
Inactivation of Endogenous Pectin Methylesterases by Radio Frequency Heating during the Fermentation of Fruit Wines


 ,
, 
Abstract
:1. Introduction
2. Methods and Materials
2.1. Raw Materials and Sample Preparation
2.2. RF Heating Treatment
2.3. PME Activity Assay
2.4. Spectroscopy Analyses of PME
2.5. Determination of Methanol
2.6. Data Analysis and Statistics
3. Results and Discussion
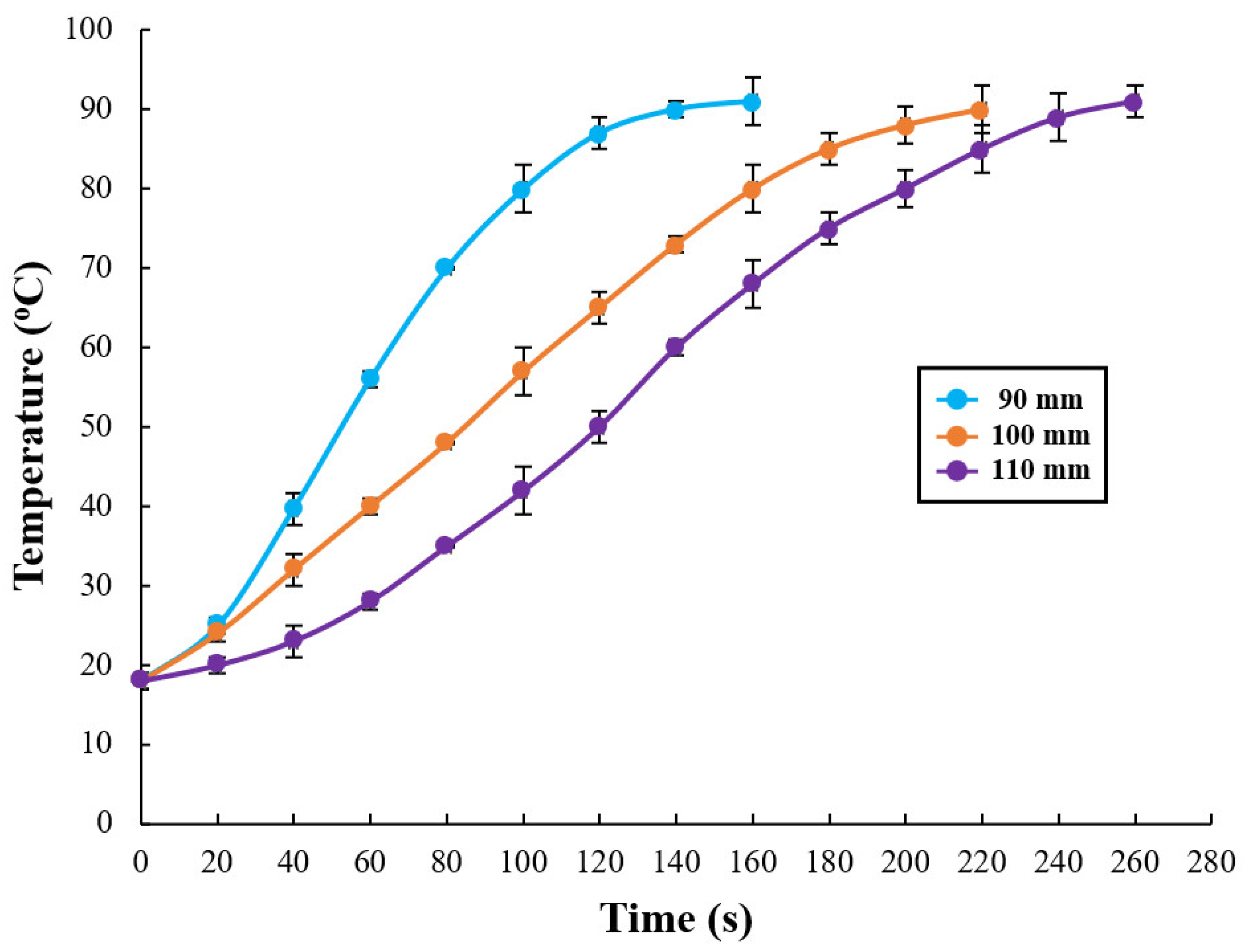
3.1. RF Heating Curve
3.2. Effects of RF Heating on PME Activity
3.3. Mechanisms of RF Heating Inactivation of PME
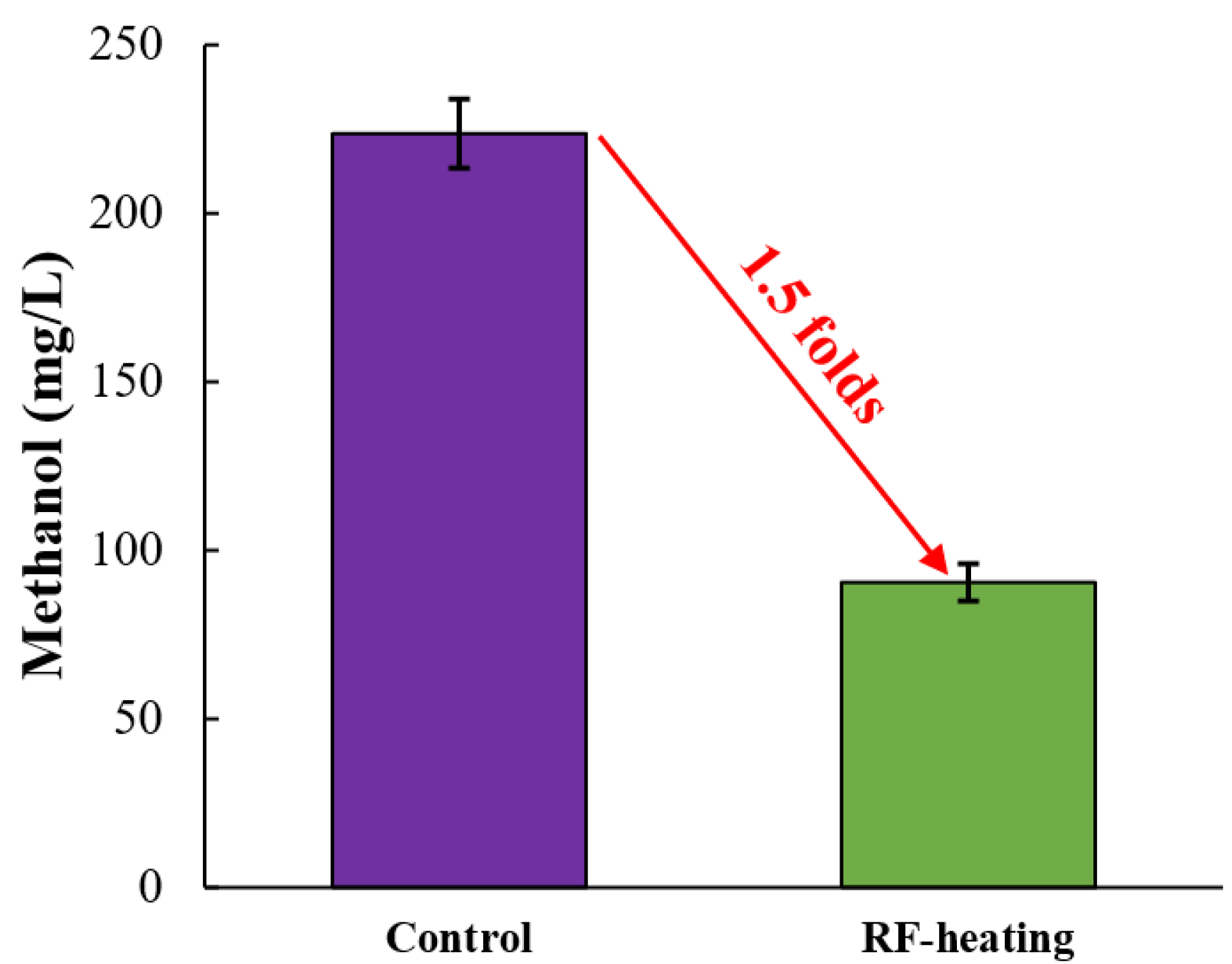
3.4. Effects of RF Heating on the Release of Methanol during Fermentation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jolie, R.P.; Duvetter, T.; Van Loey, A.M.; Hendrickx, M.E. Pectin methylesterase and its proteinaceous inhibitor: A review. Carbohydr. Res. 2010, 345, 2583–2595. [Google Scholar] [CrossRef] [PubMed]
- Takeya, T.; Yurimoto, H.; Sakai, Y. A Pichia pastoris single-cell biosensor for detection of enzymatically produced methanol. Appl. Microbiol. Biotechnol. 2018, 102, 7017–7027. [Google Scholar] [CrossRef] [PubMed]
- Prabhudev, H.; Sneharani, A.H. Extraction and characterization of pectin methylesterase from muskmelon biowaste for pectin remodeling. J. Food Biochem. 2020, 44, e13237. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Liu, X.; Gao, A.; Zhao, A.; Hu, J.; Li, B. Maintaining postharvest qualities of three leaf vegetables to enhance their shelf lives by multiple ultraviolet-C treatment. LWT 2016, 73, 1–5. [Google Scholar] [CrossRef]
- Moon, K.M.; Kwon, E.-B.; Lee, B.; Kim, C.Y. Recent Trends in Controlling the Enzymatic Browning of Fruit and Vegetable Products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef]
- Kim, A.-N.; Lee, K.Y.; Rahman, M.S.; Kim, H.-J.; Chun, J.; Heo, H.J.; Kerr, W.L.; Choi, S.-G. Effect of water blanching on phenolic compounds, antioxidant activities, enzyme inactivation, microbial reduction, and surface structure of samnamul (Aruncus dioicus var kamtschaticus). Int. J. Food Sci. Tech. 2020, 55, 1754–1762. [Google Scholar] [CrossRef]
- Xiao, H.-W.; Pan, Z.; Deng, L.-Z.; El-Mashad, H.M.; Yang, X.-H.; Mujumdar, A.S.; Gao, Z.-J.; Zhang, Q. Recent developments and trends in thermal blanching—A comprehensive review. Inf. Processing Agric. 2017, 4, 101–127. [Google Scholar] [CrossRef]
- Piyasena, P.; Dussault, C.; Koutchma, T.; Ramaswamy, H.S.; Awuah, G.B. Radio Frequency Heating of Foods: Principles, Applications and Related Properties—A Review. Crit. Rev. Food Sci. Nutr. 2003, 43, 587–606. [Google Scholar] [CrossRef]
- Ling, B.; Cheng, T.; Wang, S. Recent developments in applications of radio frequency heating for improving safety and quality of food grains and their products: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 2622–2642. [Google Scholar] [CrossRef]
- Guo, C.; Mujumdar, A.S.; Zhang, M. New Development in Radio Frequency Heating for Fresh Food Processing: A Review. Food Eng. Rev. 2019, 11, 29–43. [Google Scholar] [CrossRef]
- Yao, Y.; Sun, Y.; Cui, B.; Fu, H.; Chen, X.; Wang, Y. Radio frequency energy inactivates peroxidase in stem lettuce at different heating rates and associate changes in physiochemical properties and cell morphology. Food Chem. 2021, 342, 128360. [Google Scholar] [CrossRef] [PubMed]
- Fiore, A.; Di Monaco, R.; Cavella, S.; Visconti, A.; Karneili, O.; Bernhardt, S.; Fogliano, V. Chemical profile and sensory properties of different foods cooked by a new radiofrequency oven. Food Chem. 2013, 139, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hu, C.; Sun, Y.; Zhang, X.; Wang, Y.; Fu, H.; Chen, X.; Wang, Y. Blanching effects of radio frequency heating on enzyme inactivation, physiochemical properties of green peas (Pisum sativum L.) and the underlying mechanism in relation to cellular microstructure. Food Chem. 2021, 345, 128756. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, X.; Zhu, F.; Liu, G.; Chu, L.; Yan, X.; Ma, Y.; He, F.; Li, G.; Zhang, Y.; et al. Effects of Different Winemaking Yeasts on the Composition of Aroma-Active Compounds and Flavor of the Fermented Jujube Wine. Processes 2021, 9, 970. [Google Scholar] [CrossRef]
- Downie, B.; Dirk, L.M.A.; Hadfield, K.A.; Wilkins, T.A.; Bennett, A.B.; Bradford, K.J. A Gel Diffusion Assay for Quantification of Pectin Methylesterase Activity. Anal. Biochem. 1998, 264, 149–157. [Google Scholar] [CrossRef]
- Liu, W.; Liu, J.; Liu, C.; Zhong, Y.; Liu, W.; Wan, J. Activation and conformational changes of mushroom polyphenoloxidase by high pressure microfluidization treatment. Innov. Food Sci. Emerg. Technol. 2009, 10, 142–147. [Google Scholar] [CrossRef]
- Liao, X.; Zhang, Y.; Bei, J.; Hu, X.; Wu, J. Alterations of molecular properties of lipoxygenase induced by dense phase carbon dioxide. Innov. Food Sci. Emerg. Technol. 2009, 10, 47–53. [Google Scholar] [CrossRef]
- Qin, Y.; Shin, J.-A.; Lee, K.-T. Determination of acetaldehyde, methanol and fusel oils in distilled liquors and sakès by headspace gas chromatography. Food Sci. Biotechnol. 2020, 29, 331–337. [Google Scholar] [CrossRef]
- Jiao, Y.; Tang, J.; Wang, S.; Koral, T. Influence of dielectric properties on the heating rate in free-running oscillator radio frequency systems. J. Food Eng. 2014, 120, 197–203. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Zhang, X.; Shi, Q.; Xin, L.; Fu, H.; Wang, Y. Effects of radio frequency assisted blanching on polyphenol oxidase, weight loss, texture, color and microstructure of potato. Food Chem. 2018, 248, 173–182. [Google Scholar] [CrossRef]
- Moens, L.G.; De Laet, E.; Van Ceunebroeck, J.-C.; Van Loey, A.M.; Hendrickx, M.E.G. Thermal inactivation of pectin methylesterase from different potato cultivars (Solanum tuberosum L.). LWT 2021, 138, 110600. [Google Scholar] [CrossRef]
- Siguemoto, É.S.; Pereira, L.J.; Gut, J.A.W. Inactivation kinetics of pectin methylesterase, polyphenol oxidase, and peroxidase in cloudy apple juice under microwave and conventional heating to evaluate non-thermal microwave effects. Food Biopro. Technol. 2018, 11, 1359–1369. [Google Scholar] [CrossRef]
- Wilińska, A.; de Figueiredo Rodrigues, A.S.; Bryjak, J.; Polakovič, M. Thermal inactivation of exogenous pectin methylesterase in apple and cloudberry juices. J. Food Eng. 2008, 85, 459–465. [Google Scholar] [CrossRef]
- Yeom, H.W.; Streaker, C.B.; Zhang, Q.H.; Min, D.B. Effects of Pulsed Electric Fields on the Activities of Microorganisms and Pectin Methyl Esterase in Orange Juice. J. Food Sci. 2000, 65, 1359–1363. [Google Scholar] [CrossRef]
- Damodaran, S. In situ measurement of conformational changes in proteins at liquid interfaces by circular dichroism spectroscopy. Anal. Bioanal. Chem. 2003, 376, 182–188. [Google Scholar] [CrossRef]
- Jenkins, J.; Pickersgill, R. The architecture of parallel β-helices and related folds. Prog. Biophys. Mol. Biol. 2001, 77, 111–175. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, Z.; Chen, J.; Fu, H.; Subbiah, J.; Chen, X.; Wang, Y. Effects of radio frequency heating treatment on structure changes of soy protein isolate for protein modification. Food Bioprocess Technol. 2017, 10, 1574–1583. [Google Scholar] [CrossRef]
- Ling, B.; Ouyang, S.; Wang, S. Effect of radio frequency treatment on functional, structural and thermal behaviors of protein isolates in rice bran. Food Chem. 2019, 289, 537–544. [Google Scholar] [CrossRef]
- Kangur, L.; Timpmann, K.; Zeller, D.; Masson, P.; Peters, J.; Freiberg, A. Structural stability of human butyrylcholinesterase under high hydrostatic pressure. Biochim. Biophys. Acta 2019, 1867, 107–113. [Google Scholar] [CrossRef]
- Botelho, G.; Anjos, O.; Estevinho, L.M.; Caldeira, I. Methanol in Grape Derived, Fruit and Honey Spirits: A Critical Review on Source, Quality Control, and Legal Limits. Processes 2020, 8, 1609. [Google Scholar] [CrossRef]
- Frenkel, C.; Peters, J.S.; Tieman, D.M.; Tiznado, M.E.; Handa, A.K. Pectin Methylesterase Regulates Methanol and Ethanol Accumulation in Ripening Tomato (Lycopersicon esculentum) Fruit. J. Biol. Chem. 1998, 273, 4293–4295. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (s) | α-Helix | β-Sheet | β-Turn | Random Coil |
|---|---|---|---|---|
| 0 | 32.2 ± 0.3 | 20.2 ± 0.1 | 20.9 ± 0.1 | 29.9 ± 0.5 |
| 20 | 32.4 ± 0.2 | 20.3 ± 0.4 | 21.0 ± 0.5 | 29.9 ± 0.1 |
| 40 | 33.6 ± 0.5 | 20.3 ± 0.2 | 20.7 ± 0.2 | 30.2 ± 0.2 |
| 60 | 33.1 ± 0.1 | 19.9 ± 0.4 | 20.3 ± 0.6 | 29.2 ± 0.5 |
| 80 | 31.3 ± 0.2 | 18.9 ± 0.1 | 19.3 ± 0.1 | 27.6 ± 0.2 |
| 100 | 32.5 ± 0.4 | 18.2 ± 0.6 | 19.5 ± 0.2 | 32.4 ± 0.1 |
| 120 | 33.1 ± 0.6 | 17.9 ± 0.5 | 18.5 ± 0.3 | 25.5 ± 0.2 |
| 140 | 32.4 ± 0.1 | 13.8 ± 0.2 | 17.6 ± 0.1 | 38.5 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Yu, X.; Zhao, W.; Li, G.; Liu, G.; Ma, Y.; Chu, L.; Ma, Y.; Zhang, Y.; Lu, Y.; et al. Inactivation of Endogenous Pectin Methylesterases by Radio Frequency Heating during the Fermentation of Fruit Wines. Fermentation 2022, 8, 265. https://doi.org/10.3390/fermentation8060265
Zhao Y, Yu X, Zhao W, Li G, Liu G, Ma Y, Chu L, Ma Y, Zhang Y, Lu Y, et al. Inactivation of Endogenous Pectin Methylesterases by Radio Frequency Heating during the Fermentation of Fruit Wines. Fermentation. 2022; 8(6):265. https://doi.org/10.3390/fermentation8060265
Chicago/Turabian StyleZhao, Yan, Xiaobin Yu, Wei Zhao, Gen Li, Guangpeng Liu, Yanrui Ma, Le Chu, Yinfei Ma, Ying Zhang, Yao Lu, and et al. 2022. "Inactivation of Endogenous Pectin Methylesterases by Radio Frequency Heating during the Fermentation of Fruit Wines" Fermentation 8, no. 6: 265. https://doi.org/10.3390/fermentation8060265
APA StyleZhao, Y., Yu, X., Zhao, W., Li, G., Liu, G., Ma, Y., Chu, L., Ma, Y., Zhang, Y., Lu, Y., He, F., & Liu, X. (2022). Inactivation of Endogenous Pectin Methylesterases by Radio Frequency Heating during the Fermentation of Fruit Wines. Fermentation, 8(6), 265. https://doi.org/10.3390/fermentation8060265