
Chondroitin Sulfate and Its Derivatives: A Review of Microbial and Other Production Methods

 ,
, 
Abstract
:1. Introduction
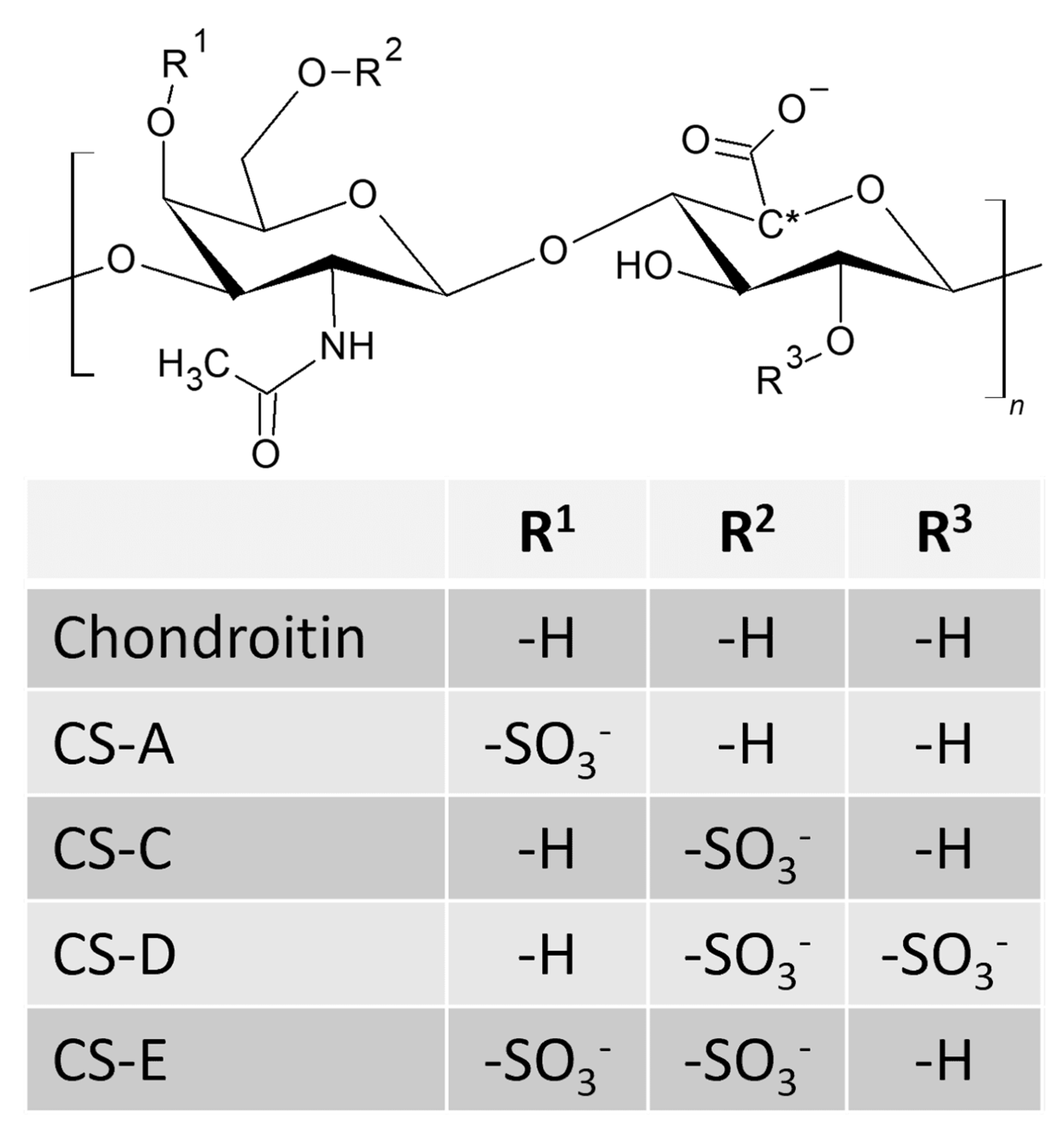
2. Structure and Characteristics of Chondroitin and Chondroitin Sulfates
3. Medical Significance of CS
4. Alternate Methods for CS Production
5. Chemical Synthesis, Chemoenzymatic Synthesis and Bioengineering of CS
6. Current CS Sources and Market
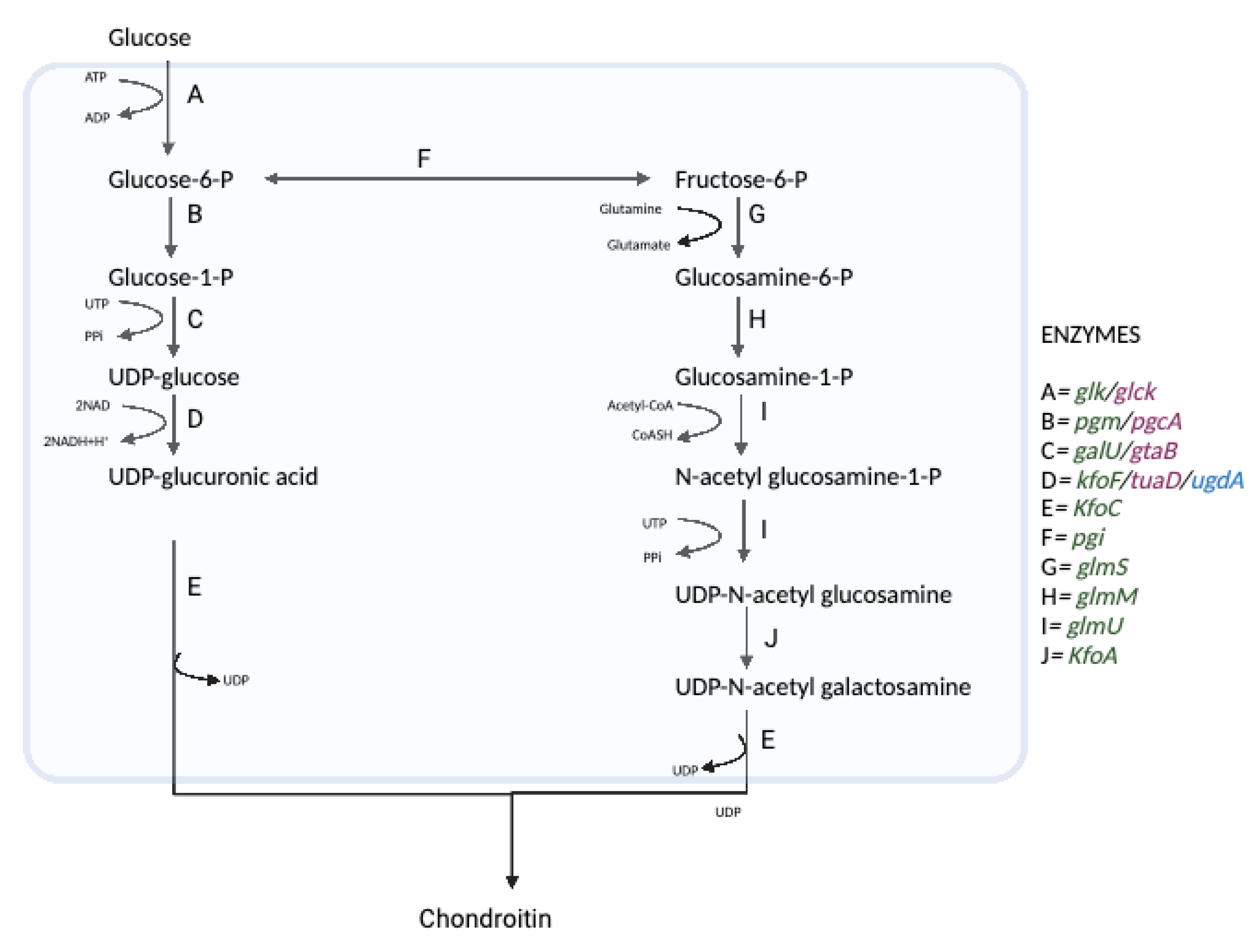
7. Microbial Synthesis of Chondroitin and Chondroitin Sulfate
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Volpi, N. Chondroitin Sulfate Safety and Quality. Molecules 2019, 24, 1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, M.-L.; Jann, B.; Jann, K. Structure and serological characteristics of the capsular K4 antigen of Escherichia coli O5:K4:H4, a fructose-containing polysaccharide with a chondroitin backbone. Eur. J. Biochem. 1988, 177, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Benito-Arenas, R.; Doncel-Pérez, E.; Fernández-Gutiérrez, M.; Garrido, L.; García-Junceda, E.; Revuelta, J.; Fernández-Mayoralas, A. A holistic approach to unravelling chondroitin sulfation: Correlations between surface charge, structure and binding to growth factors. Carbohydr. Polym. 2018, 202, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Doncel-Pérez, E.; Aranaz, I.; Bastida, A.; Revuelta, J.; Camacho, C.; Acosta, N.; Garrido, L.; Civera, C.; García-Junceda, E.; Heras, A.; et al. Synthesis, physicochemical characterization and biological evaluation of chitosan sulfate as heparan sulfate mimics. Carbohydr. Polym. 2018, 191, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Huang, L.; Liu, Y.; Jin, W.; Yao, X.; Rong, J.; Bai, F.; Song, X.; Wang, Z. Strategy for Isolation, Preparation, and Structural Analysis of Chondroitin Sulfate Oligosaccharides from Natural Sources. Anal. Chem. 2020, 92, 11644–11653. [Google Scholar] [CrossRef]
- Da Costa, D.S.; Reis, R.L.; Pashkuleva, I. Sulfation of Glycosaminoglycans and Its Implications in Human Health and Disorders. Annu. Rev. Biomed. Eng. 2017, 19, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Takaoka, T.; Mori, K.; Okimoto, N.; Neya, S.; Hoshino, T. Prediction of the Structure of Complexes Comprised of Proteins and Glycosaminoglycans Using Docking Simulation and Cluster Analysis. J. Chem. Theory Comput. 2007, 3, 2347–2356. [Google Scholar] [CrossRef]
- Tat, S.K.; Pelletier, J.-P.; Mineau, F.; Duval, N.; Martel-Pelletier, J. Variable Effects of 3 Different Chondroitin Sulfate Compounds on Human Osteoarthritic Cartilage/Chondrocytes: Relevance of Purity and Production Process. J. Rheumatol. 2010, 37, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Zhang, C.; Mo, H.; Zhang, H.; Qin, X.; Li, J.; Zhang, Z.; Richel, A. Fabrication of chondroitin sulfate calcium complex and its chondrocyte proliferation in vitro. Carbohydr. Polym. 2020, 254, 117282. [Google Scholar] [CrossRef]
- Li, S.; Ma, F.; Pang, X.; Tang, B.; Lin, L. Synthesis of chondroitin sulfate magnesium for osteoarthritis treatment. Carbohydr. Polym. 2019, 212, 387–394. [Google Scholar] [CrossRef]
- Ma, F.; Liu, N.; Hu, N.; Wen, C.; Tang, B. Synthesis of strontium chondroitin sulfate and the evaluation of its capability to attenuate osteoarthritis. Carbohydr. Polym. 2017, 170, 217–225. [Google Scholar] [CrossRef]
- Bayliss, M.T.; Osborne, D.; Woodhouse, S.; Davidson, C. Sulfation of Chondroitin Sulfate in Human Articular Cartilage. J. Biol. Chem. 1999, 274, 15892–15900. [Google Scholar] [CrossRef] [Green Version]
- Pudełko, A.; Wisowski, G.; Olczyk, K.; Koźma, E.M. The dual role of the glycosaminoglycan chondroitin-6-sulfate in the development, progression and metastasis of cancer. FEBS J. 2019, 286, 1815–1837. [Google Scholar] [CrossRef]
- Da Cunha, A.L.; Aguiar, J.A.K.; da Silva, F.S.C.; Michelacci, Y.M. Do chondroitin sulfates with different structures have different activities on chondrocytes and macrophages? Int. J. Biol. Macromol. 2017, 103, 1019–1031. [Google Scholar] [CrossRef]
- Volpi, N.; Galeotti, F.; Maccari, F.; Capitani, F.; Mantovani, V. Structural definition of terrestrial chondroitin sulfate of various origin and repeatability of the production process. J. Pharm. Biomed. Anal. 2020, 195, 113826. [Google Scholar] [CrossRef]
- Maccari, F.; Ferrarini, F.; Volpi, N. Structural characterization of chondroitin sulfate from sturgeon bone. Carbohydr. Res. 2010, 345, 1575–1580. [Google Scholar] [CrossRef]
- Okamoto, Y.; Higashi, K.; Linhardt, R.J.; Toida, T. Comprehensive analysis of glycosaminoglycans from the edible shellfish. Carbohydr. Polym. 2018, 184, 269–276. [Google Scholar] [CrossRef]
- Rosner, H.; Grimmecke, H.-D.; Knirel, Y.A.; Shashkov, A.S. Hyaluronic acid and a (1→4)-β-d-xylan, extracellular polysaccharides of Pasteurella multocida (Carter type A) strain 880. Carbohydr. Res. 1992, 223, 329–333. [Google Scholar] [CrossRef]
- Bélanger, M.; Burrows, L.L.; Lam, J.S. Functional analysis of genes responsible for the synthesis of the B-band O antigen of Pseudomonas aeruginosa serotype O6 lipopolysaccharide. Microbiology 1999, 145, 3505–3521. [Google Scholar] [CrossRef] [Green Version]
- Bengoechea, J.A.; Pinta, E.; Salminen, T.; Oertelt, C.; Holst, O.; Radziejewska-Lebrecht, J.; Skurnik, M. Functional Characterization of Gne (UDP- N -Acetylglucosamine- 4-Epimerase), Wzz (Chain Length Determinant), and Wzy (O-Antigen Polymerase) of Yersinia enterocolitica Serotype O:8. J. Bacteriol. 2002, 184, 4277–4287. [Google Scholar] [CrossRef] [Green Version]
- Green, D.E.; DeAngelis, P.L. Identification of a chondroitin synthase from an unexpected source, the green sulfur bacterium Chlorobium phaeobacteroides. Glycobiology 2017, 27, cwx008. [Google Scholar] [CrossRef] [Green Version]
- Bhotmange, D.U.; Singhal, R.S. Identification of chondroitin-like molecules from biofilm isolates Exiguobacterium indicum A11 and Lysinibacillus sp. C13. J. Appl. Microbiol. 2015, 119, 1046–1056. [Google Scholar] [CrossRef] [Green Version]
- Djerbal, L.; Lortat-Jacob, H.; Kwok, J. Chondroitin sulfates and their binding molecules in the central nervous system. Glycoconj. J. 2017, 34, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Kastana, P.; Choleva, E.; Poimenidi, E.; Karamanos, N.; Sugahara, K.; Papadimitriou, E. Insight into the role of chondroitin sulfate E in angiogenesis. FEBS J. 2019, 286, 2921–2936. [Google Scholar] [CrossRef] [Green Version]
- Campoli, M.; Ferrone, S.; Wang, X. Functional and Clinical Relevance of Chondroitin Sulfate Proteoglycan 4; Woude, G.F., Klein, G.B., Eds.; Academic Press: Cambridge, MA, USA, 2010; Volume 109, pp. 73–121. [Google Scholar]
- Vallet, S.D.; Clerc, O.; Ricard-Blum, S. Glycosaminoglycan–Protein Interactions: The First Draft of the Glycosaminoglycan Interactome. J. Histochem. Cytochem. 2020, 69, 93–104. [Google Scholar] [CrossRef]
- Sharma, R.; Kuche, K.; Thakor, P.; Bhavana, V.; Srivastava, S.; Mehra, N.K.; Jain, S. Chondroitin Sulfate: Emerging biomaterial for biopharmaceutical purpose and tissue engineering. Carbohydr. Polym. 2022, 286, 119305. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Panina, E.G.; Sanamyan, N.P.; Dmitrenok, A.S.; Tsvetkova, E.A.; Ushakova, N.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure and Anti-Inflammatory Activity of a New Unusual Fucosylated Chondroitin Sulfate from Cucumaria djakonovi. Mar. Drugs 2018, 16, 389. [Google Scholar] [CrossRef] [Green Version]
- Shinmei, M.; Miyauchi, S.; Machida, A.; Miyazaki, K. Quantitation of chondroitin 4-sulfate and chondroitin 6-sulfate in pathologic joint fluid. Arthritis Care Res. 1992, 35, 1304–1308. [Google Scholar] [CrossRef]
- Rnjak-Kovacina, J.; Tang, F.; Whitelock, J.M.; Lord, M.S. Glycosaminoglycan and Proteoglycan-Based Biomaterials: Current Trends and Future Perspectives. Adv. Health Mater. 2017, 7, e1701042. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, L.H.; Ye, S.Z.; Liu, S.J.; Liu, Z.Y.; Wu, C.Y. Lipid-lowering and anti-oxidation effects of chondroitin sulfate prepared from squid cartilage in hypercholesterolemia mice. Int. J. Clin. Exp. Med. 2017, 10, 2230–2240. [Google Scholar]
- Shen, Q.; Zhang, C.; Qin, X.; Zhang, H.; Zhang, Z.; Richel, A. Modulation of gut microbiota by chondroitin sulfate calcium complex during alleviation of osteoporosis in ovariectomized rats. Carbohydr. Polym. 2021, 266, 118099. [Google Scholar] [CrossRef] [PubMed]
- Benito-Arenas, R.; Zárate, S.G.; Revuelta, J.; Bastida, A. Chondroitin Sulfate-Degrading Enzymes as Tools for the Development of New Pharmaceuticals. Catalysts 2019, 9, 322. [Google Scholar] [CrossRef] [Green Version]
- Iwanaga, M.; Kodama, Y.; Muro, T.; Nakagawa, H.; Kurosaki, T.; Sato, K.; Nakamura, T.; Kitahara, T.; Sasaki, H. Biocompatible complex coated with glycosaminoglycan for gene delivery. J. Drug Target. 2017, 25, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Vieira, R.P.; Mulloy, B.; Mourão, P.A. Structure of a fucose-branched chondroitin sulfate from sea cucumber. Evidence for the presence of 3-O-sulfo-beta-D-glucuronosyl residues. J. Biol. Chem. 1991, 266, 13530–13536. [Google Scholar] [CrossRef]
- Higashi, K.; Okamoto, Y.; Mukuno, A.; Wakai, J.; Hosoyama, S.; Linhardt, R.J.; Toida, T. Functional chondroitin sulfate from Enteroctopus dofleini containing a 3-O-sulfo glucuronic acid residue. Carbohydr. Polym. 2015, 134, 557–565. [Google Scholar] [CrossRef]
- Linhardt, R.; Zhang, F. Glycosaminoglycans. In Handbook of Glycomics; 2009; Available online: https://www.sciencedirect.com/science/article/pii/B9780123736000000032 (accessed on 7 July 2022).
- Nakano, T.; Betti, M.; Pietrasik, Z. Extraction, Isolation and Analysis of Chondroitin Sulfate Glycosaminoglycans. Recent Pat. Food Nutr. Agric. 2010, 2, 61–74. [Google Scholar] [CrossRef]
- Silva, L.F. Isolation and purification of chondroitin sulfate. In Chondroitin Sulfate: Structure, Role and Pharmacological Activity; Academic Press: Cambridge, MA, USA, 2006; Volume 53, pp. 21–31. [Google Scholar]
- Mende, M.; Bednarek, C.; Wawryszyn, M.; Sauter, P.; Biskup, M.B.; Schepers, U.; Bräse, S. Chemical Synthesis of Glycosaminoglycans. Chem. Rev. 2016, 116, 8193–8255. [Google Scholar] [CrossRef]
- Li, J.; Su, G.; Liu, J. Enzymatic Synthesis of Homogeneous Chondroitin Sulfate Oligosaccharides. Angew. Chem. Int. Ed. 2017, 56, 11784–11787. [Google Scholar] [CrossRef]
- DeAngelis, P.L.; Gunay, N.S.; Toida, T.; Mao, W.-J.; Linhardt, R.J. Identification of the capsular polysaccharides of Type D and F Pasteurella multocida as unmodified heparin and chondroitin, respectively. Carbohydr. Res. 2002, 337, 1547–1552. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.; Xu, X.; Tan, Y.; Liu, X.-W.; Fang, J. Chemoenzymatic synthesis of homogeneous chondroitin polymers and its derivatives. Carbohydr. Polym. 2019, 232, 115822. [Google Scholar] [CrossRef]
- Vessella, G.; Traboni, S.; Cimini, D.; Iadonisi, A.; Schiraldi, C.; Bedini, E. Development of Semisynthetic, Regioselective Pathways for Accessing the Missing Sulfation Patterns of Chondroitin Sulfate. Biomacromolecules 2019, 20, 3021–3030. [Google Scholar] [CrossRef]
- Klara, J.; Marczak, A.; Łatkiewicz, A.; Horak, W.; Lewandowska-Łańcucka, J. Lysine-functionalized chondroitin sulfate improves the biological properties of collagen/chitosan-based injectable hydrogels. Int. J. Biol. Macromol. 2022, 202, 318–331. [Google Scholar] [CrossRef]
- Yang, S.; Liu, Q.; Zhang, G.; Zhang, X.; Zhao, Z.; Lei, P. An Approach to Synthesize Chondroitin Sulfate-E (CS-E) Oligosaccharide Precursors. J. Org. Chem. 2018, 83, 5897–5908. [Google Scholar] [CrossRef]
- Fuentes, E.P.; Diaz, V.B. Oligosaccharide mapping of chondroitin sulfate obtained from different animal sources. Acta Farm. Bonaer. 1998, 17, 135–142. [Google Scholar]
- Luo, X.; Fosmire, G.; Leach, R. Chicken keel cartilage as a source of chondroitin sulfate. Poult. Sci. 2002, 81, 1086–1089. [Google Scholar] [CrossRef]
- Wang, P.; Tang, J. Solvent-free mechanochemical extraction of chondroitin sulfate from shark cartilage. Chem. Eng. Process. Process Intensif. 2009, 48, 1187–1191. [Google Scholar] [CrossRef]
- Watch, M. Chondroitin Sulfate Market to Witness Size, Share, Trend, Demand, Business Outlook and Growth Acceleration by. 2022. Available online: https://www.marketwatch.com/press-release/chondroitin-sulfate-market-to-witness-size-share-trend-demand-business-outlook-and-growth-acceleration-by-2027-2022-04-27 (accessed on 7 July 2022).
- SBwire. Chondroitin Sulfate Market to Hit 3960 Million US $ by 2025; QY Research, Inc; Available online: http://www.sbwire.com/press-releases/chondroitin-sulfate-market-to-hit-3960-million-us-by-2025-qy-research-inc-1219532.htm (accessed on 7 July 2022).
- Griffin, M.E.; Hsieh-Wilson, L.C. Synthetic probes of glycosaminoglycan function. Curr. Opin. Chem. Biol. 2013, 17, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- DeAngelis, P.L.; Liu, J.; Linhardt, R.J. Chemoenzymatic synthesis of glycosaminoglycans: Re-creating, re-modeling and re-designing nature’s longest or most complex carbohydrate chains. Glycobiology 2013, 23, 764–777. [Google Scholar] [CrossRef] [Green Version]
- Badri, A.; Williams, A.; Awofiranye, A.; Datta, P.; Xia, K.; He, W.; Fraser, K.; Dordick, J.S.; Linhardt, R.J.; Koffas, M.A.G. Complete biosynthesis of a sulfated chondroitin in Escherichia coli. Nat. Commun. 2021, 12, 1389. [Google Scholar] [CrossRef]
- Couto, M.R.; Rodrigues, J.L.; Rodrigues, L.R. Heterologous production of chondroitin. Biotechnol. Rep. 2022, 33, e00710. [Google Scholar] [CrossRef]
- Wu, J.-R.; Chen, P.-Y.; Shien, J.-H.; Shyu, C.-L.; Shieh, H.K.; Chang, F.; Chang, P.-C. Analysis of the biosynthesis genes and chemical components of the capsule of Avibacterium paragallinarum. Veter- Microbiol. 2010, 145, 90–99. [Google Scholar] [CrossRef]
- Liu, J.; Yang, A.; Liu, J.; Ding, X.; Liu, L.; Shi, Z. KfoE encodes a fructosyltransferase involved in capsular polysaccharide biosynthesis in Escherichia coli K4. Biotechnol. Lett. 2014, 36, 1469–1477. [Google Scholar] [CrossRef]
- Whitfield, C. Structure and Assembly of Escherichia coli Capsules. EcoSal Plus 2009, 3, 2. [Google Scholar] [CrossRef]
- He, W.; Fu, L.; Li, G.; Jones, J.A.; Linhardt, R.J.; Koffas, M. Production of chondroitin in metabolically engineered E. coli. Metab. Eng. 2015, 27, 92–100. [Google Scholar] [CrossRef]
- Cimini, D.; Fantaccione, S.; Volpe, F.; De Rosa, M.; Restaino, O.F.; Aquino, G.; Schiraldi, C. IS2-mediated overexpression of kfoC in E. coli K4 increases chondroitin-like capsular polysaccharide production. Appl. Microbiol. Biotechnol. 2014, 98, 3955–3964. [Google Scholar] [CrossRef]
- Manzoni, M.; Bergomi, S.; Molinari, F.; Cavazzoni, V. Production and purification of an extracellularly produced K4 polysaccharide from Escherichia coli. Biotechnol. Lett. 1996, 18, 383–386. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, W.; Wang, Y.; Sheng, J.; Xu, R.; Li, J.; Kang, Z. Biosynthesis of non-animal chondroitin sulfate from methanol using genetically engineered Pichia pastoris. Green Chem. 2021, 23, 4365–4374. [Google Scholar] [CrossRef]
- Cimini, D.; Restaino, O.F.; Catapano, A.; de Rosa, M.; Schiraldi, C. Production of capsular polysaccharide from Escherichia coli K4 for biotechnological applications. Appl. Microbiol. Biotechnol. 2009, 85, 1779–1787. [Google Scholar] [CrossRef]
- Schiraldi, C.; Alfano, A.; Cimini, D.; de Rosa, M.; Panariello, A.; Restaino, O.F. Application of a 22L scale membrane bioreactor and cross-flow ultrafiltration to obtain purified chondroitin. Biotechnol. Prog. 2012, 28, 1012–1018. [Google Scholar] [CrossRef]
- Cimini, D.; de Rosa, M.; Carlino, E.; Ruggiero, A.; Schiraldi, C. Homologous overexpression of rfaH in E. coli K4 improves the production of chondroitin-like capsular polysaccharide. Microb. Cell Factories 2013, 12, 46. [Google Scholar] [CrossRef] [Green Version]
- D’Ambrosio, S.; Alfano, A.; Cassese, E.; Restaino, O.F.; Ferraiuolo, S.B.; Finamore, R.; Cammarota, M.; Schiraldi, C.; Cimini, D. Production and purification of higher molecular weight chondroitin by metabolically engineered Escherichia coli K4 strains. Sci. Rep. 2020, 10, 13200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, Q.; Huang, H.; Wang, H.; Wang, Y.; Du, G.; Chen, J.; Kang, Z. A microbial-enzymatic strategy for producing chondroitin sulfate glycosaminoglycans. Biotechnol. Bioeng. 2018, 115, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Zhang, L.; Yuan, P.; Kang, Z.; Du, G.; Chen, J. Efficient biosynthesis of polysaccharides chondroitin and heparosan by metabolically engineered Bacillus subtilis. Carbohydr. Polym. 2016, 140, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Luozhong, S.; Yu, H.; Guo, Z. Biosynthesis of Chondroitin in Engineered Corynebacterium glutamicum. J. Microbiol. Biotechnol. 2019, 29, 392–400. [Google Scholar] [CrossRef]
- He, W.; Zhu, Y.; Shirke, A.; Sun, X.; Liu, J.; Gross, R.A.; Koffas, M.A.G.; Linhardt, R.J.; Li, M. Expression of chondroitin-4-O-sulfotransferase in Escherichia coli and Pichia pastoris. Appl. Microbiol. Biotechnol. 2017, 101, 6919–6928. [Google Scholar] [CrossRef]
- Badri, A.; Williams, A.; Xia, K.; Linhardt, R.J.; Koffas, M.A.G. Increased 3′-Phosphoadenosine-5′-phosphosulfate Levels in Engineered Escherichia coli Cell Lysate Facilitate the In Vitro Synthesis of Chondroitin Sulfate A. Biotechnol. J. 2019, 14, 1800436. [Google Scholar] [CrossRef]
- Awofiranye, A.E.; Baytas, S.N.; Xia, K.; Badri, A.; He, W.; Varki, A.; Koffas, M.; Linhardt, R.J. N-glycolyl chondroitin synthesis using metabolically engineered E. coli. AMB Express 2020, 10, 144. [Google Scholar] [CrossRef]
- Schiraldi, C.; Cimini, D.; de Rosa, M. Production of chondroitin sulfate and chondroitin. Appl. Microbiol. Biotechnol. 2010, 87, 1209–1220. [Google Scholar] [CrossRef]
- Sundaresan, G.; Abraham, R.J.J.; Rao, V.A.; Babu, R.N.; Govind, V.; Meti, M.F. Established method of chondroitin sulphate extraction from buffalo (Bubalus bubalis) cartilages and its identification by FTIR. J. Food Sci. Technol. 2018, 55, 3439–3445. [Google Scholar] [CrossRef] [Green Version]
- Sevimli, R.; Erenler, A.S.; Karabulut, A.B.; Akpolat, N.; Geçkil, H. Microbial chondroitin sulfate in experimental knee osteoarthritis model investigation of chondroprotective effect. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 5402–5411. [Google Scholar]
- Bergfeld, A.K.; Lawrence, R.; Diaz, S.L.; Pearce, O.M.T.; Ghaderi, D.; Gagneux, P.; Leakey, M.G.; Varki, A. N-glycolyl groups of nonhuman chondroitin sulfates survive in ancient fossils. Proc. Natl. Acad. Sci. USA 2017, 114, E8155–E8164. [Google Scholar] [CrossRef] [Green Version]
- Leroux, M.; Michaud, J.; Bayma, E.; Armand, S.; Drouillard, S.; Priem, B. Misincorporation of Galactose by Chondroitin Synthase of Escherichia coli K4: From Traces to Synthesis of Chondbiuronan, a Novel Chondroitin-Like Polysaccharide. Biomolecules 2020, 10, 1667. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | Strain | Gag Type | Yield | Reference |
|---|---|---|---|---|
| 1. | E. coli K4 | Unsulfated chondroitin | 200 mg/L | [61] |
| 2. | E. coli K4 | Unsulfated chondroitin | 1.4 g/L | [63] |
| 3. | E. coli K4 | Unsulfated chondroitin | 1.74 g/L | [57] |
| 4. | E. coli K4 | Unsulfated chondroitin | 5.3 g/L | [65] |
| 5. | E. coli K4 | Unsulfated chondroitin | - | [66] |
| 6. | E. coli K4 | CS-A | - | [54] |
| 7. | E. coli K4 | Fructosylated chondroitin | 4.5g/L | [64] |
| 8. | E. coli K4 | N-glycolyl chondroitin | - | [72] |
| 9. | E. coli K-12 MG1655 | CS-A | ~27 μg/g DCW with about 96% sulfation | [54] |
| 10. | E. coli BL21 Star™ (DE3) | Unsulfated chondroitin | 2.4 g/L | [59] |
| 11. | B. subtilis natto | Chondroitin sulfate | 237.7 mg/L | [73] |
| 12. | B. subtilis | Unsulfated chondroitin | 5.22 g/L | [68] |
| 13. | B. subtilis | Unsulfated chondroitin | 7.15 g/L | [67] |
| 14. | C. glutamicum | Unsulfated chondroitin | 1.91 g/L | [69] |
| 15. | P. multocida | Unsulfated chondroitin | - | [42] |
| 16. | P. pastoris | CS-A | 2.1 g/L with 4.0% sulfation | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Awofiranye, A.E.; Hudson, J.; Tithi, A.D.; Linhardt, R.J.; Vongsangnak, W.; Koffas, M.A.G. Chondroitin Sulfate and Its Derivatives: A Review of Microbial and Other Production Methods. Fermentation 2022, 8, 323. https://doi.org/10.3390/fermentation8070323
Awofiranye AE, Hudson J, Tithi AD, Linhardt RJ, Vongsangnak W, Koffas MAG. Chondroitin Sulfate and Its Derivatives: A Review of Microbial and Other Production Methods. Fermentation. 2022; 8(7):323. https://doi.org/10.3390/fermentation8070323
Chicago/Turabian StyleAwofiranye, Adeola E., Jon Hudson, Aditi Dey Tithi, Robert J. Linhardt, Wanwipa Vongsangnak, and Mattheos A. G. Koffas. 2022. "Chondroitin Sulfate and Its Derivatives: A Review of Microbial and Other Production Methods" Fermentation 8, no. 7: 323. https://doi.org/10.3390/fermentation8070323
APA StyleAwofiranye, A. E., Hudson, J., Tithi, A. D., Linhardt, R. J., Vongsangnak, W., & Koffas, M. A. G. (2022). Chondroitin Sulfate and Its Derivatives: A Review of Microbial and Other Production Methods. Fermentation, 8(7), 323. https://doi.org/10.3390/fermentation8070323