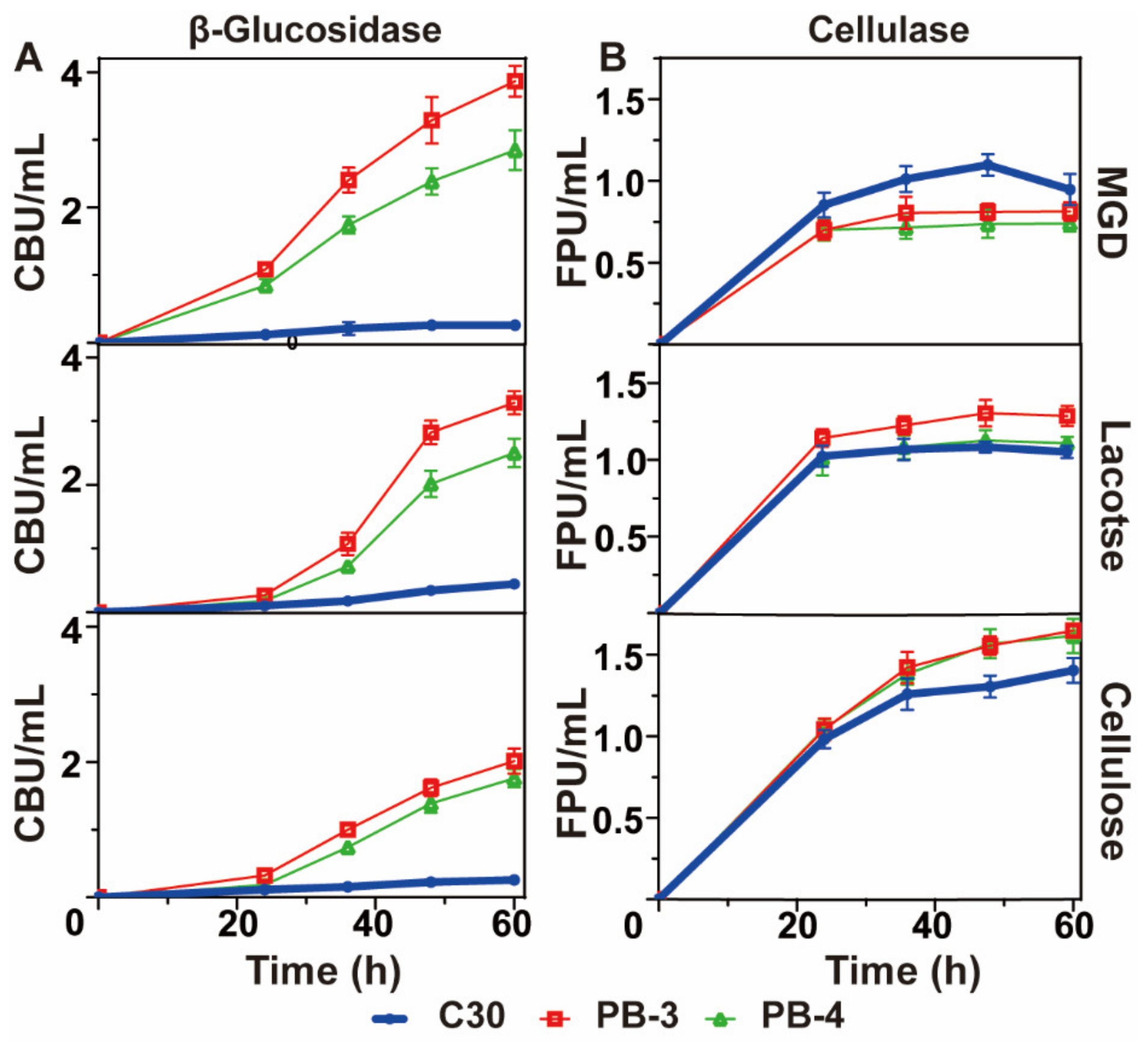
2.2. Production of Cellulase and β-Glucosidase from T. reesei C30, PB3 and PB4
2.3. Development and Validation of β-Glucosidase Inhibitors
2.4. Molecular Docking
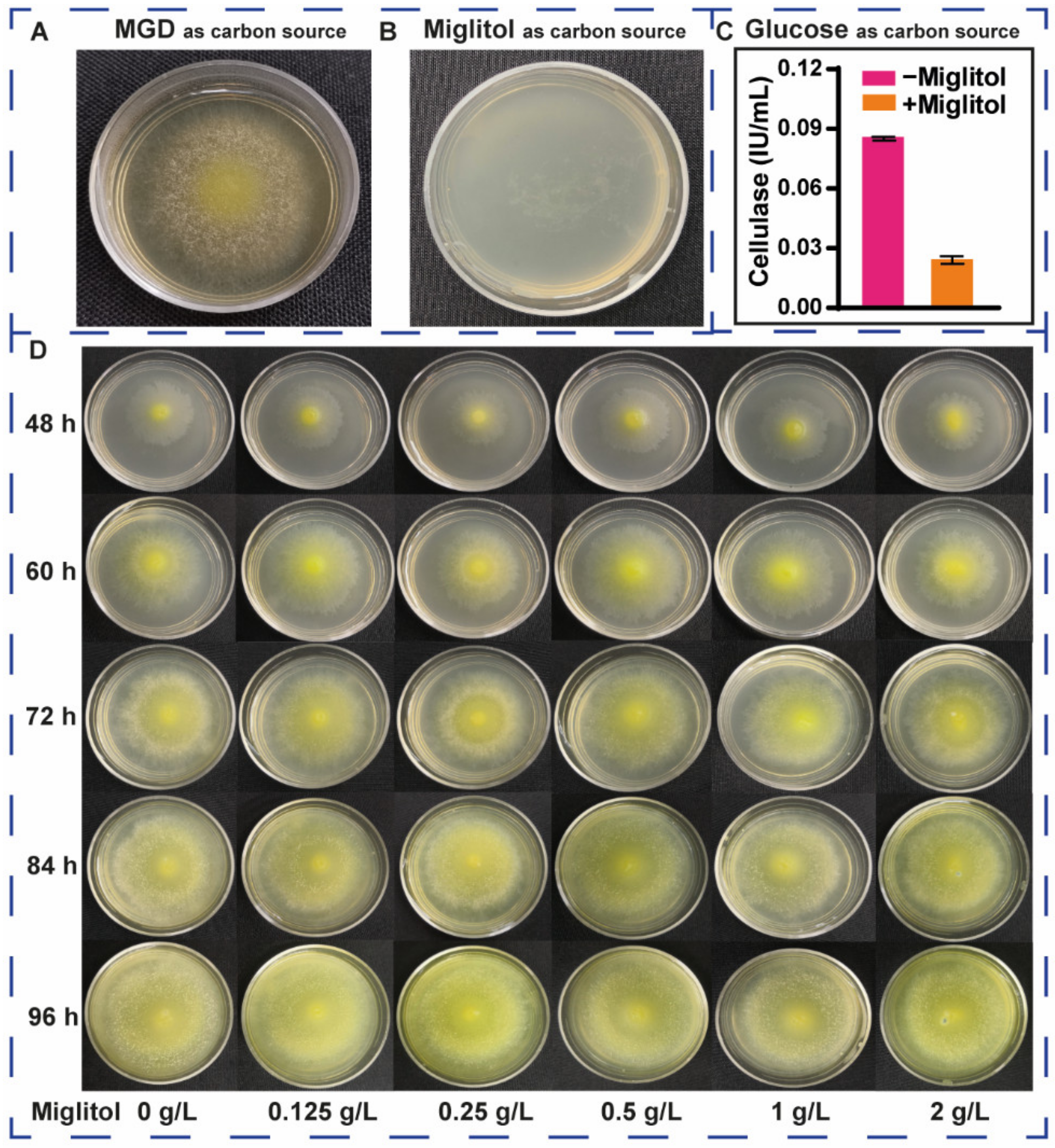
2.5. Effects of Miglitol on the Growth and Cellulase Induction of T. reesei
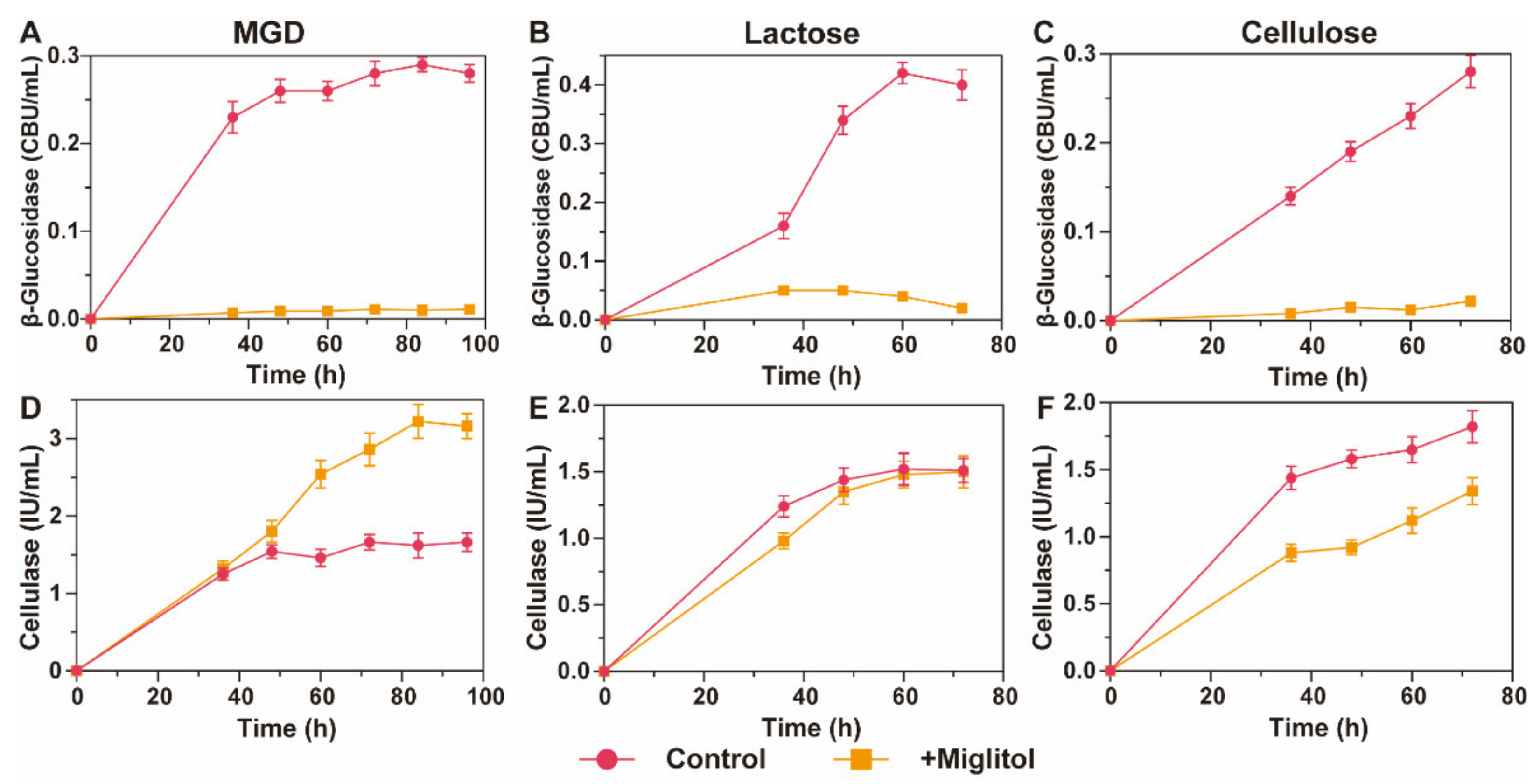
2.6. Enzyme Production Induced by Different Inducers with/without Miglitol Supplemented
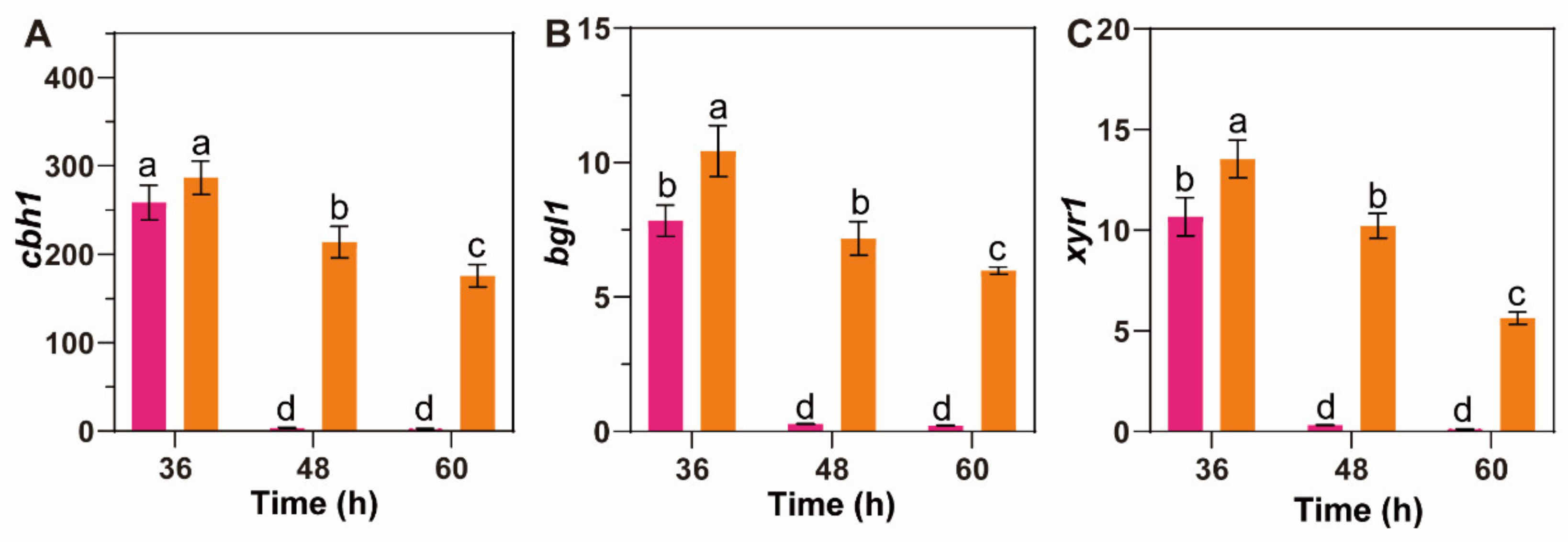
2.7. qPCR
2.8. Protein Analysis
2.9. Analysis Methods
3. Results and Discussion
3.1. Effect of Different Inducers on the Synthesis of Cellulase from T. reesei Rut C30, PB3 and PB4
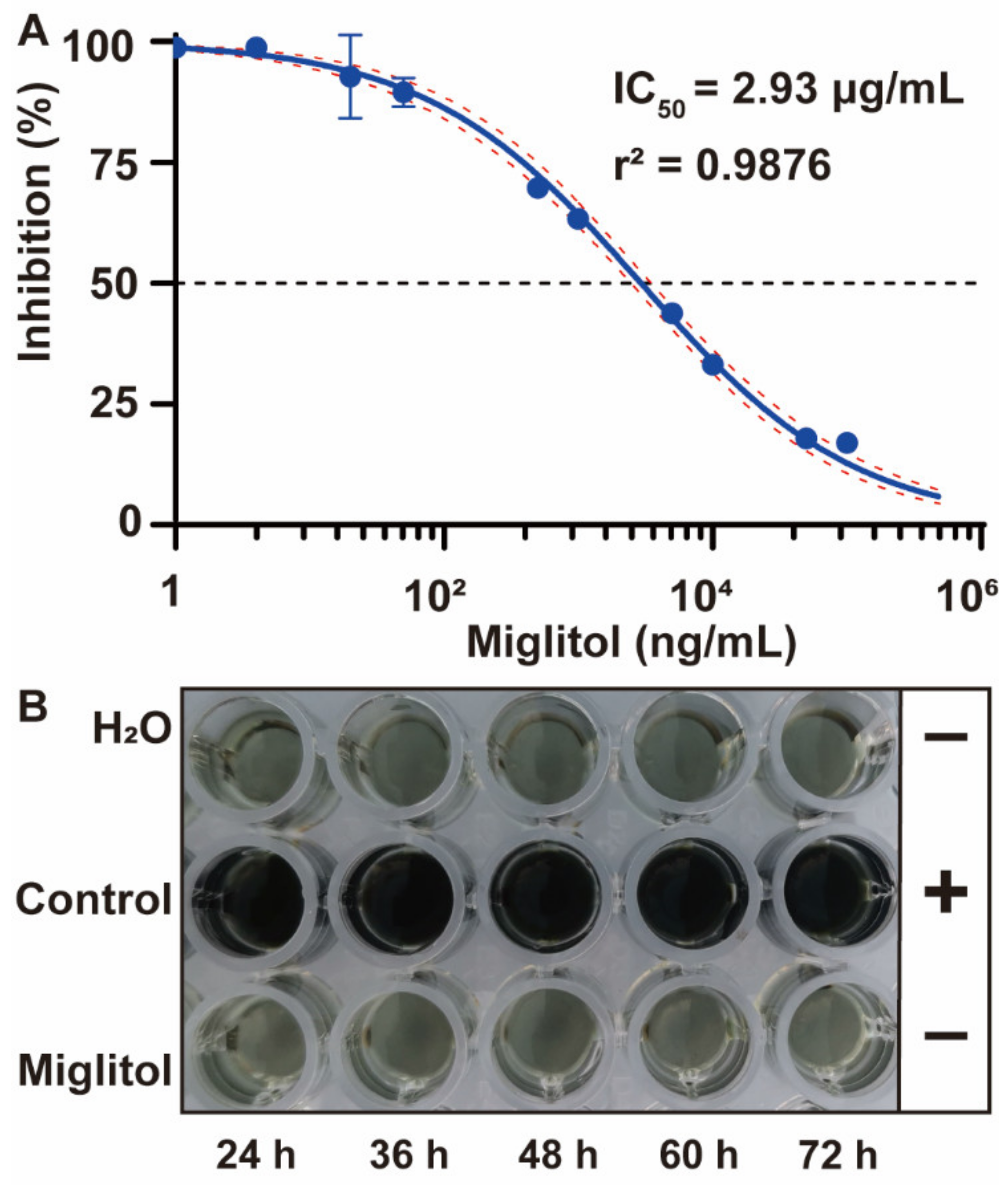
3.2. Development and Validation of β-Glucosidase Inhibitors
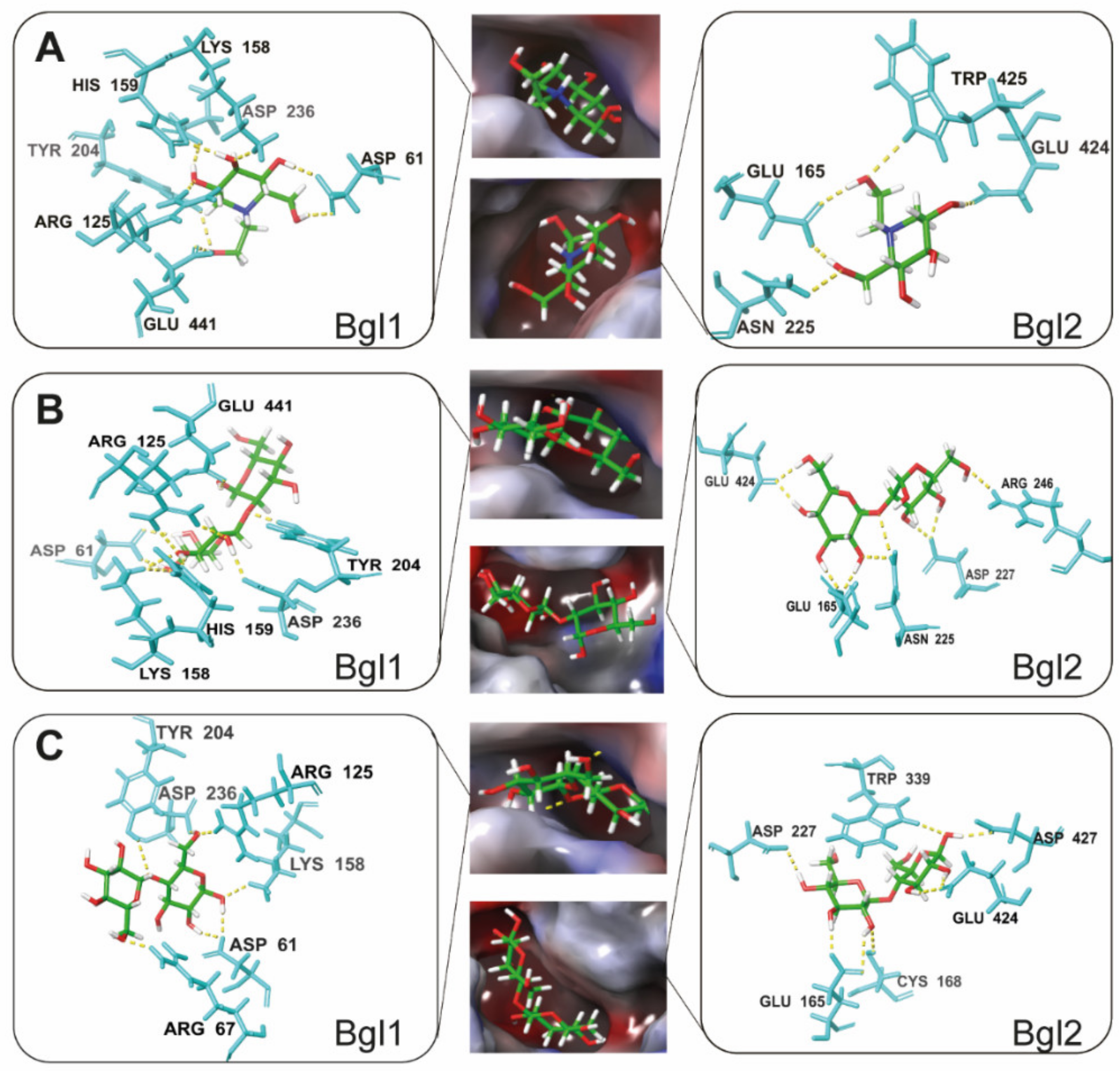
3.3. Molecular Mechanism by which Miglitol Inhibits β-Glucosidase
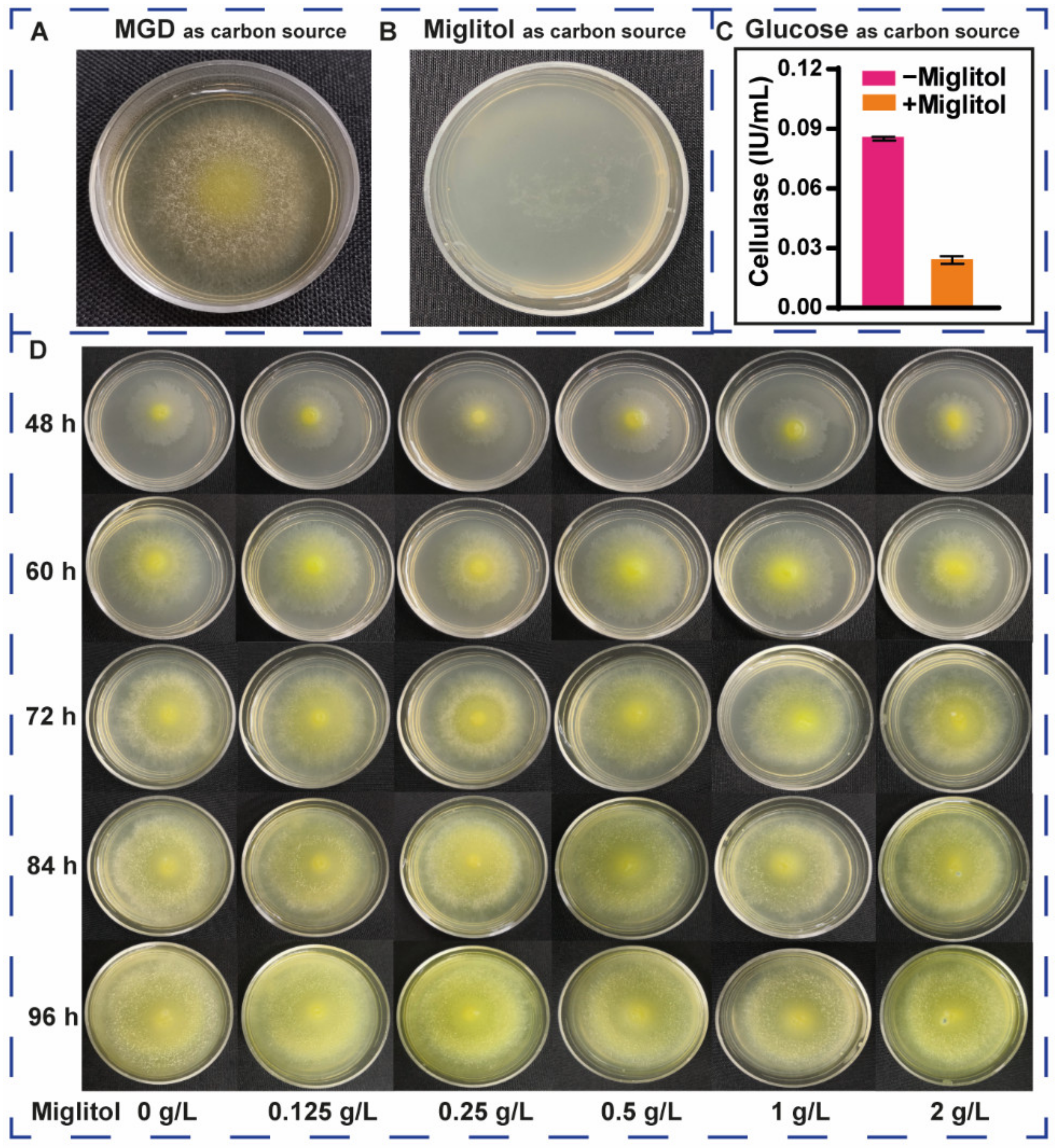
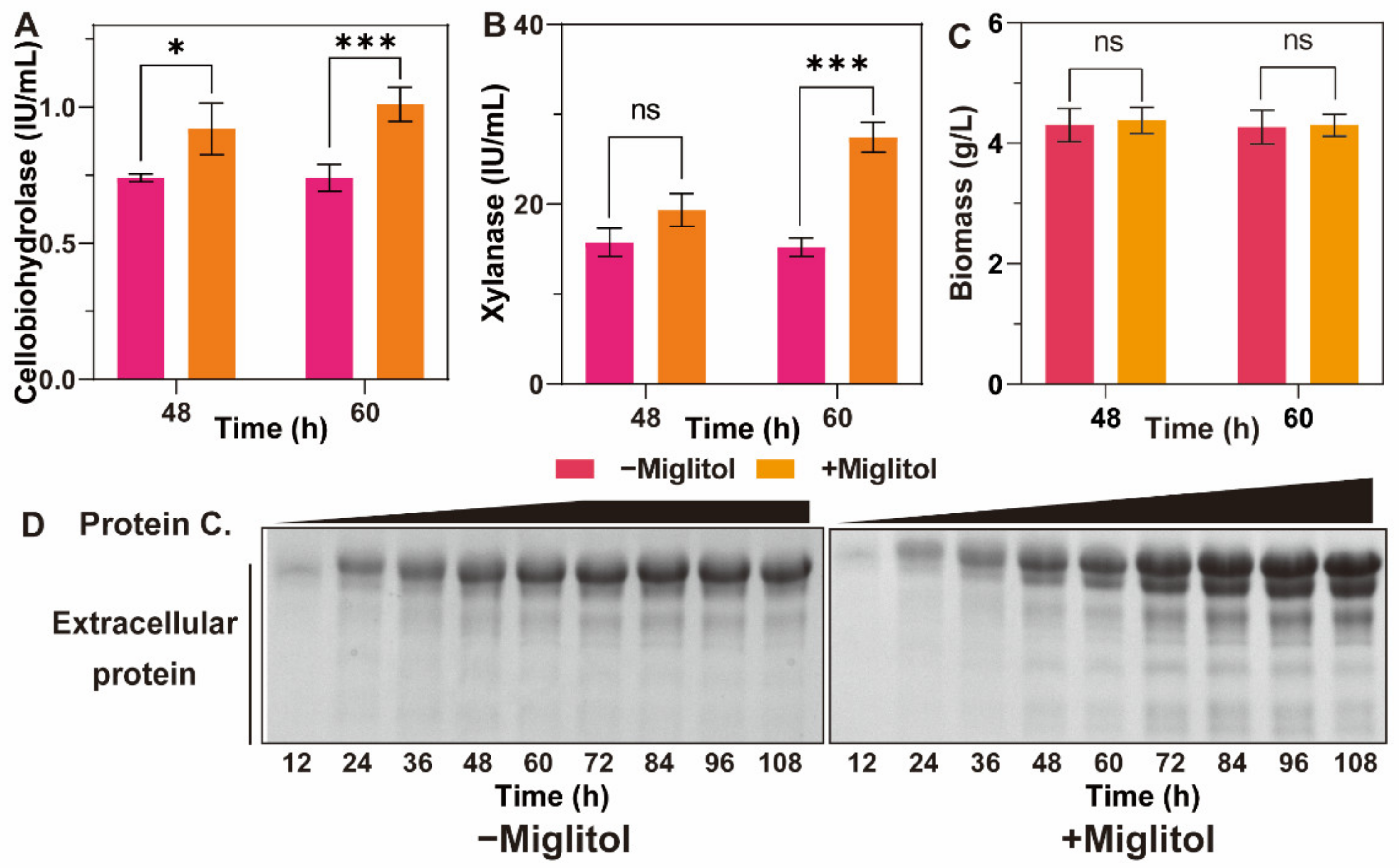
3.4. Effects of Miglitol on the Growth and Induction of T. reesei
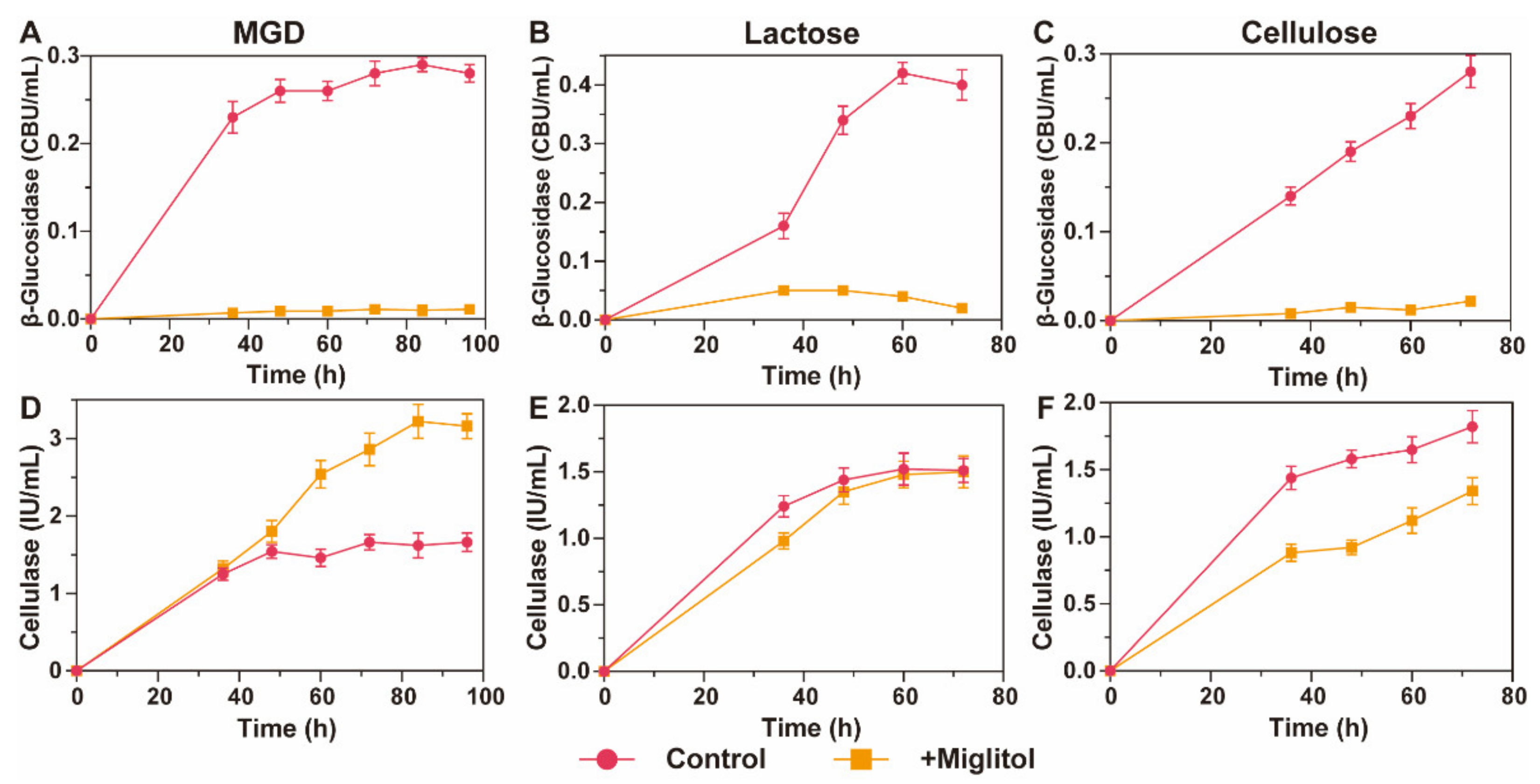
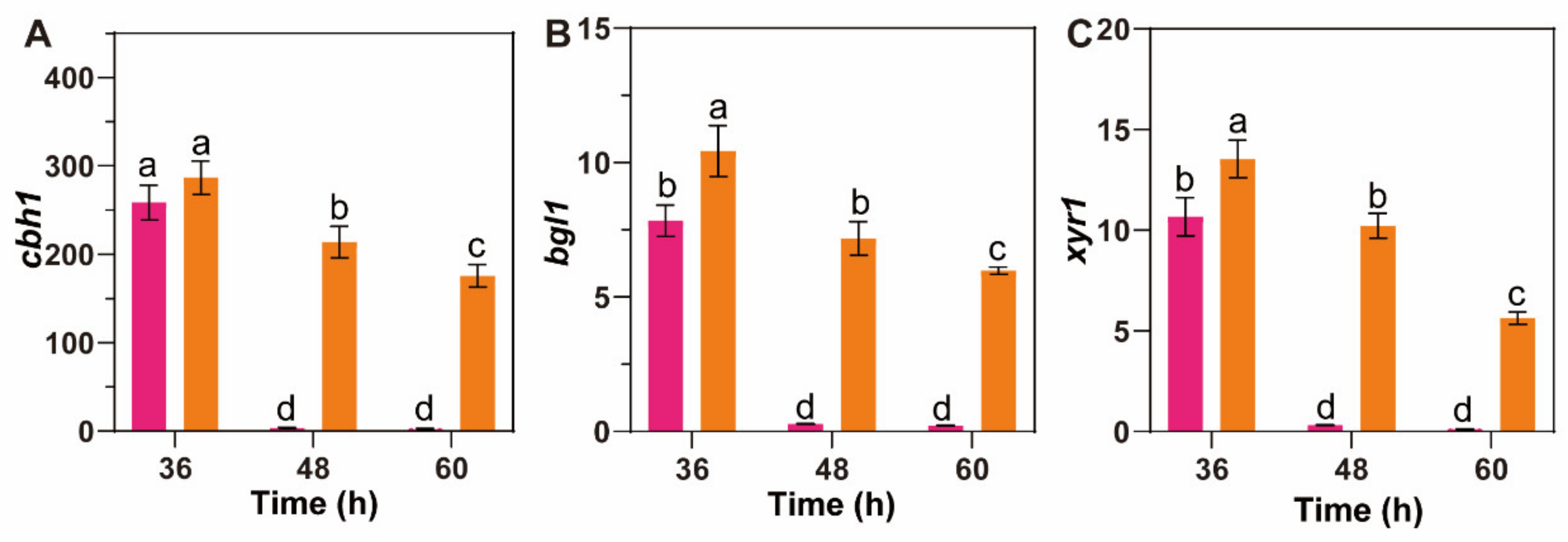
3.5. β-Glucosidase Suppression Promotes MGD to Persistently Induce Cellulase Synthesis in T. reesei
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Himmel, M.E.; Edward, A.B. Lignocellulose conversion to biofuels: Current challenges, global perspectives. Curr. Opin. Biotechnol. 2009, 20, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Reaney, M.; Wiens, D.; Tse, T. Production of bioethanol-A review of factors affecting ethanol yield. Fermentation 2021, 7, 268. [Google Scholar] [CrossRef]
- Liu, C.G.; Xiao, Y.; Xia, X.X.; Zhao, X.Q.; Peng, L.; Srinophakun, P.; Bai, F. Cellulosic ethanol production: Progress, challenges and strategies for solutions. Biotechnol. Adv. 2019, 37, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Prasoulas, G.; Gentikis, A.; Konti, A.; Kalantzi, S.; Kekos, D.; Mamma, D. Bioethanol production from food waste applying the multienzyme system produced on-site by Fusarium oxysporum F3 and mixed microbial cultures. Fermentation 2020, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Bhati, N.; Sharma, A.K. Cost-effective cellulase production, improvement strategies, and future challenges. J. Food Process Eng. 2021, 44, e13623. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Bikash, K.; Komal, A.; Pradeep, V. Current perspective on production and applications of microbial cellulases: A review. Bioresour. Bioprocess. 2021, 8, 95. [Google Scholar] [CrossRef]
- Peterson, R.; Nevalainen, H. Trichoderma reesei RUT-C30–thirty years of strain improvement. Microbiology 2012, 158, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Li, C.; Zhao, J.; Zhao, C. Biotechnological advances and trends in engineering Trichoderma reesei towards cellulase hyperproducer. Biotechnol. Bioprocess Eng. 2021, 26, 517–528. [Google Scholar] [CrossRef]
- Mandels, M.; Parrish, F.W.; Reese, E.T. Sophprpse as an inducer of cellulase in Trichoderma viride. J. Bacteriol. 1962, 83, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Humbird, D.; Davis, R.; Tao, L.; Kinchin, C.; Hsu, D.; Aden, A.; Schoen, P.; Lukas, J.; Olthof, B.; Worley, M.; et al. Process Design and Economics for Biochemical Conversion of Lignocellulosic Biomass to Ethanol: Dilute-Acid Pretreatment and Enzymatic Hydrolysis of Corn Stover; Technical Report NREL/TP-5100-47764; National Renewable Energy Laboratory: Golden, CO, USA, 2012. [CrossRef] [Green Version]
- Li, Y.; Liu, C.; Bai, F.; Zhao, X. Overproduction of cellulase by Trichoderma reesei RUT C30 through batch-feeding of synthesized low-cost sugar mixture. Bioresour. Technol. 2016, 216, 503–510. [Google Scholar] [CrossRef]
- Xia, Y.; Yang, L.; Xia, L. High-level production of a fungal β-glucosidase with application potentials in the cost-effective production of Trichoderma reesei cellulase. Process Biochem. 2018, 70, 55–60. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Xiong, L.; Mehmood, M.A.; Zhao, X.; Bai, F. On-site cellulase production and efficient saccharification of corn stover employing cbh2 overexpressing Trichoderma reesei with novel induction system. Bioresour. Technol. 2017, 238, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, P.; Zhu, D.; Yao, B.; Hasunuma, T.; Kondo, A.; Zhao, X. Efficient preparation of soluble inducer for cellulase production and saccharification of corn stover using in-house generated crude enzymes. Biochem. Eng. J. 2022, 178, 108296. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, C.; Fan, X.; Zhao, X.; Wang, W. Engineering of Trichoderma reesei for enhanced degradation of lignocellulosic biomass by truncation of the cellulase activator ACE3. Biotechnol. Biofuels 2020, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, X.; Zhang, F.; Peng, L.; Zhang, D.; Kondo, A.; Bai, F.; Zhao, X. Optimization of cellulolytic enzyme components through engineering Trichoderma reesei and on-site fermentation using the soluble inducer for cellulosic ethanol production from corn stover. Biotechnol. Biofuels 2018, 11, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yu, J.; Zhang, P.; Long, T.; Mo, Y.; Li, J.; Li, Q. Comparative transcriptome analysis of Trichoderma reesei reveals different gene regulatory networks induced by synthetic mixtures of glucose and β-disaccharide. Bioresour. Bioprocess. 2021, 8, 57. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, G.; Kou, Y.; Zhang, W.; Zhou, Q.; Chen, G.; Liu, W. Intracellular β-glucosidases CEL1a and CEL1b are essential for cellulase induction on lactose in Trichoderma reesei. Eukaryot. Cell 2014, 13, 1001–1013. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Xu, J.; Kou, Y.; Lv, X.; Zhang, X.; Zhao, G.; Zhang, W.; Chen, G.; Liu, W. Differential involvement of β-glucosidases from Hypocrea jecorina in rapid induction of cellulase genes by cellulose and cellobiose. Eukaryot. Cell 2012, 11, 1371–1381. [Google Scholar] [CrossRef] [Green Version]
- Qi, K.; Chen, C.; Yan, F.; Feng, Y.; Bayer, E.A.; Kosugi, A.; Cui, Q.; Liu, Y. Coordinated β-glucosidase activity with the cellulosome is effective for enhanced lignocellulose saccharification. Bioresour. Technol. 2021, 337, 125441. [Google Scholar] [CrossRef]
- Dashtban, M.; Qin, W. Overexpression of an exotic thermotolerant β-glucosidase in Trichoderma reesei and its significant increase in cellulolytic activity and saccharification of barley straw. Microb. Cell Factories 2012, 11, 63. [Google Scholar] [CrossRef] [Green Version]
- Treebupachatsakul, T.; Shioya, K.; Nakazawa, H.; Kawaguchi, T.; Morikawa, Y.; Shida, Y.; Ogasawara, W.; Okada, H. Utilization of recombinant Trichoderma reesei expressing Aspergillus aculeatus β-glucosidase I (JN11) for a more economical production of ethanol from lignocellulosic biomass. J. Biosci. Bioeng. 2015, 120, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhong, Y.; Zhao, X.; Wang, T. Development of the cellulolytic fungus Trichoderma reesei strain with enhanced β-glucosidase and filter paper activity using strong artifical cellobiohydrolase 1 promoter. Bioresour. Technol. 2010, 101, 9815–9818. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, H.; Kawai, T.; Ida, N.; Shida, Y.; Kobayashi, Y.; Okada, H.; Tani, S.; Sumitani, J.-I.; Kawaguchi, T.; Morikawa, Y.; et al. Construction of a recombinant Trichoderma reesei strain expressing Aspergillus aculeatus β-glucosidase 1 for efficient biomass conversion. Biotechnol. Bioeng. 2012, 109, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lin, F.; Li, Y.; Wei, W.; Wang, H.; Qin, L.; Zhou, Z.; Li, B.; Wu, F.-G.; Chen, Z. A β-glucosidase hyper-production Trichoderma reesei mutant reveals a potential role of cel3D in cellulase production. Microb. Cell Factories 2016, 15, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sels, J.P.J.; Huijberts, M.S.; Wolffenbuttel, B.H. Miglitol, a new α-glucosidase inhibitor. Expert Opin. Pharmacother. 1999, 1, 149–156. [Google Scholar] [CrossRef]
- Pérez, G.; Farina, L.; Barquet, M.; Boido, E.; Gaggero, C.; Dellacassa, E.; Carrau, F. A quick screening method to identify β-glucosidase activity in native wine yeast strains: Application of Esculin Glycerol Agar (EGA) medium. World J. Microbiol. Biotechnol. 2011, 27, 47–55. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Jalak, J.; Väljamäe, P. Mechanism of initial rapid rate retardation in cellobiohydrolase catalyzed cellulose hydrolysis. Biotechnol. Bioeng. 2010, 106, 871–883. [Google Scholar] [CrossRef]
- Törrönen, A.; Mach, R.L.; Messner, R.; Gonzalez, R.; Kalkkinen, N.; Harkki, A.; Kubicek, C.P. The two major xylanases from Trichoderma reesei: Characterization of both enzymes and genes. Nat. Biotechnol. 1992, 10, 1461–1465. [Google Scholar] [CrossRef]
- Xia, Y.; Yang, L.; Xia, L. Combined strategy of transcription factor manipulation and β-glucosidase gene overexpression in Trichoderma reesei and its application in lignocellulose bioconversion. J. Ind. Microbiol. Biotechnol. 2018, 45, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Saito, T.; Shinoda, S.; Haruki, M.; Hirano, N. In vitro assembly and cellulolytic activity of a β-glucosidase-integrated cellulosome complex. FEMS Microbiol. Lett. 2019, 366, fnz209. [Google Scholar] [CrossRef]
- Du, X.; Li, Y.; Xia, Y.; Ai, S.; Liang, J.; Sang, P.; Ji, X.; Liu, S. Insights into protein–ligand interactions: Mechanisms, models, and methods. Int. J. Mol. Sci. 2016, 17, 20144. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhai, Y.; Zhang, Y.; Xu, S.; Yu, S.; Wei, Y.; Xiao, H.; Song, Y. Inhibition of α-glucosidase by trilobatin and its mechanism: Kinetics, interaction mechanism and molecular docking. Food Funct. 2022, 13, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.; Arora, R.; Nandhagopal, N.; Kumar, S. Importance of chemical pretreatment for bioconversion of lignocellulosic biomass. Renew. Sustain. Energy Rev. 2014, 36, 91–106. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Ligand | Binding Energy/(Kcal/mol) |
|---|---|---|
| Bgl1 | Miglitol | −6.341 |
| Sophorose | −5.680 | |
| Cellobiose | −5.868 | |
| Bgl2 | Miglitol | −7.511 |
| Sophorose | −7.586 | |
| Cellobiose | −6.396 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, T.; Zhang, P.; Yu, J.; Gao, Y.; Ran, X.; Li, Y. Regulation of β-Disaccharide Accumulation by β-Glucosidase Inhibitors to Enhance Cellulase Production in Trichoderma reesei. Fermentation 2022, 8, 232. https://doi.org/10.3390/fermentation8050232
Long T, Zhang P, Yu J, Gao Y, Ran X, Li Y. Regulation of β-Disaccharide Accumulation by β-Glucosidase Inhibitors to Enhance Cellulase Production in Trichoderma reesei. Fermentation. 2022; 8(5):232. https://doi.org/10.3390/fermentation8050232
Chicago/Turabian StyleLong, Tingting, Peng Zhang, Jingze Yu, Yushan Gao, Xiaoqin Ran, and Yonghao Li. 2022. "Regulation of β-Disaccharide Accumulation by β-Glucosidase Inhibitors to Enhance Cellulase Production in Trichoderma reesei" Fermentation 8, no. 5: 232. https://doi.org/10.3390/fermentation8050232
APA StyleLong, T., Zhang, P., Yu, J., Gao, Y., Ran, X., & Li, Y. (2022). Regulation of β-Disaccharide Accumulation by β-Glucosidase Inhibitors to Enhance Cellulase Production in Trichoderma reesei. Fermentation, 8(5), 232. https://doi.org/10.3390/fermentation8050232