To obtain a better understanding of the impact of fermentation parameters on the interactions between S. cerevisiae and L. thermotolerans, and consequently on the transformation of must into wine, it was vital to study fermentation through a global approach. To reach this goal, the population dynamics were first studied due to the direct influence of the environment on yeast development. Then, fermentation kinetics and volatile metabolites production were assessed in that order, as they not only depend on environmental conditions, but also on the population dynamics and yeast–yeast interactions that took place during the fermentation process. The resulting combination of these three datasets made it possible to better understand the overall impacts of each factor studied.
3.1. Impact of Fermentation Parameters on Population Dynamics
The first objective was to study how fermentation parameters such as nitrogen content, inoculum ratio, oxygenation, and temperature could impact the development of both yeasts: how their maximal growth rate and maximal population were modified and, mainly, which strain would be predominant throughout fermentation. Here, the aim was not to study the impact of chosen parameters independently as in previous studies, but simultaneously through the experimental design to allow for a comparison between effects.
The impacts of fermentation parameters on variables describing the population dynamics are given in
Table 3. For each parameter/variable couple, the standard effect,
p-value, and contribution to variance are shown. The standardized effect provides information on the nature and level of the impact of the parameter on the variable: an increase in the parameter chosen leads to an increase in the variable studied when the standardized effect is positive, and to a decrease when the standardized effect is negative. The
p-value provides information on the significance of this variation, and the contribution to variance indicates the importance of the parameter chosen in explaining the total variation of the variable studied.
Regarding
Saccharomyces cerevisiae, the growth rate µ
max was positively and significantly impacted by temperature (stand. effect 4.094) and not by the other factors (contribution to variance of 73.4% for temperature). In contrast, the growth rate of
Lachancea thermotolerans seemed to be impacted only by the inoculum ratio
Sc/
Lt, and at the limit of significance (stand. effect −3.010,
p-value 0.057): µ
max Lt tended to be higher when the inoculum ratio favored
Lt. In this study, oxygenation had no impact on the growth rate of either yeast, whereas previous works showed a positive effect on the µ
max of
Lt [
20]. No significant effect of nitrogen content was observed here, despite nitrogen sources having been previously shown to influence the growth of yeasts differently according to the nature and quantity of nitrogen compounds [
21,
35]. However, nitrogen content significantly impacted the growth rate only up to around 100 mg/L, depending on the species and strains of
Sc [
36]. As this “limiting concentration” was below both our chosen nitrogen levels (in line with our goal of avoiding nitrogen-scarce conditions), this explained the lack of a significant effect of this particular parameter on growth rate.
Thus, a decrease in temperature favored the faster development of
Lt versus
Sc: the µ
max Lt/µ
max Sc ratio was indeed higher at 18 °C than at 28 °C (stand. effect −3.594), which supported previous results [
16]. Although both
Lt and
Sc were able to grow at 15–20 °C and even better at 25–30 °C on YPD agar [
37], temperature affected the population dynamics in mixed cultures [
38,
39].
S. cerevisiae’s ability to grow at higher temperatures than most yeasts gives it an advantage: the temperature increase during fermentation can lead to its dominance versus non-
Saccharomyces species during wine fermentation [
40,
41,
42,
43]. On the contrary, a decrease in temperature can enhance the growth and survival of non-
Saccharomyces yeasts by attenuating their susceptibility to ethanol [
29,
44]. In addition, culture at low temperature could induce an increase in the intracellular level of ROS (reactive oxygen species) and an oxidative stress response (through mechanisms that are still not well understood) in
S. cerevisiae: this can lead to a decrease in its maximal growth rate and reduce its competitiveness against other yeasts [
36,
45]. These different factors can provide an explanation for the significant impact of temperature on growth rates, both absolute (µ
max Sc and µ
max Lt) and relative (µ
max Lt/µ
max Sc).
Otherwise, in the conditions tested, the maximal population of both yeasts was affected differently by the factors studied. For Sc, the Nmax was significatively influenced by both temperature (positive effect, stand. effect 4.546, contrib. var. 45.7%) and oxygenation (negative effect, stand. effect −4.443, contrib. var. 43.7%). On the contrary, the maximal population of Lt was significantly impacted by temperature (which explained 77.5% of the variance, with a standard effect of −7.750) and with low significance by nitrogen (p-value 0.052, stand. effect 3.131).
Thus, an increase in temperature from 18 °C to 28 °C led to a higher
Sc and a lower
Lt population. Gobbi et al. (2013) [
16] also described a lower maximal population of
Sc when the temperature was decreased, but they did not observe a higher
Lt population (they noticed the same population with a higher persistence during fermentation). In addition to temperature, in other studies [
17,
20] oxygenation favored the development of
Lt and led to a higher maximal population, which was not observed as significant here. Garcia et al. (2021) [
46] also showed that the survival of
Lt in ethanol-stress conditions was enhanced in the presence of oxygen. The fact that such an impact was not observed here could be explained by differences in the oxygenation levels studied. Indeed, in this study, simple sparging of compressed air at the end of exponential growth was used to stay closer to cellar capabilities, although the works cited made use of either constant aerobic conditions [
20,
46] or multiple oxygen pulses over the fermentation process [
17].
Concerning nitrogen, a higher concentration in the medium seemed to positively affect the N
max Lt (stand. effect 3.131) but with low significance (
p-value 0.052). As previously discussed for µ
max, the chosen nonscarce nitrogen conditions may explain this lack of significant impact on this population variable. The incubation ratio had no significant effect on the maximal population obtained for both yeasts, thereby supporting the work of Comitini et al. (2011) [
47], who observed the same maximum population (but with longer persistence) of
Lt when favored by the inoculum ratio.
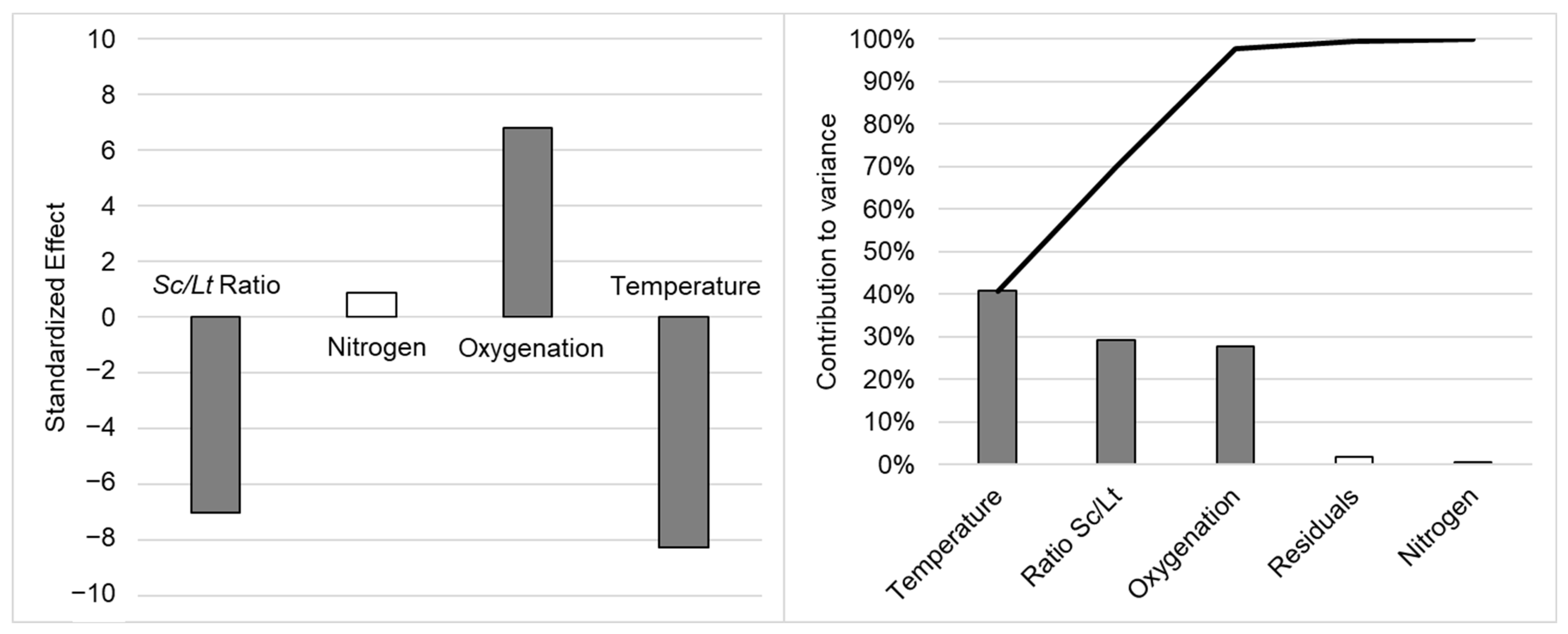
As the effects observed on the µ
max and N
max were different, the AUC was calculated to assess the impact of fermentation parameters on the population dynamics more precisely (see additional data in
Table A1). Yeast distribution over time (AUC
Lt/AUC
Sc) was significantly and mainly managed by temperature (var. contribution 40.80%, stand. effect −8.267), but also by yeast ratio (var. contribution 29.20%, stand. effect −6.999) and oxygenation (var. contribution 27.80%, stand. effect 6.822), as illustrated in
Figure 1.
A lower fermentation temperature, an inoculation ratio favoring
Lt (lower
Sc/
Lt ratio), and air sparging at the end of exponential growth phase all led to a significant increase in the overall presence of
Lt compared to
Sc throughout fermentation (higher AUC
Lt/AUC
Sc). The inoculation ratio was quite straightforward, favoring
Lt during the early days of culture, either directly by increasing its relative population at inoculation, or more indirectly by limiting the
Sc population early on and thus reducing its potential negative interactions toward
Lt. Oxygenation probably acted as described in the literature, as hypothesized by Shekhawat et al., by allowing the
Lt population to better fulfill its oxygen needs, consequently allowing for better persistence over time and leading to higher cell counts through the stimulation of the respiratory metabolism over the fermentative one [
17]. Moreover, Varela et al. (2021) [
48] observed the population dynamics during the spontaneous fermentation of natural grape musts and also showed that aeration led to an increased proportion of non-
Saccharomyces yeasts in the natural consortium, possibly related to the modification of membrane lipid composition, conferring a higher ethanol tolerance to the yeasts in question. The temperature effect on the AUC ratio probably resulted from previously discussed effects on the µ
max, with quicker
Sc growth inducing a quicker takeover of the culture media.
Considering these very important impacts of temperature, ratio, and oxygenation on the population dynamics (especially the AUC ratio), only two conditions (D and G: 18 °C, ratio
Sc/
Lt 0.1, oxygenation) led to the predominance of
Lt on
Sc and to the persistence of
Lt with a high population throughout fermentation (see additional data in
Figure A1). The results also showed that the amount of nitrogen in the culture medium had no significant effect on the population dynamics when modulated between the two levels chosen (150 mg·L
−1 and 300 mg·L
−1), except for a small positive but nonsignificant (
p-value = 0.052) effect on the N
max for
Lt. As discussed for the single variables, the nature of nitrogen sources (ammonium, amino acids, or both) can impact maximal growth and maximal population of
Lt in pure culture, but with strain-dependent variability [
21,
35,
49]. In addition, the consumption kinetics of amino acids can vary according to the amino acid and differ from those observed in
Sc, leading to possible competition in
Sc/
Lt coculture. Nevertheless, Kemsawad et al. (2015) noticed that
Lt could grow and maintain itself even if no nitrogen was present in the synthetic medium, making use of autolysis phenomena to obtain nitrogen compounds [
21]. In addition, the amount of nitrogen, even at the lowest value, is usually sufficient to allow for yeast growth and complete fermentation, as a minimum of 140 mg/L is usually recommended [
22]. For all these reasons, the result observed here could be explained by the fact that the experimental conditions chosen did not aim to emulate complete nitrogen scarcity, or by a strain-specific tolerance to these lower levels of nitrogen, or both.
By studying the different fermentation factors, we expected to see an effect of all these factors on the population dynamics, as observed by other authors. Nevertheless, when all these factors were combined, their relative importance could be seen. It seemed in the conditions studied that overall, temperature was the most important factor in optimizing Lt presence over time during fermentation (AUCLt/AUCSc ratio). In addition to temperature, oxygenation and the inoculum ratio also appeared to be significant, but were still secondary factors in obtaining the persistence or predominance of Lt over Sc throughout the fermentation process.
To our knowledge, this is the first time information has been obtained regarding the combined effect of different fermentation parameters on the population dynamics. Such information constitutes a tool to control proper Lt implantation, and also to help in determining which yeast (Sc or Lt) will dominate the other during a mixed culture with given or adjustable parameters. This ensures both that Lt will have enough overall persistence to potentially influence final wine quality and that Sc will still persist to ensure fermentation does not go on for too long or become stuck.
3.2. Impact of Fermentation Parameters on Fermentation Time and Physicochemical Variables
As mentioned in the general introduction of this study, both Sc and Lt have different fermentative capabilities when used in pure culture, with Sc leading to a shorter fermentation and a higher amount of ethanol, and Lt culture leading to less alcohol and a decrease in pH via lactic acid production, but with a higher fermentation time. The goal of an optimized mixed fermentation is to obtain a compromise between the properties of both yeasts. Thus, after having studied the influence of the different parameters on the population dynamics, it was necessary to study their general impact on fermentation to see how both these different population equilibria and the more direct impact of the yeast metabolism parameters chosen could modify the characteristics of the medium.
The effects of incubation ratio, nitrogen content, oxygenation, and temperature on fermentation are given in
Table 4.
Fermentation took between 7 and 16 days to obtain the total consumption of sugars, depending on the conditions tested (see additional data in
Figure A1 and
Table A2). Thus, the choice of environmental conditions could considerably affect the resulting fermentation kinetics.
The four factors studied had different impacts on fermentation time. Oxygenation seemed to lead to longer fermentation times (stand. effect 3.086), which was to be expected, given that it was shown above to favor
Lt more than
Sc. On the contrary, an increase in the
Sc/
Lt ratio or in the amount of nitrogen resulted in a decrease in fermentation time (stand. effect −3.057 and −2.384, respectively, but with a slight significance (
p-value 0.055 and 0.097, respectively)) due to their positive impact on the
Sc population. An increase in temperature led to a shorter fermentation (stand. effect −7.122,
p-value 0.006). This very significant effect (which explained 64.8% of the variance) was certainly linked to the positive effect of temperature on the
Sc population (see
Section 3.1), and supported the work of Gobbi et al. (2013) [
16].
Ethanol production reached values of about 11–12% ethanol (
v/
v) at the end of the fermentation for all the conditions tested (see additional data in
Table A2). This production was not impacted by temperature or the amount of nitrogen, but was impacted, albeit with a low significance (
p-value of 0.063 and 0.065, respectively), by the
Sc/
Lt ratio and oxygenation. An increase in the
Sc/
Lt ratio led to an increase in
Sc at the expense of
Lt (see
Section 3.1.) and to an increase in ethanol production (stand. effect 2.899). On the contrary, oxygenation, which led to a decrease in the
Sc compared to
Lt population (see
Section 3.1), led at the same time to a decrease in ethanol production (stand. effect −2.857). As the Crabtree effect, which was weaker in
Lt than in
Sc, favors respiration and leads to lower ethanol production by
Lt [
11], this lower ethanol production could be explained by the modulation of this metabolic pathway. Moreover, the ethanol yield of
Lt has also been described as being significantly lower than for
Sc [
12]. Although the respective impacts of these two hypotheses were impossible to properly determine within the scope of this study, the overall negative impact of oxygenation on ethanol production was still observable, and could be of interest to winemakers willing to decrease the ethanol content of their wines.
The final pH was not significantly impacted by temperature, oxygenation, or nitrogen content. The
Sc/
Lt ratio had a positive effect on pH (stand. effect 2.575, var. contrib. 57.90%). The pH values observed when the inoculum ratio was 0.1 (predominance of
Lachancea thermotolerans) were lower than those obtained with a ratio of 1. This pH reduction in mixed cultures
Sc/
Lt compared to
Sc pure culture is often observed whatever the conditions (modalities of inoculation, temperature, medium) [
16].
This decrease in pH could be linked at least partially to the production of lactic acid.
Lachancea thermotolerans, as discussed above, is specifically known amongst non-
Saccharomyces for its ability to transform pyruvate into this acid through lactate dehydrogenases [
11,
50]. As
S. cerevisiae is not able to produce lactic acid [
51] or, depending on the strain, can produce it in trace amounts only [
50], the final amount of this acid can be directly related to the population dynamics, and specifically to the
Lt population. Conditions either favoring
Lt development compared to
Sc or inhibiting antagonistic interactions from
Sc that limit
Lt growth or persistence will therefore indirectly favor lactic acid production during the fermentation process.
In the different conditions tested, the production of lactic acid varied between 0.82 and 3.52 g·L
−1. These values are classical ones as the production of this acid is highly dependent on the strain (<0.1 to >15 g·L
−1) and on conditions [
11,
50,
52,
53]. Lactic acid concentration was considerably impacted by the
Sc/
Lt ratio (var. contrib. 83.30%, stand. effect −7.322). A decrease in this ratio indeed favored the predominance of
Lt in the earlier stages of the fermentation, and this species’ lactic acid production has previously been shown to occur predominantly at the beginning of fermentation, during exponential growth [
2,
6,
16,
54]. This hypothesis was further supported by the pH dynamics shown in
Figure 2, in which a rapid pH decrease during the first 72 h can be observed, with this falloff being on average significantly greater for conditions with an
Sc/
Lt ratio of 0.1.
Using an
Sc/
Lt ratio of 0.1 in mixed culture in natural must, Gobbi et al. (2013) [
16] showed that the effects of the interaction between
Lt and
Sc on fermentation (decreases in ethanol and pH, increase in lactic acid compared to a pure culture of
Sc) were more significant at 20 °C than at 30 °C, but this could have been related to the better persistence of
Lt in these conditions (as also observed earlier in this study—see
Section 3.1).
Morata et al. (2018) also observed that a higher lactic acid production was correlated with a more numerous
Lt population and a higher nitrogen content (up to 500 mg/L) [
54]. This impact of nitrogen was not significant here within our chosen concentration levels.
Lactic acid production did not appear to be impacted by oxygenation, although Shekhawat et al. (2020) showed an induction of lactate dehydrogenase gene expression in pure cultures of
Lt and the redirection of metabolism toward lactic acid in response to anaerobic conditions [
55]. However, the direct impact of oxygenation conditions on effective lactic acid production has not yet been shown in the current literature, and conclusions are therefore hard to draw based only on transcriptomic results.
This study highlighted the key role of temperature to control fermentation time over other factors that did not have as much impact or significance. This role can be linked to the population dynamics seen in
Section 3.1, and confirmed that overfavoring
Lt in a mixed culture can lead to sluggish or stuck fermentations, an eventuality that must be avoided. Decreasing the ethanol concentration in these
Sc/
Lt cultures was difficult in these conditions, since the parameters chosen had no or very little impact on ethanol production. In contrast, the inoculum ratio turned out to be the most important factor in modulating lactic acid production, without it being necessary to modulate other fermentation parameters. It could be an interesting tool to control wine acidification without drastic changes in usual winemaking practices.
3.3. Impact of Fermentation Parameters on Volatile Compound Production
As the production of aroma compounds is variable according to previous studies [
2], it was also interesting to observe this production in the mixed
Sc/
Lt culture and study the influence of fermentation parameters, as the volatile compounds produced modified the final sensorial characteristics of the wine. As seen above, the parameters chosen had a significant impact on the population dynamics, a modulation that also had repercussions on fermentation variables such as fermentation time and lactic acid production. We therefore assumed it would also impact the production of aroma compounds in a similar way, both directly (metabolism modifications) and indirectly (population dynamics).
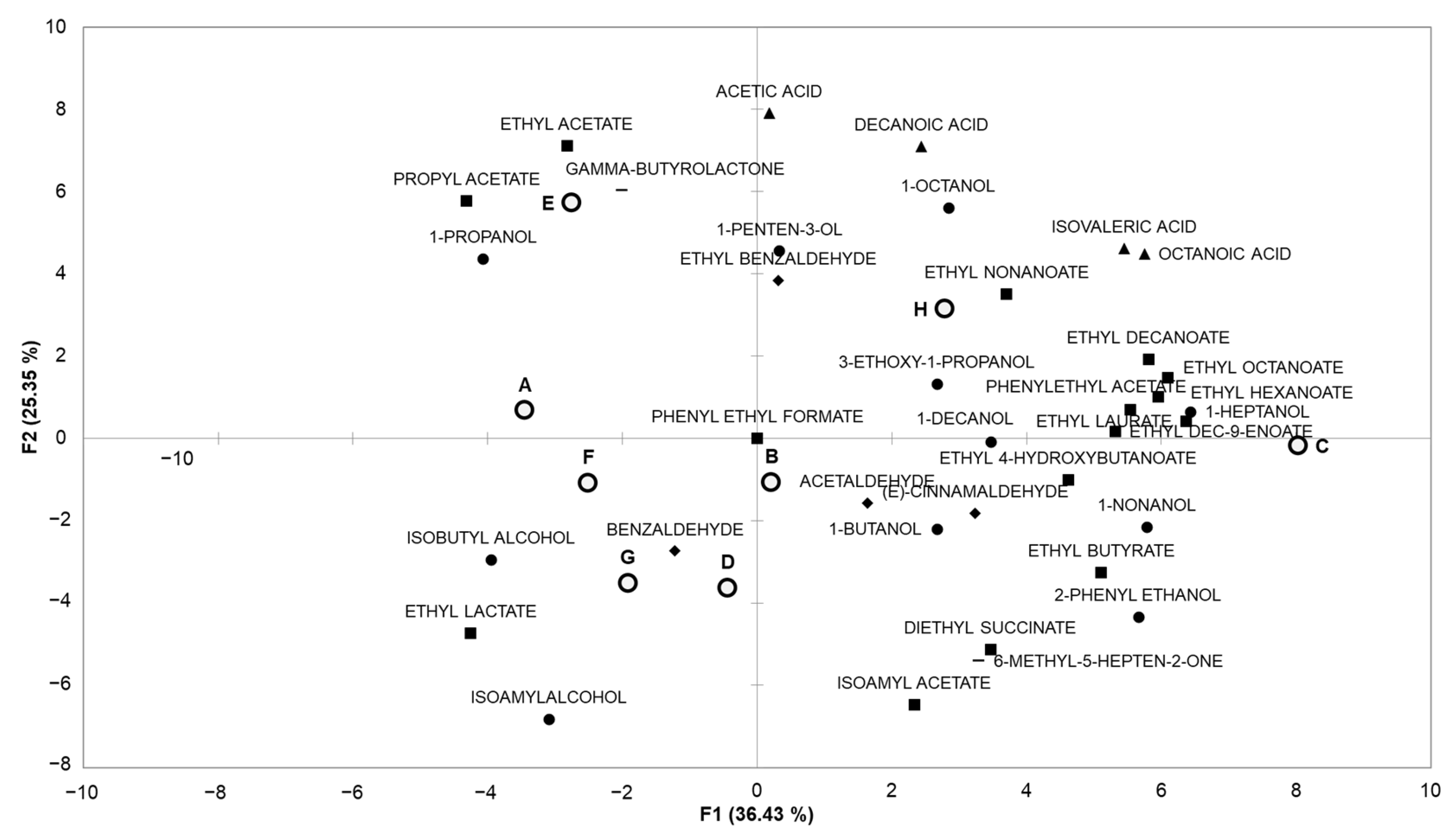
The volatile compounds present in the final synthetic wines obtained with the different conditions tested were subjected to a principal component analysis. The first two principal components, F1 and F2, explained 36.4 and 25.4% of the variance, respectively, as shown in
Figure 3.
Axis F1 marks a separation between four groups: condition E, which is close to ɣ-butyrolactone and esters with short chains; condition H, which is closer to carboxylic acids; conditions G and D, which are close to benzaldehyde (conditions corresponding to the predominance of Lt throughout the fermentation); and the other conditions.
Axis F2 separates condition C, which is close to esters derived from C6 to C10 carboxylic acids; condition H, which is closer to alcohols and acids; and the other conditions
Some conditions (A, B, D, F, G) led to media with quite similar volatilomes and others (C, E, H) with different volatile compound profiles. These results showed that the fermentation parameters studied seemed to broadly impact the production of aroma compounds, and could therefore be useful in modifying the volatile composition of the final wine.
A statistical analysis was performed first on the results concerning each class (aroma compounds grouped into alcohols, aldehydes, ketones, carboxylic acids, and esters), then on the results concerning each volatile molecule. The results are presented in
Table 5,
Table 6,
Table 7 and
Table 8.
The
Sc/
Lt ratio had a significant positive impact on the production of carboxylic acids and esters (stand. effect 6.199 and 2.614, respectively) and a negative impact on alcohol production (stand. effect −9.037). The
Sc population seemed to be mainly linked to the formation of acids and esters. This was in line with the work of Morales et al. (2019) [
23], who showed an increase in ethyl esters, acetals, and acids, as well as a decrease in alcohols, when the proportion of
Saccharomyces cerevisiae increased in the inoculum. As the
Sc/
Lt ratio significantly impacted the proportion of each yeast in the media over time (as seen in
Section 3.1), and as these two species did not produce the same aroma compounds [
16,
20,
23], the aroma profile of a given mixed culture was indeed expected to be very different from either pure culture. However, interactions between yeasts can also change their metabolism and their aroma production, resulting in an apparent synergy (higher amount in mixed culture compared to pure cultures), as observed previously [
16,
23].
The amount of nitrogen had a significant effect (stand. effect 4.742) only on alcohol production. Rollero et al. (2021) highlighted the link between nitrogen sources and aroma production in cocultures of
Sc with several non-
Saccharomyces yeasts [
56]. Higher alcohols can be produced from both sugars and amino acids: differences in the consumption kinetics of nitrogen sources and in metabolism flux according to the yeast can then lead to various productions of aroma compounds. Some authors also found that modifying nitrogen levels can lead to the production of different volatile compounds in pure
Sc cultures [
22], thus correlating with this conclusion.
Oxygenation had no significant impact on the production of the different families of volatile compounds. This was in contradiction with the results of Shekhawat et al. (2017), who noticed an increase in the production of alcohols and volatile acids and a decrease in medium-chain fatty acids and corresponding esters [
20]. However, the differences in oxygenation protocols already discussed in
Section 3.1 could explain this lack of a significant effect in the conditions of our study.
Temperature had a negative impact on the production of alcohols and ketones (stand. effect −10.175 and −3.825, respectively), a positive effect on ester and acid production (stand. effect 4.514 and 2.685, respectively), and no significant effect on aldehyde production. As temperature was previously shown to significantly impact population dynamics in this study, such shifts in aroma production were to be expected, as the media was fermented by very different yeast populations depending on the temperature chosen.
An increase in aroma production, notably esters, was observed previously at low temperature with some strains of
S. cerevisiae, and this increase seemed to be related to a change in membrane lipid composition in response to the stress linked to the temperature decrease [
57,
58].
More specifically, regarding the volatile molecules produced during fermentation, several alcohols are produced: primary alcohols with 3 to 10 carbons, isobutyl alcohol, isoamyl alcohol, 2-phenylethanol, etc.
Table 6 shows the impacts of fermentation parameters on the production of these alcohols. These results are also illustrated in
Figure 4, which presents the standardized effects of each fermentation parameter on this production.
A significant impact of environmental factors was observed only on three molecules: 1-propanol, isoamyl alcohol, and 2-phenylethanol. For the other alcohols, the contribution of residuals to total variance was high, but no significant impact of interactions between factors could be observed, even when examined in more detail. This furthered the hypothesis that this lack of observed effects was due to a real absence of effects of the conditions studied on the production of these molecules.
The production of 1-propanol was significantly influenced by nitrogen content (negatively, stand. effect −3.271) and to a lesser extent (
p-value 0.097) by temperature (positively, stand. effect 2.392). The negative influence of nitrogen content showed the complexity of forecasting the production of an aroma compound in a coculture. Propanol is indeed produced from amino acids (while other alcohols can come from both amino acids and sugars) [
56]: an increase in nitrogen content should have favored its production. Gobbi et al. (2013) [
16] and Balikci et al. (2016) [
59] observed a higher production of propanol by
Lt compared to
Sc in pure culture, in contrast to the observations of Morales et al. (2019) [
23] in a sugar-rich must. Vicente et al. (2021) noticed that
L. thermotolerans usually produced low amounts of higher alcohols, but with a high strain variability [
11]. Temperature had a positive effect (stand. effect 2.392) on the production of 1-propanol, contrary to the results of Gobbi et al. (2013) [
16]. All these conflicting observations highlighted the variability of propanol production, depending on strains, media, and culture conditions chosen, especially in mixed cultures [
16,
23,
53,
59].
The production of 2-phenylethanol was impacted mainly by nitrogen content (positively, stand. effect 9.281) and temperature (negatively, stand. effect −9.895). Gonzalez et al. (2018) observed that nutrient conditions (nitrogen sources, glucose amount) affected the production of phenylethanol (a molecule that can play a quorum-sensing role) with other non-
Saccharomyces yeasts [
60]. Fairbairn et al. (2021) noticed an increase in the production of 2-phenyl ethanol by
Lt when the medium was supplemented with phenylalanine (its precursor) compared to a medium with an addition of ammonium only [
49]. In a mixed culture
Sc/
Lt under anaerobic conditions compared to a pure culture of
Lt, Shekhawat et al. (2019) [
61] observed an upregulation of genes involved in the metabolism of amino acids, and especially those involved in the conversion of phenylalanine into phenylethanol. This suggested that this upregulation, which probably was linked to interactions, could be lower in the case of aerobic conditions. This could then explain the negative impact that oxygenation seemed to have on phenylethanol production in this study (stand. effect −2.367, with a lower significance:
p-value 0.099).
Temperature had a negative effect on 2-phenylethanol production (stand. effect −9.895). Gobbi et al. (2013) observed no effect on this production [
16]. However, they observed a higher concentration of 2-phenylethanol in the presence of
Lt compared to a pure culture of
Sc under all of the conditions tested (inoculation modalities, temperature of fermentation, different grape juices). They explained that this production was due to synergy between
Sc and
Lt, as both of these yeasts are low producers of this compound. On the other hand, Morales et al. (2019) observed an increase in phenylethanol production in natural must when the
Sc/
Lt ratio decreased in the inoculum from 0.2 to 0.02 [
23]: the results here showed a slight inverse trend (stand. effect 2.350,
p-value 0.1), although it did not seem to be the most impactive factor, accounting for only 2.79% of the observed variance. Both these impacts of temperature and
Sc/
Lt ratio on 2-phenylethanol production may, at first glance, be perceived as the sole result of the impact of temperature and
Sc/
Lt ratio on yeast populations shown in
Section 3.1. However, metabolism modifications via yeast–yeast interactions tend to complexify the phenomenon, as discussed earlier in this study.
The production of isoamyl alcohol decreased when the inoculum ratio favored
Sc (stand. effect −9.407, var. contrib. 83.70%) or when temperature was increased (stand. effect −3.636, var. contrib. 12.50%). A negative impact of the
Sc/
Lt ratio on isoamyl alcohol production was also observed by Morales et al. (2019) [
23]. Temperature had a negative effect on isoamyl alcohol, but Gobbi et al. (2013) did not observe any impact of temperature on the production of this molecule [
16]. Fairbairn et al. (2021) noticed an increase in the production of isoamyl alcohol by
Lt when the medium was supplemented with the amino acid precursors of this alcohol (leucine, isoleucine) compared to a medium with an addition of ammonium only [
49]: such an impact of nitrogen was not observed here. Shekhawat et al. (2017) [
20] observed, in mixed
Sc/
Lt cultures, that oxygenation resulted in an increase in higher alcohols, particularly isoamyl alcohol, 2-phenylethanol, and isobutanol: the absence of an effect of oxygenation observed here can be linked to different conditions of temperature and medium composition (yeast extract as a nitrogen source versus individual amino acids), to the differences in oxygenation levels mentioned previously, or to a strain-dependence factor.
Table 7 shows the impacts of fermentation parameters on the production of various volatile compounds (aldehydes, ketones, and acids). These results are also illustrated in
Figure 5, which presents the standardized effects of each fermentation parameter on this production.
During fermentation in the different conditions tested, aldehydes (acetaldehyde, benzaldehyde) and some ketones were produced. Concerning the global production of aldehydes, none of the four factors tested had a significant effect on their production. As far acetaldehyde is concerned, the
Sc/
Lt ratio had a positive effect on its production according to Morales et al. (2019) [
23], but this was not observed here. The results concerning the production of acetaldehyde in mixed
Sc/
Lt cultures indeed seemed to be variable and strain-dependent [
11]. A slight effect of the
Sc/
Lt ratio on benzaldehyde production (stand. effect −2.424,
p-value 0.094) was nevertheless observed.
Regarding ketones, the production of 6-methyl-5-hepten-2-one was negatively impacted by an increase in temperature (stand. effect −3.838), but not by the other factors. Morales et al. 2019 [
23] showed that this compound was produced more by
Sc than
Lt in pure culture, and that in mixed culture with different
Sc/
Lt ratios, it led to lower quantities than in
Lt culture, leading to the assumption that it may be regulated by an interaction between the two species.
The fermentation of the different synthetic musts in tested conditions led to the production of various organic acids: isovaleric acid, octanoic acid, decanoic acid, and acetic acid. Octanoic acid was not affected by the factors studied. Decanoic acid production was impacted by the
Sc/
Lt ratio as isovaleric acid (stand. effects 3.250 and 6.826,
p-value 0.048 and 0.006, respectively) and also by temperature (stand. effect 3.100,
p-value 0.053). Regarding acetic acid, its final content was managed by temperature and oxygenation (stand. effect 23.103 and 11.375, respectively). Its production also increased when the
Sc/
Lt ratio increased (stand. effect 20.805), supporting previous results [
23,
47].
Lt is indeed known to produce acetic acid at low concentrations, leading to a lower volatile acidity of wine [
2,
14]; however, high strain variability was observed in previous studies [
11]. The positive impact of oxygenation on acetic acid production was unexpected, as this parameter favors the overall presence of
Lt over time.
Lt does not produce as much acetic acid as
Sc [
20,
23]. However, it has also been shown in previous works that oxygenation significantly modulated acetic acid metabolism and increased its production for both species in both pure and mixed cultures. Moreover, this modulation was linked to a decrease in ethanol production [
20], a tendency noted in
Section 3.2 (albeit with low significance), which further corroborated this hypothesis.
Table 8 shows the impacts of fermentation parameters on the production of esters. These results are also illustrated in
Figure 6, which presents the standardized effects of each fermentation parameter on this production.
After fermentation, the media contained different esters derived from different acids and fatty acids: ethyl acetate, isoamyl acetate, ethyl lactate, ethyl hexanoate, octanoate, etc. Although some factors influenced the production of fatty acids such as decanoic acid, no significant effect was observed on esters from these acids. The environmental factors studied significantly impacted only four molecules: ethyl acetate, isoamyl acetate, ethyl hexanoate, and ethyl lactate. Ethyl acetate production increased with an increase in the
Sc/
Lt ratio, oxygenation, and particularly temperature (stand. effects 5.914, 6.856 and 21.058, respectively), but with a decrease in nitrogen content (stand. effect −6.856). These results were in contradiction with previous works [
16,
23] in which an increase in ethyl acetate was observed in conditions favoring the development of
Lt (low temperature, low
Sc/
Lt ratio). However, other studies have also shown that the population dynamics were not the only factor in play here, and that synergy effects could occur as mixed
Sc/
Lt cultures can end up with higher ethyl acetate content than either of the pure ones [
20,
59]. Isoamyl acetate production was negatively impacted by temperature (stand. effect −5.746), while ethyl hexanoate was influenced mostly and positively by the
Sc/
Lt ratio (stand. effect 8.486), and negatively by oxygenation and temperature (stand. effects −4.185 and −4.766, respectively). Ethyl lactate production was negatively influenced by the
Sc/
Lt ratio (stand. effect −4.883). This result should be linked to results concerning lactic acid, since it is a precursor of ethyl lactate. Hranilovic et al. (2021) [
53] showed that sequential inoculation, which allows
Lt to develop better at the beginning of fermentation, led to an increase in ethyl lactate, but these authors also noted that the production of this aroma compound was strain-dependent.
Overall, the production of aroma compounds was impacted by fermentation parameters, but in very different ways depending on both the modulated factor and the volatile molecule studied. Some impacts could be explained by the population dynamics, but interactions between
Lt and
Sc often occurred. These interactions led to metabolism changes and consequently to volatile production, which was different from what would be expected. As far as the production of higher alcohols is concerned, nitrogen content seemed to be a key factor to control. As was shown in
Section 3.1 nitrogen content did not significantly impact the population dynamics; this suggested an interaction between
Sc and
Lt, and more specifically competition for amino acids known to be aroma precursors [
22,
35]. These results then highlighted both the possibility of impacting the final wine volatilome by modulating specific fermentation parameters depending on the goal sought: carboxylic acid production could be modulated by modifying only the inoculum ratio (in a way similar to that of lactic acid, as noted in
Section 3.2), whereas for ketones and esters, temperature seemed to be the most important (and only significant) lever of action. The presence of higher alcohols could be impacted by both of these parameters and, as discussed above, by supplementation with nitrogen, which could offset competition between
Sc and
Lt for specific precursors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





