Abstract
In this experiment, B. subtilis was added to pelletized straw-based total mixed ration, and the rumen microbial diversity of house-fed Duhan hybrid sheep was compared. Ten 3-month-old weaned Duhan hybrid lambs were separated into two groups and fattened for 80 days using a single-factor trial design. During the fattening period, the control and the experiment groups were fed with the same ration, except that the experiment group was supplemented with B. subtilis. The results showed that the addition of B. subtilis could significantly increase the daily weight gain, total weight gain, rumen microbial abundance, and rumen microbial diversity of the Duhan lamb. Among them, the proportion of microbial flora such as Bacteroidetes was significantly increased, producing more acetate, iso-butyrate, and butyrate, obtaining higher energy efficiency.
1. Introduction
The use of natural alternatives in the livestock industry is rapidly increasing due to the global ban of the use of in-feed antibiotics as growth promoting agents, which include probiotics, prebiotics, enzymes, and plant nutraceuticals [1,2,3]. Direct-fed microbials (DFM) are products that contain live (viable) probiotic microorganisms (bacteria and fungi) with the primary goal of enhancing animal health and improving production in ruminants, and therefore has led to increasing interest in the animal industry. However, the efficacy of DFM in promoting animal health and production performance varies greatly owning to the different microbial species compositions of the DFM products, animal species and diets, and feeding conditions [4,5,6].
Bacterial DFM for ruminants mostly consist of propionate-forming bacteria and lactic acid-producing bacteria from a range of genera. Recently, interest in the use of Bacillus species as DFM in ruminants has been increasing owing to their specific properties, e.g., heat stability and ability to survive the low pH of the digestive tract. Bacillus subtilis (BS) is one of the species from genus Bacillus with strong environmental adaptability [7] that produces polymyxin, nystatin, gramicidin, and other active substances such as enzymes (alpha-amylase, protease, lipase, cellulase, etc.) and vitamins during growth. These active substances possess strong inhibitory effects on pathogens and the subsequent infection as well as favorably modulating nutrient metabolism and immune function [8,9]. B. subtilis rapidly consumes free oxygen in the intestine, resulting in hypoxia, which promotes the growth of beneficial anaerobic bacteria while preventing the growth of harmful bacteria. Therefore, BS could be a promising DFM for ruminants. Several strains of BS (e.g., BS natto, BS PB6, BS 10071, and BS C-3102) have been assessed in cattle and in vitro for their effects on rumen fermentation and productive performance [10,11,12,13,14,15]. However, there is scarce information about the effects of BS on the rumen microbiome that is crucial for elucidating the mechanism by which BS improves rumen metabolism, animal health, and productive performance.
The objectives of this study were to evaluate the effects of the supplementation of BS on the rumen metabolism and growth performance of lambs and to determine its effects on the rumen bacterial microbiome.
2. Materials and Methods
2.1. Experimental Design, Animals, and Diet Preparation
Ten (five male and five female) lambs (3 months old) with an initial body weight (BW) of 24 ± 1.01 kg were randomly divided into two groups stratified by gender, and were randomly allocated to two dietary treatments. The treatments were basal diet only (Control; C) and basal diet supplemented with B. subtilis C-3102, B. subtilis at the concentration of 300 g/ton; 3 × 108 CFU/kg (experimental group; BS) [16]. The basal diet was a corn, corn stalk, and DDG-based total mixed ration (TMR) that was formulated to meet the nutrient requirements of growing sheep (NRC; 2012) for two growing stages (Table 1). The ingredients of the TMR were mixed thoroughly, tempered at 90 °C for 45 s, and then pelletized at 60 °C to an average size of 0.5 × 5 cm pellets using a YPM508E Granulator (Jiangsu Yongli Machinery Co., Ltd., Liyang City, Jiangsu Province, China). The B. subtilis product was obtained from Calpis Trading Co., Ltd. of Japan Asahi Group (Shanghai, China) that contained minimum 1 × 109 CFU/g of viable cells and was mixed with other dietary ingredients prior to pelleting. All diets were made in one batch at the beginning of the experiment and stored in covered containers for the entire experiment period.

Table 1.
Ingredients and nutrient composition of the diet fed to the lambs.
2.2. Experimental Procedure and Sampling
A total of 87 d feeding experiment was conducted with a 7 d adaptation followed by an 80 d data collection period. The lambs were individually fed twice daily for ad libitum intake throughout the adaptation and experimental periods. The lambs were fed the Stage 1 diet for the first 35 d followed by the Stage 2 diet for the remaining experimental period. The orts were weighed weekly for measuring dry matter intake (DMI). The lambs were weighed twice after overnight fast at the beginning and at the end of the experiment, and weekly between. The animals had free access to water during the entire experimental period. The animal care protocol was approved by the Animal Care Committee, Inner Mongolia Autonomous Region Academy of Agriculture and Animal Husbandry, Hohhot, Inner Mongolia, China.
At the end of the experiment (d 87), all lambs were slaughtered after being fasted for 18 h using the procedure described by Geng et al. [12]. The carcass characteristics including carcass weight, carcass ratio, and weights of the heart, liver, lung, spleen, kidney, rumen, reticulum, ovum, true stomach, and intestines were determined using the procedures described by Geng et al. [17].
The rumen was opened immediately upon removal and the rumen content was strained through four layers of cheesecloth to obtain the rumen fluid. The rumen fluid from each lamb was divided into two portions, one being stored at −20 °C in 50 mL centrifuge tubes for the determination of the total volatile fatty acids (VFA), microbial protein, and ammonia nitrogen and the other being stored at −80 °C in 5 mL cryopreservation tubes for DNA extraction and high-throughput sequencing analysis.
2.3. Laboratory Analyses
2.3.1. Determinations of Rumen Ammonia Nitrogen (NH3-N), VFA, and Microbial Protein
The rumen fluid samples were processed and analyzed for NH3-N as described by Feng et al. [18], microbial protein using procedures described by Bradford [19], and for VFA using gas chromatography [20].
2.3.2. Determination of Microbial Compositions by Sequencing
Genomic DNA was extracted from the rumen fluid samples using the CTAB method described by Rogers et al. [21]. The purity and concentration of the extracted DNA were determined by agarose gel electrophoresis, as described by Lee et al. [22]. The extracted rumen microbial DNA was diluted to 1 ng/μL and the genome sequencing was conducted by Nuohe Zhiyuan Technology Co., Ltd. (Beijing, China). Then, 16S rRNA genes of the distinct regions V3-V4 were amplified using a specific primer (341F 5′-CCTAYGGGRBGCASCAG-3′; 806R5′-GGACTACNNGGGTATCTAAT--3′) with the barcode. All PCR reactions were carried out in a 30 μL reaction system with 15 μL of Phusion® High-Fidelity PCR Master Mix (New England Biolabs), 0.2 μmol of forward and reverse primers, and 10 ng of DNA. The thermal cycling consisted of initial denaturation at 98 °C for 1 min, followed by 30 cycles of denaturation at 98 °C for 10 s, annealing at 50 °C for 30 s, and elongation at 72 °C for 30 s.
The PCR product obtained from the above procedure was then mixed with the same amount of buffer containing SYB green and was electrophoresed on 2% agarose gel and purified with a Gene JETTM Gel Extraction Kit (Thermo Scientific, Waltham, MA, USA).
Sequencing libraries were generated using an Ion Plus Fragment Library Kit 48 rxns (Thermo Scientific, Waltham, MA, USA) following the manufacturer’s recommendations. The library quality was assessed on the Qubit@ 2.0 Fluorometer (Thermo Scientific). The library was sequenced on an Ion S5 TM XL platform and 400 bp/600 bp single-end reads were generated.
2.4. Calculation and Statistical Analysis
The operational taxonomic unit (OTU) database single-end reads were assigned to samples based on their unique barcode and truncated by cutting off the barcode and primer sequence. Quality filtering on the raw reads was performed under specific filtering conditions to obtain the high-quality clean reads according to the Cutadapt quality control process [23]. The reads were compared with the reference database using the UCHIME algorithm (UCHIME Algorithm, http://www.drive5.com/usearch/manual/uchime_algo.html accessed on 20 December 2022) [24] to identify chimera sequences. The clean reads were obtained after the removal of the chimera sequences [25]. Sequence analysis was performed by Uparse software (Uparse v7.0.1001, http://drive5.com/uparse/ accessed on 20 December 2022) [26]. Sequences with ≥97% similarity were assigned to the same OTUs. Representative sequences from each OTU were screened by Chao1, observed species, and Shannon’s index to obtain α diversity. QIIME was employed to calculate both weighted and unweighted UniFrac to obtain beta diversity. Unweighted UniFrac was used for principal coordinate analysis (PCoA).
The data were analyzed by analysis of variance using the PROC MIXED procedure of SAS with the individual animal as a statistical unit. Differences were determined using the PDIFF option in SAS 9.4. Significance was declared at p < 0.05 and the tendency of significant differences was described at p values between 0.05 and 0.1.
3. Results
3.1. Growth Performance and Carcass Characteristics
Lambs had similar BW (kg) at the beginning of the experiment (25.3 ± 2.07 kg vs. 25.9 ± 2.11 kg; p = 0.140). Dietary supplementation of B. subtilis had no effects on DMI (1.52 ± 0.15 kg vs. 1.53 ± 0.20 kg; p = 0.830), but increased average daily gain (255.8 ± 56.22 g vs. 284.3 ± 58.78 g; p < 0.02) of the lambs, resulting in significantly heavier (48.7 ± 5.82 vs. 45.7 ± 5.53 kg; p < 0.05) lambs in the BS than in the C group at the end of the experiment.
All lambs had similar carcass weight, carcass ratio, and weights of heart, liver, lung, spleen, kidney, rumen, reticulum, ovum, true stomach, and small intestine (p > 0.05). However, the large intestine was significantly heavier (p < 0.05) for the BS (265.1 ± 11.12 g)-supplemented group than for the C group (224.9 ± 32.61 g) lambs. The total weight of the digestive tract of the lambs in the BS group (2.4 ± 0.14 kg) was also significantly higher (p < 0.05) than that in the C group (2.2 ± 0.16 kg).
3.2. Rumen Metabolites
The dietary supplementation of B. subtilis did not affect (p > 0.05) ruminal pH or concentrations of NH3-N and total VFA, but increased (p ≤ 0.05) the concentration of microbial protein (Table 1 and Table 2). The concentrations of acetate, iso-butyrate, and butyrate were higher (p < 0.01), but that of propionate was lower (p < 0.01) for the BS group than for the C group, resulting in a higher ratio of acetate to propionate (p < 0.01) for BS than for C lambs.
Table 2.
pH and concentrations (mean ± SE; n = 5) of ammonia-N (NH3-N), volatile fatty acids (VFA), and microbial protein in the rumen fluid of lambs fed basal diet only (Control; C) or basal diet supplemented with Bacillus subtilis at the concentration of 3 × 108 CFU/kg DM (BS).
3.3. Bacterial Microbiome of the Rumen Fluid
Effects of B. Subtilis on Rumen Microbiota
An average of 56,934 clean read sequences with an average length of 417 bp were obtained from all 10 rumen fluid samples, which sufficiently covered the bacterial communities in these samples (Figure 1). There were 17 phyla and 27 genera (eight unidentified families) among these samples. Overall, Firmicutes was the most dominant phylum for the rumen fluid from both the BS group (49.9%) and the C group (57.7%), and at the genus level, Succiniclasticum was relatively the most abundant genus, although on average, 86.1% of sequences could not be classified to a particular genus.
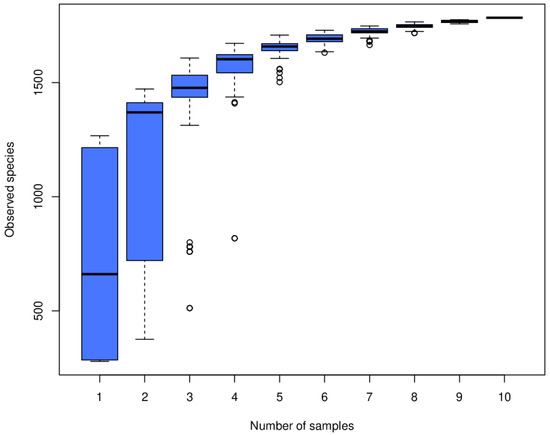
Figure 1.
Horizontal axis is the sample size; vertical axis is the number of OTUs after sampling.
The alpha diversity analysis showed that the OTUs, Chao1, ACE, PD whole tree, and Shannon indices of BS group were significantly higher (p < 0.01) than that of the C group (Table 1 and Table 3).
Table 3.
Alpha indices (mean ± SE; n = 5) of observed species, Shannon, Chao1, ACE, and PD whole tree in the rumen fluid of lambs fed basal diet only (Control; C) or basal diet supplemented with Bacillus subtilis at the concentration of 3 × 108 CFU/kg DM (BS).
The Venn diagram (Figure 2) revealed significant differences in the OTUs between the C group and the BS group. The two groups shared 660 OTUs, but the C group had 184, whilst the BS group had 862 unique OTUs, respectively.
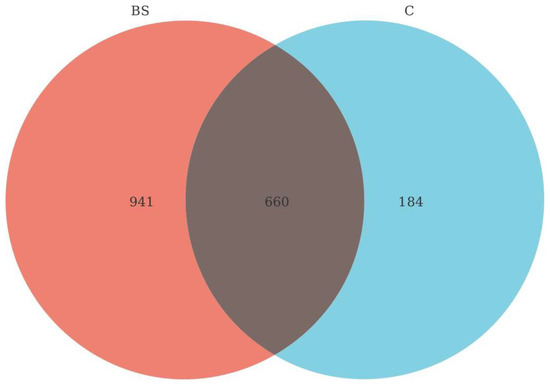
Figure 2.
Venn diagram of OTU distribution in GIT. Each petal in the diagram represents a sample, and different colors represent different samples. The core number in the middle represents the total number of mutual OTUs in all samples, and the number on the Venn represents the number of unique OTUs in the sample.
At the phylum level, the abundances of Firmicutes, Actinobacteria, Oxyphotobacteria, and Proteobacteria in the BS group were significantly lower than the C group (p < 0.05), whereas the abundances of Bacteroidetes and Tenericutes were higher (p < 0.05) for the BS group than for the C group of sheep. Other taxonomies less than 1% and those not annotated only accounted for 1.6% and 2.2% for the C and BS groups, respectively (Table 1 and Table 4). The heat map of the phylum-level classification also showed the same trend (Figure 3). Firmicutes and Bacteroidetes were the most abundant phylum in both group of sheep, although the abundance of microorganisms in each phylum varied between the two groups (Figure 4). The total abundances of Firmicutes and Bacteroidetes were 94.9% in the BS group and 84.6% in the C group. Interestingly, the Firmicutes/Bacteroidetes (F/B) ratios were 1.11 and 2.14 for the BS and C groups, respectively.
Table 4.
Abundance (mean ± SE; n = 5; >1%) at phylum level of bacteria in the rumen fluid of lambs fed basal diet only (Control; C) or basal diet supplemented with Bacillus subtilis at the concentration of 3 × 108 CFU/kg DM (BS).
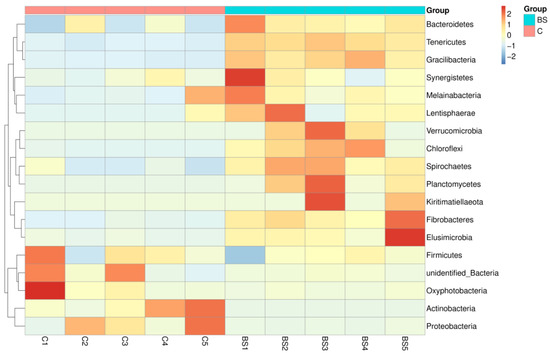
Figure 3.
Analysis of rumen bacterial populations at the phylum classification level.
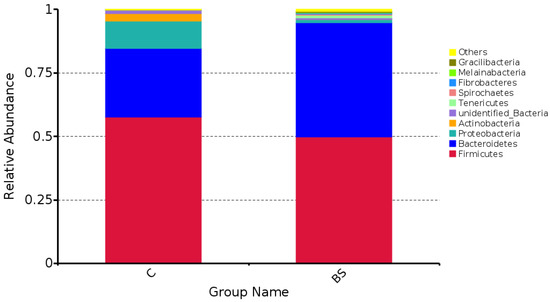
Figure 4.
Species relative abundance column chart at the phylum level.
At the genus level, the abundances of unidentified Lachnospiraceae, Rikenellaceae and Bacteroidales, and Saccharofermentans were higher (p < 0.05), but the abundances of Succiniclasticum, Succinivibrio, Dialister, Syntrophococcus, and Shuttleworthia were lower (p < 0.05) in the rumen fluid of the BS lambs than that of the C group lambs. The abundances of Ruminococcaceae, Pseudoscardovia, Roseburia, Selenomonas, and unidentified Prevotellaceae and Clostridiales were similar between the two groups (Table 1 and Table 5). The taxonomy accounting for less than 1% of the abundance with no annotation was higher (p < 0.05) for the BS than for the C group of lambs. The heat map of the genus-level classification also showed the same trend (Figure 5).
Table 5.
Abundance (mean ± SE; n = 5; >1%) at genus level of bacteria in the rumen fluid of lambs fed basal diet only (Control; C) or basal diet supplemented with Bacillus subtilis at the concentration of 3 × 108 CFU/kg DM (BS).
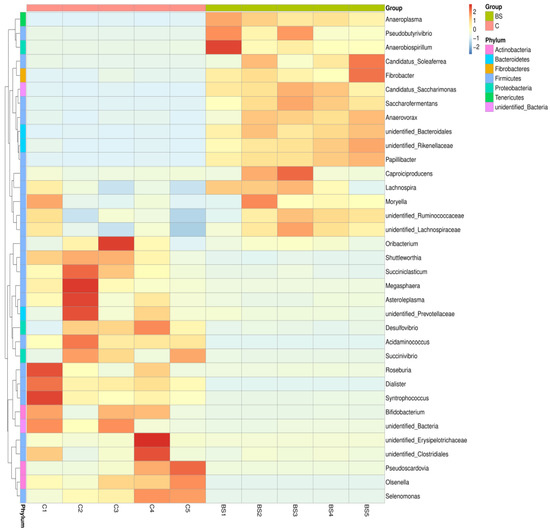
Figure 5.
Heat map of genus-level classification.
The evolutionary cladogram revealed that the species playing an important role were Rikenellaceae, Bacteroidales, Bacteroidia, Bacteroidetes, Christensenellaceae, and Ruminococcaceae in the BS group, whereas these were Bifidobacteriaceae, Bifidobacteriales, Actinobacter, Acidaminococcaceae, Veillonellaceae, Selenomonadales, Negativicutes, and Lachnospiraceae for the C group (Figure 6). The other taxonomies that accounted for less than 1% in both groups were excluded.
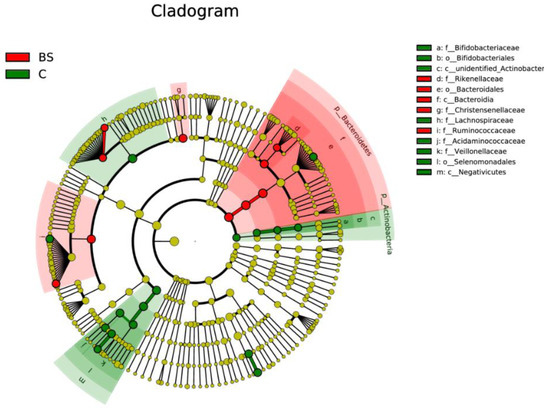
Figure 6.
Taxonomic cladogram reporting the different taxon abundances among GIT groups.
4. Discussion
4.1. Effects of BS Supplementation on Ruminal Fermentation
Although feed intake was not affected, the ADG and final body weight were increased by B. subtilis, indicating that the supplementation of B. subtilis increased the growth rate and the feed efficiency of the lambs in this study. It needs to be pointed out that only five lambs were used in each group because this study focused on the effects of treatment on the rumen microbiome. The animal replicate number was relatively small compared with a typical growth performance study. Therefore, this part of the results was only indicative, and a large-scale feeding experiment needs to be conducted to confirm the findings of this study. Nevertheless, our previous study also showed that supplementation of the same B. subtilis product promoted the growth and development and increased the feed intake of Duhan hybrid sheep [27]. The higher growth rate of the lambs supplemented with B. subtilis might due to the greater microbial protein synthesis, as indicated by its higher microbial protein concentration in the rumen than that of the C group lambs. This is also consistent with the microbiome analysis results that showed that bacterial populations, diversity, and abundance were all increased by B. subtilis supplementation. Although the total VFA concentration was not affected, the lambs supplemented with B. subtilis had higher concentrations of acetate and butyrate than those in the C group. This may indicate that B. subtilis increased the fiber digestion because it is known that the digestion of fiber is prone to produce acetate and butyrate [28]. It has been shown that short-chain fatty acids are an important source of energy for ruminants, with up to 70% of the energy requirements of adult animals being absorbed as short-chain fatty acids through the stratified squamous epithelium of the rumen [29]. It has been shown that at the same nutritional level, feeding B. subtilis could improve energy efficiency and increase the butyric acid content [30], which is consistent with the observations in this study.
4.2. Effects of BS Supplementation on Bacterial Diversity
The bacterial microbiome analysis showed that the OTU number in the BS group was more than two times higher (p < 0.01) than that in the C group, demonstrating that more bacterial species were observed in the BS group lambs than the C group lambs. Higher Shannon index and PD whole tree values were observed for the B. subtilis-supplemented lambs (BS group) than for the C group lambs. Shannon and PD whole tree are positively correlated with bacterial diversity, demonstrating that the supplementation of B. subtilis under the conditions of this study increased the bacterial phylogenetic diversity. Increased bacterial diversity by BS supplementation was also observed for cattle and broilers [31]. It has been shown that B. subtilis could produce a variety of secondary metabolites that inhibit and kill pathogenic bacteria such as Staphylococcus aureus, Candida albicans, Listeria monocytogenes, Escherichia coli, and Enterococcus when it was supplemented to sheep and lambs [32]. This suggested that the supplementation of B. subtilis has the potential to promote intestinal health via the modification of the microbiome in the digestive tract [33]. Research has shown that rumen microbial diversity is positively correlated with the resistance of animals to environmental influences [34]. Therefore, the increased bacterial diversity achieved by BS supplementation may have partially contributed to the enhanced growth performance of the lambs observed in this study. Zhang et al. also reported a similar relationship between rumen bacterial diversity and animal growth performance in cattle [35].
4.3. Effects of BS Supplementation on Bacterial Abundances
Chao1 and ACE are positively correlated with bacterial abundances. The higher values of Chao1 and ACE observed for the B. subtilis-supplemented lambs than for the C group lambs indicated that the supplementation of B. subtilis increased rumen bacterial abundance under the conditions of this study. This study showed that Firmicutes and Bacteroidetes were the two most dominant phyla regardless of the treatment, which is consistent with the observations of other studies [35], indicating that these two phyla play a key role in rumen metabolism. It has been suggested that the F/B ratio in the gut impacts intestinal homeostasis and the energy metabolism of the body [36]. This study found that the supplementation of BS decreased the F/B ratio mainly due to its effect on increasing the abundance of Bacteroidetes. A literature review showed that the dietary supplementation of probiotics affected the F/B ratio depending on the specific probiotics [37]. The phylum Bacteroidetes includes approximately 7000 different species of Gram-negative bacteria that are predominantly from the genera Bacteroides, Alistipes, Parabacteroides, and Prevotella. Bacteroidetes express a relatively large number of genes encoding carbohydrate active enzymes, thus promoting the breakdown of rumen structural polysaccharides. They can also ferment amino acids into acetate [35]. These are consistent with the increased acetate proportion by BS supplementation in this study.
Although the decrease of the abundances of the phyla Proteobacteria and Actinobacteria by BS supplementation did not reach statistical difference, the large scale of the reduction and its potentially biological implications still need to be noted. Further analysis showed that the genera Succinivibrio and Pseudoscardovia accounted for more than 70% of the genera in the phyla Proteobacteria and Actinobacteria for the C group of lambs, whereas it decreased to less than 1% for the BS group of lambs, indicating that the negative effect of BS supplementation on the phyla Proteobacteria and Actinobacteria was mainly due to its effects on decreasing the abundance of the genera Succinivibrio and Pseudoscardovia, respectively. Both Succinivibrio and Pseudoscardovia are opportunistic pathogens possessing health implications to animals [35,38]. Tenericutes were significantly higher (p < 0.05) for the BS group than for the C group. It is one of the inhabitants of the gastrointestinal tract; many studies have focused on pathogenic species, but recent studies found that Tenericutes expressed more carbon metabolism genes. Sugars such as xylose, galactose, and fructose might be fermented to lactate, formate, and acetate. These are consistent with the increased acetate proportion by BS supplementation in this study [39].
Within the phylum Firmicutes, the abundances of unidentified Ruminococcaceae, unidentified Lachnospiraceae, and Succiniclasticum each accounted for more than 1% of the total bacterial populations. In both groups, the abundances of Dialister, unidentified Clostridiales, Syntrophococcus, Roseburia, Selenomonas, and Shuttleworthia each exceeded 1% of the total bacterial populations for the C group of lambs only. On the contrary, the abundance of Saccharofermentans was greater than 1% in the BS group only. This study found that the supplementation of BS increased the abundances of unidentified Lachnospiraceae, unidentified Lachnospiraceae, and Saccharofermentans, but decreased the abundances of Dialister, unidentified Clostridiales, Syntrophococcus, Roseburia, Selenomonas, and Shuttleworthia. Unidentified Ruminococcaceae and unidentified Lachnospiraceae belong to the genera Ruminococcaceae and Lachnospiraceae, respectively, that have been shown to be positively correlated to gut health [40,41] and Saccharofermentans was reported to be positively correlated with feed efficiency [40]. In contrast, Syntrophococcus, Shuttleworthia, unidentified Clostridiales, and Roseburia have been suggested to be positively correlated with diseases. Therefore, the increased abundances of the beneficial bacteria that promote animal health and feed efficiency and decrease the abundances of those disease-causing bacteria by BS supplementation would have promoted the overall health status and production efficiency of the lambs, thereby contributing to the enhanced growth performance of the BS-supplemented lambs in this study.
Within the phylum Bacteroidetes, BS supplementation increased the abundances of unidentified Rikenellaceae and unidentified Bacteroidales. Unidentified Rikenellaceae belongs to Rikenellaceae that has been shown to possess immune-improving effects [42]. Unidentified Bacteroidales belongs to Bacteroidales that possess unique and powerful carbohydrate-utilization systems, and Bacteroidales species are considered potential probiotics [43].
5. Conclusions
The supplementation of BS at the dietary concentration of 3 × 108 CFU/kg enhanced the lamb growth rate and feed efficiency via increasing ruminal microbial protein synthesis and VFA, and favorably modulating the rumen microbiota under the conditions of this study. Both rumen bacterial diversity and the abundances of microflora that are positively related to rumen metabolism and health condition (probiotic populations) were increased, whereas the abundances of those bacteria that are negatively correlated with health conditions were decreased by the BS supplementation. Overall, this study showed that B. subtilis has potential as a direct-fed microbial to enhance the sheep production efficiency and requires further evaluation under the commercial production situation.
Author Contributions
Data curation, F.T., J.L. and Y.W.; Writing—original draft, Y.G. and W.W.; Writing—review & editing, J.Q.; Funding acquisition, S.X. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the China Agriculture Research System of MOF and MARA (CARS-02-55).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, Y.; Nan, X.; Zhao, Y.; Jiang, L.; Wang, H.; Zhang, F.; Hua, D.; Liu, J.; Yao, J.; Yang, L.; et al. Dietary Supplementation of Inulin Ameliorates Subclinical Mastitis via Regulation of Rumen Microbial Community and Metabolites in Dairy Cows. Microbiol. Spectr. 2021, 9, e0010521. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Guo, B.; Yang, M.; Luo, J.; Hu, Y.; Qu, M.; Song, X. Response of Growth Performance, Blood Biochemistry Indices, and Rumen Bacterial Diversity in Lambs to Diets Containing Supplemental Probiotics and Chinese Medicine Polysaccharides. Front. Vet. Sci. 2021, 8, 681389. [Google Scholar] [CrossRef] [PubMed]
- Zeitz, J.O.; Guertler, P.; Pfaffl, M.W.; Eisenreich, R.; Wiedemann, S.; Schwarz, F.J. Effect of non-starch-polysaccharide-degrading enzymes as feed additive on the rumen bacterial population in non-lactating cows quantified by real-time PCR. J. Anim. Physiol. Anim. Nutr. 2013, 97, 1104–1113. [Google Scholar] [CrossRef]
- Maake, T.W.; Aiyegoro, O.A.; Adeleke, M.A. Effects of Lactobacillus rhamnosus and Enterococcus faecalis Supplementation as Direct-Fed Microbials on Rumen Microbiota of Boer and Speckled Goat Breeds. Vet. Sci. 2021, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.F.; Lelis, A.L.J.; Fan, P.; Calvo Agustinho, B.; Lobo, R.R.; Arce-Cordero, J.A.; Dai, X.; Jeong, K.C.; Faciola, A.P. Effects of lactic acid-producing bacteria as direct-fed microbials on the ruminal microbiome. J. Dairy Sci. 2022, 105, 2242–2255. [Google Scholar] [CrossRef] [PubMed]
- Jeyanathan, J.; Martin, C.; Morgavi, D.P. Screening of bacterial direct-fed microbials for their antimethanogenic potential in vitro and assessment of their effect on ruminal fermentation and microbial profiles in sheep. J. Anim. Sci. 2016, 94, 739–750. [Google Scholar] [CrossRef]
- Nie, L.; Zhang, A.; Jiang, N.; Yang, Z.N. The application of probiotics in the production of juvenile ruminants. Feed Expo. 2017, 9, 11–14. [Google Scholar]
- Pan, K.; He, M. Research and application of microbial additives in my country. Vet. Med. Feed Addit. 2002, 4, 35–37. [Google Scholar]
- Chen, G.; Pan, K.; Yuan, C.; Shen, J. Study on the effect of Bacillus PAS38 and β-mannan on 5-HT in rabbit small intestine. Chin. Anim. Husb. Newsl. 2008, 22, 11–13. [Google Scholar]
- Choi, P.; Rhayat, L.; Pinloche, E.; Devillard, E.; De Paepe, E.; Vanhaecke, L.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F.; Goossens, E. Bacillus Subtilis 29784 as a Feed Additive for Broilers Shifts the Intestinal Microbial Composition and Supports the Production of Hypoxanthine and Nicotinic Acid. Animals 2021, 11, 1335. [Google Scholar] [CrossRef]
- Chang, M.; Ma, F.; Wei, J.; Liu, J.; Nan, X.; Sun, P. Live Bacillus subtilis natto promotes rumen fermentation by modulating rumen microbiota in vitro. Animals 2021, 11, 1519. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, J.Q.; Deng, L.F. Effects of Bacillus subtilis natto on milk production, rumen fermentation and ruminal microbiome of dairy cows. Animal 2013, 7, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; He, Z.; Beauchemin, K.A.; Tang, S.; Zhou, C.; Han, X.; Wang, M.; Kang, J.; Odongo, N.E.; Tan, Z. Comparison of two live Bacillus species as feed additives for improving in vitro fermentation of cereal straws. Anim. Sci. J. 2016, 87, 27–36. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Bampidis, V.; Azimonti, G.; Bastos, M.L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; et al. Safety and efficacy of a feed additive consisting of Bacillus subtilis strains CNCM I-4606, CNCM I-5043 and CNCM I-4607 and Lactococcus lactis CNCM I-4609 for all animal species (Nolivade). EFSA J. 2021, 19, e06907. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Zheng, D.Z. Application of Calsporin in livestock feed. Anim. Husb. Mark. 2007, 10, 49–51. [Google Scholar]
- Geng, C. Comparison of Growth Performance, Carcass Index and Beef Quality of Fattening Cattle Fed with Active Yeast and Yeast Culture. Ph.D. Dissertation, China Agricultural University, Beijing, China, 2015. [Google Scholar]
- Feng, Z.; Gao, M. Improvement of the method for determination of ammonia nitrogen content in rumen liquid by colorimetry. Inn. Mong. Anim. Husb. Sci. 1993, 4, 40–41. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Stewart, C.S.; Duncan, S.H. The effect of avoparcin on cellulolytic bacteria of the ovine rumen. J. Gen. Microbiol. 1985, 131, 427–435. [Google Scholar] [CrossRef][Green Version]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from plant tissues. In Plant Molecular Biology Manual; Springer: Dordrecht, The Netherlands, 1989; pp. 73–83. [Google Scholar]
- Lee, P.Y.; Costumbrado, J.; Hsu, C.Y.; Kim, Y.H. Agarose gel electrophoresis for the separation of DNA fragments. J. Vis. Exp. 2012, 62, 3923. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wurilege. The Effect of Adding Bacillus subtilis to the Diet on the Growth Performance, Blood Indicators and Rumen Microbes of Fattening Sheep. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2019. [Google Scholar] [CrossRef]
- Xiros, C.; Shahab, R.L.; Studer, M.H. A cellulolytic fungal biofilm enhances the consolidated bioconversion of cellulose to short chain fatty acids by the rumen microbiome. Appl. Microbiol. Biotechnol. 2019, 103, 3355–3365. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, R.L., VI; Connor, E.E. Rumen Function and Development. Vet. Clin. N. Am. Food Anim. Pract. 2017, 33, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, D.; Angthong, W.; Keaokliang, O.; Ishida, T.; Takahashi, T.; Kawashima, T. Effect of feeding Bacillus subtilis on rumen fermentation, blood metabolites, nutrient digestibility, and energy and nitrogen balances in non-lactating crossbred cows. Anim. Sci. J. 2021, 92, e13531. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yu, Y.; Shen, Y.; Li, Q.; Lan, J.; Wu, Y.; Zhang, R.; Cao, G.; Yang, C. Effects of Bacillus subtilis and Bacillus licheniformis on growth performance, immunity, short chain fatty acid production, antioxidant capacity, and cecal microflora in broilers. Poult. Sci. 2021, 100, 101358. [Google Scholar] [CrossRef] [PubMed]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- Devyatkin, V.; Mishurov, A.; Kolodina, E. Probiotic effect of Bacillus subtilis B-2998D, B-3057D, and Bacillus licheniformis B-2999D complex on sheep and lambs. J. Adv. Vet. Anim. Res. 2021, 8, 146–157. [Google Scholar] [CrossRef]
- Konopka, A. What is microbial community ecology? ISME J. 2009, 3, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.K.; Zhang, X.X.; Li, F.D.; Li, C.; Li, G.Z.; Zhang, D.Y.; Song, Q.Z.; Li, X.L.; Zhao, Y.; Wang, W.M. Characterization of the rumen microbiota and its relationship with residual feed intake in sheep. Animal 2021, 15, 100161. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Sekita, A.; Chiji, H.; Kato, N. Consumption of lily bulb modulates fecal ratios of firmicutes and bacteroidetes phyla in rats fed a high-fat diet. Food Sci. Biotechnol. 2016, 25 (Suppl. S1), 153–156. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Ren, Z.; Fang, H.; Zhang, J.; Wang, R.; Xiao, W.; Zheng, K.; Yu, H.; Zhao, Y. Dietary Aronia melanocarpa Pomace Supplementation Enhances the Expression of ZO-1 and Occludin and Promotes Intestinal Development in Pigs. Front. Vet. Sci. 2022, 9, 904667. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.M.; Zhou, Y.L.; Almeida, A.; Finn, R.D.; Danchin, A.; He, L.S. Phylogenomics of expanding uncultured environmental Tenericutes provides insights into their pathogenicity and evolutionary relationship with Bacilli. BMC Genom. 2020, 21, 408. [Google Scholar] [CrossRef]
- Santarossa, S.; Sitarik, A.R.; Johnson, C.C.; Li, J.; Lynch, S.V.; Ownby, D.R.; Ramirez, A.; Yong, G.L.M.; Cassidy-Bushrow, A.E. Associations of physical activity with gut microbiota in pre-adolescent children. Phys. Act. Nutr. 2021, 25, 24–37. [Google Scholar] [CrossRef]
- Sun, D.; Bai, R.; Zhou, W.; Yao, Z.; Liu, Y.; Tang, S.; Ge, X.; Luo, L.; Luo, C.; Hu, G.F.; et al. Angiogenin maintains gut microbe homeostasis by balancing α-Proteobacteria and Lachnospiraceae. Gut 2021, 70, 666–676. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Li, M.; Campbell, T.B.; Flores, S.C.; Linderman, D.; Gebert, M.J.; Knight, R.; Fontenot, A.P.; Palmer, B.E. Alterations in the gut microbiota associated with HIV-1 infection. Cell Host Microbe 2013, 14, 329–339. [Google Scholar] [CrossRef]
- Xu, Y.; Ai, C.; Jiang, P.; Sun, X.; Liu, Y.; Jiang, G.; Song, S. Oligosaccharides from Gracilaria lemaneiformis better attenuated high fat diet-induced metabolic syndrome by promoting the Bacteroidales proliferation. Food Funct. 2020, 11, 1049–1062. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).