
Fructan Catabolism by Rumen Microbiota of Cattle and Sheep


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Media Composition
2.2. Animals and Rumen Fluid Collection
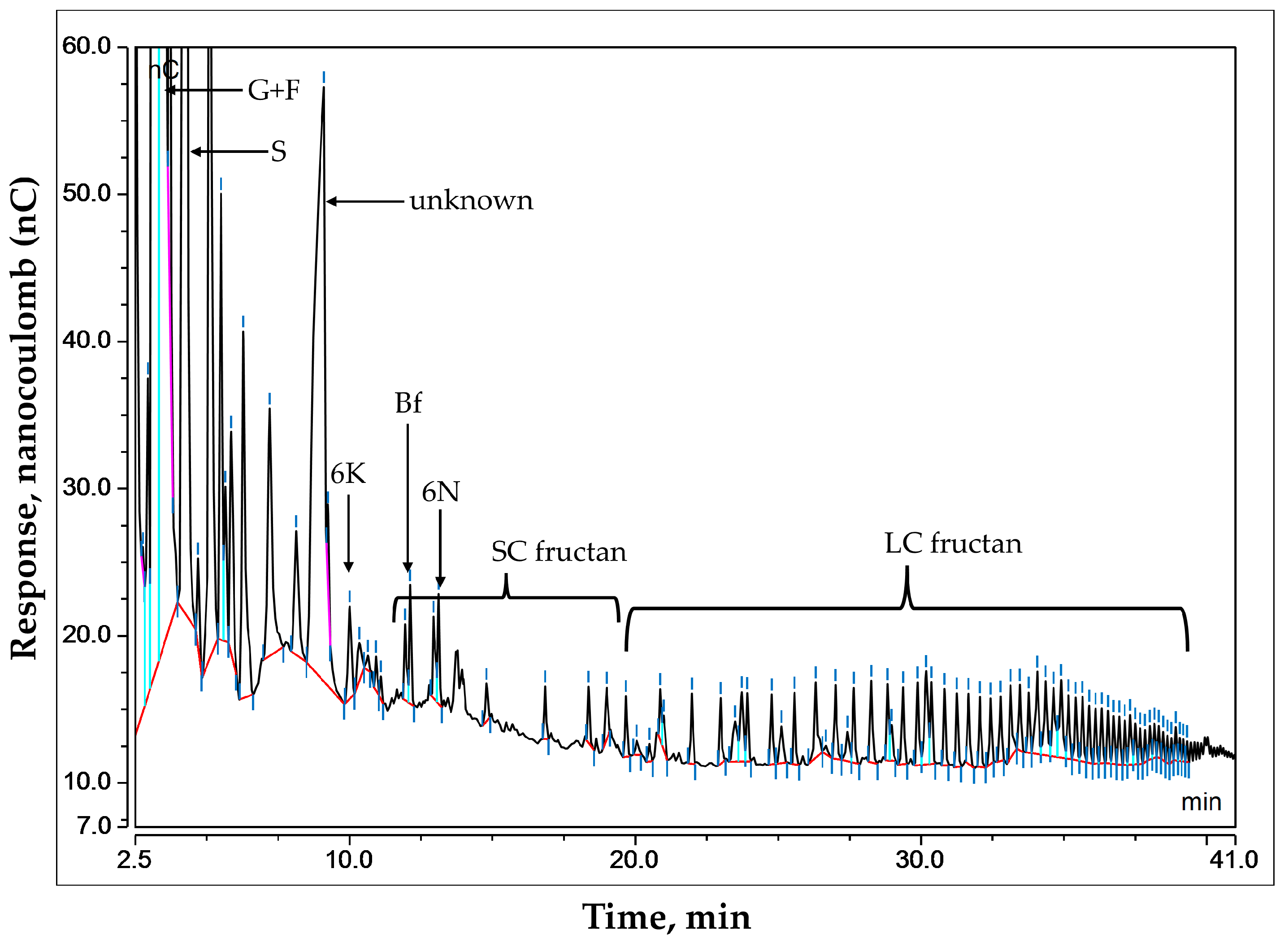
2.3. Analysis of Fructans and Mono/Disaccharides
2.4. Quantification of Fructans and Mono/Disaccharides
2.5. Fermentation End-Product Analyses
2.6. Chemical Composition of Orchardgrass Substrate
2.7. Statistical Analysis
3. Results
3.1. Orchardgrass Controls
3.2. Fructan Degradation
3.3. Fermentation pH
3.4. Fermentation End-Products
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hendry, G.A.F.; Wallace, R.K. The origin, distribution, and evolutionary significance of fructans. In Science and Technology of Fructans; Suzuki, M., Chatterton, N.J., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1993; pp. 119–139. [Google Scholar]
- Pollock, C.J. Tansley review no. 5 fructans and the metabolism of sucrose in vascular plants. New Phytol. 1986, 104, 1–24. [Google Scholar] [CrossRef]
- Livingston, D.P.; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef]
- Ernst, M.; Chatterton, N.J.; Harrison, P.A. Purification and characterization of a new fructan series from species of Asteraceae. New Phytol. 1996, 132, 63–66. [Google Scholar] [CrossRef]
- French, A.D.; Waterhouse, A.L. Chemical structure and characteristics. In Science and Technology of Fructans; Suzuki, M., Chatterton, N.J., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1993; pp. 119–139. [Google Scholar]
- Vijn, I.; Smeekens, S. Fructan: More than a reserve carbohydrate? Plant Physiol. 1999, 120, 351–359. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Chatterton, N.J. Glossary of fructan terms. In Science and Technology of Fructans; Suzuki, M., Chatterton, N.J., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1993; pp. 119–139. [Google Scholar]
- Benkeblia, N. Fructooligosaccharides and fructans analysis in plants and food crops. J. Chromatogr. A 2013, 1313, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Pollock, C.J.; Jones, T. Seasonal patterns of fructan metabolism in forage grasses. New Phytol. 1979, 83, 9–15. [Google Scholar] [CrossRef]
- Suzuki, M. Fructans in forage grasses with varying degrees of coldhardiness. J. Plant Physiol. 1989, 134, 224–231. [Google Scholar] [CrossRef]
- Pavis, N.; Boucaud, J.; Prud’Homme, M.P. Fructans and fructan-metabolizing enzymes in leaves of Lolium perenne. New Phytol. 2001, 150, 97–109. [Google Scholar] [CrossRef]
- Kagan, I.A.; Kirch, B.H.; Thatcher, C.D.; Strickland, J.R.; Elvinger, F.; Teutsch, C.D.; Pleasant, S.R. Seasonal and diurnal variation in simple sugar and fructan composition of orchardgrass pasture and hay in the Piedmont region of the United States. J. Equine Vet. Sci. 2011, 31, 488–497. [Google Scholar] [CrossRef]
- Waite, R.; Boyd, J. The water-soluble carbohydrates of grasses: Part I. Changes occurring during the normal life-cycle. J. Sci. Food Agric. 1953, 4, 197–204. [Google Scholar] [CrossRef]
- Volaire, F.; Lelièvre, F. Production, persistence, and water-soluble carbohydrate accumulation in 21 contrasting populations of Dactylis glomerata L. subjected to severe drought in the south of France. Aust. J. Agric. Res. 1997, 48, 933–944. [Google Scholar] [CrossRef]
- Chatterton, N.J.; Harrison, P.A.; Bennett, J.H.; Asay, K.H. Carbohydrate partitioning in 185 accessions of Gramineae grown under warm and cool temperatures. J. Plant Physiol. 1989, 134, 169–179. [Google Scholar] [CrossRef]
- Jensen, K.B.; Harrison, P.; Chatterton, N.J.; Bushman, B.S.; Creech, J.E. Seasonal trends in nonstructural carbohydrates in cool-and warm-season grasses. Crop. Sci. 2014, 54, 2328–2340. [Google Scholar] [CrossRef]
- Thomas, G.J. Metabolism of the soluble carbohydrates of grasses in the rumen of the sheep. J. Agric. Sci. 1960, 54, 360–372. [Google Scholar] [CrossRef]
- Hall, M.B.; Weimer, P.J. Divergent utilization patterns of grass fructan, inulin, and other nonfiber carbohydrates by ruminal microbes. J. Dairy Sci. 2016, 99, 245–257. [Google Scholar] [CrossRef]
- Perrin, S.; Fougnies, C.; Grill, J.P.; Jacobs, H.; Schneider, F. Fermentation of chicory fructo-oligosaccharides in mixtures of different degrees of polymerization by three strains of bifidobacteria. Can. J. Microbiol. 2002, 48, 759–763. [Google Scholar] [CrossRef]
- Ince, J.C.; Longland, A.C.; Moore-Colyer, M.J.S.; Harris, P.A. In vitro degradation of grass fructan by equid gastrointestinal digesta. Grass Forage Sci. 2013, 69, 514–523. [Google Scholar] [CrossRef]
- Harlow, B.E.; Kagan, I.A.; Lawrence, L.M.; Flythe, M.D. Effects of inulin chain length on fermentation by equine fecal bacteria and Streptococcus bovis. J. Equine Vet. Sci. 2017, 48, 113–120. [Google Scholar] [CrossRef]
- Kagan, I.A.; Harlow, B.E.; Flythe, M.D. A chromatographic method to monitor fructan catabolism in two cool-season grasses fermented by mixed bovine ruminal microbiota. JSFA Rep. 2022, 2, 264–271. [Google Scholar] [CrossRef]
- Ziołecki, A.; Guczynska, W.; Wojciechowicz, M. Some rumen bacteria degrading fructan. Lett. Appl. Microbiol. 1992, 15, 244–247. [Google Scholar] [CrossRef]
- Weinert-Nelson, J.R.; Ely, D.G.; Flythe, M.D.; Hamilton, T.A.; Ferrell, J.L.; Davis, B.E. Ex vivo fermentation of hay and corn by rumen bacteria from cattle and sheep. Fermentation 2023, 101, 66–67. [Google Scholar]
- Biggs, D.R.; Hancock, K.R. In vitro digestion of bacterial and plant fructans and effects on ammonia accumulation in cow and sheep rumen fluids. J. Gen. Appl. Microbiol. 1998, 44, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Stack, R.J.; Hungate, R.E.; Opsahl, W.P. Phenylacetic acid stimulation of cellulose digestion by Ruminococcus albus 8. Appl. Environ. Microbiol. 1983, 46, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Harlow, B.E.; Flythe, M.D.; Aiken, G.E. Effect of biochanin A on corn grain (Zea mays) fermentation by bovine rumen amylolytic bacteria. J. Appl. Microbiol. 2017, 122, 870–880. [Google Scholar] [CrossRef]
- FASS. Guide for the Care and Use of Agricultural Animals in Research and Teaching; FASS Inc.: Champaign, IL, USA, 2020. [Google Scholar]
- Chatterton, N.J.; Harrison, P.A. Fructan oligomers in Poa ampla. New Phytol. 1997, 136, 3–10. [Google Scholar] [CrossRef]
- Rocklin, R.D.; Pohl, C.A. Determination of carbohydrates by anion exchange chromatography with pulsed amperometric detection. J. Liq. Chromatogr. 1983, 6, 1577–1590. [Google Scholar] [CrossRef]
- Rocklin, R.D.; Clarke, A.P.; Weitzhandler, M. Improved long-term reproducibility for pulsed amperometric detection of carbohydrates via a new quadruple-potential waveform. Anal. Chem. 1998, 70, 1496–1501. [Google Scholar] [CrossRef]
- Pöhnl, T.; Böttcher, C.; Schulz, H.; Stürtz, M.; Widder, S.; Carle, R.; Schweiggert, R.M. Comparison of high performance anion exchange chromatography with pulsed amperometric detection (HPAEC-PAD) and ultra-high performance liquid chromatography with evaporative light scattering (UHPLC-ELSD) for the analyses of fructooligosaccharides in onion (Allium cepa L.). J. Food Comp. Anal. 2017, 63, 48–156. [Google Scholar] [CrossRef]
- ANKOM Technology. Method 6: Acid Detergent Fiber in Feeds—Filter Bag Technique (for A200 and A2001); ANKOM Technology: Macedon, NY, USA, 2005; Available online: https://www.ankom.com/analytical-methods-support/fiber-analyzer-a200?f%5B0%5D=field_faq_group%3A89#:~:text=Acid-,Detergent,-Fiber%20Method%20(A200) (accessed on 26 July 2022).
- ANKOM Technology. Method 6: Neutral Detergent Fiber in Feeds—Filter Bag Technique (for A200 and A2001); ANKOM Technology: Macedon, NY, USA, 2006; Available online: https://www.ankom.com/analytical-methods-support/fiber-analyzer-a200?f%5B0%5D=field_faq_group%3A89#:~:text=View%20PDF-,Neutral,-Detergent%20Fiber%20Method (accessed on 26 July 2022).
- Kagan, I.A.; Lawrence, L.M.; Seman, D.H.; Prince, K.J.; Fowler, A.L.; Smith, S.R. Effects of sampling time, cultivar, and methodology on water- and ethanol-soluble carbohydrate profiles of three cool-season grasses. J. Equine Vet. Sci. 2018, 61, 99–107. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hall, M.B.; Hoover, W.H.; Jennings, J.P.; Webster, T.K.M. A method for partitioning neutral detergent-soluble carbohydrates. J. Sci. Food Agric. 1999, 79, 2079–2086. [Google Scholar] [CrossRef]
- Li, J.; Yan, H.; Chen, J.; Duan, C.; Guo, Y.; Liu, Y.; Zhang, Y.; Ji, S. Correlation of ruminal fermentation parameters and rumen bacterial community by comparing those of the goat, sheep, and cow in vitro. Fermentation 2022, 8, 427. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed]
- Chatterton, N.J.; Harrison, P.A.; Thornley, W.R.; Bennett, J.H. Structures of fructan oligomers in orchardgrass (Dactylis glomerata L.). J. Plant Physiol. 1993, 142, 552–556. [Google Scholar] [CrossRef]
- Cairns, A.J.; Nash, R.; Machado de Carvalho, M.-A.; Sims, I.M. Characterization of the enzymatic polymerization of 2,6-linked fructan by leaf extracts from timothy grass (Phleum pratense). New Phytol. 1999, 142, 79–91. [Google Scholar] [CrossRef]
- Kagan, I.A.; Kirch, B.H.; Strickland, J.R. A chromatographic survey of methods for extracting long-chain grass fructans. Grass Forage Sci. 2011, 66, 434–448. [Google Scholar] [CrossRef]
- Chatterton, N.J.; Harrison, P.A.; Thornley, W.R.; Draper, E.A. Oligosaccharides in foliage of Agropyron, Bromus, Dactylis, Festuca, Lolium and Phleum. New Phytol. 1990, 114, 67–171. [Google Scholar] [CrossRef]
- Joyce, G.E.; Kagan, I.A.; Flythe, M.D.; Davis, B.E.; Schendel, R.R. Profiling of cool-season forage arabinoxylans via a validated HPAEC-PAD method. Front. Plant Sci. 2023, 14, 1116995. [Google Scholar] [CrossRef]
- Müller, M.; Seyfarth, W. Purification and substrate specificity of an extracellular fructanhydrolase from Lactobacillus paracasei ssp. paracasei P 4134. New Phytol. 1997, 136, 89–96. [Google Scholar] [CrossRef]
- Kasperowicz, A.; Michalowski, T. Assessment of the fructanolytic activities in the rumen bacterium Treponema saccharophilum strain S. J. Appl. Microbiol. 2002, 92, 140–146. [Google Scholar] [CrossRef]
- Kasperowicz, A.; Pristaš, P.; Piknová, M.; Javorský, P.; Guczyńska, W.; Michałovski, T.; Kwiatkowska, E. Fructanolytic and saccharolytic enzymes of Treponema zioleckii strain kT. Anaerobe 2010, 16, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Owens, F.; Secrist, D.; Hill, W.; Gill, D. Acidosis in cattle: A review. J. Anim. Sci. 1998, 76, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Tech. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Nagaraja, T.; Titgemeyer, E. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook 1, 2. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef]
- Russell, J.B.; Dombrowski, D. Effect of pH on the efficiency of growth by pure cultures of rumen bacteria in continuous culture. Appl. Environ. Microbiol. 1980, 39, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Weimer, P. Response surface analysis of the effects of pH and dilution rate on Ruminococcus flavefaciens FD-1 in cellulose-fed continuous culture. Appl. Environ. Microbiol. 1992, 58, 2583–2591. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J. Effects of dilution rate and pH on the ruminal cellulolytic bacterium Fibrobacter succinogenes S85 in cellulose-fed continuous culture. Arch. Microbiol. 1993, 160, 288. [Google Scholar] [CrossRef]
- Kasperowicz, A.; Stan-Głasek, K.; Kowalik, B.; Vandzurova, A.; Pristas, P.; Pająk, J.; Kwiatkowska, E.; Michałowski, T. Effect of dietary fructose polymers or sucrose on microbial fermentation, enzyme activity, ciliate concentration and diversity of bacterial flora in the rumen of rams. Anim. Feed Sci. Technol. 2014, 195, 38–46. [Google Scholar] [CrossRef]
- Fan, Y.; Xia, G.; Jin, Y.; Wang, H. Ambient pH regulates lactate catabolism pathway of the ruminal Megasphaera elsdenni BE2-2083 and Selenomonas ruminantium HD4. J. Appl. Microbiol. 2022, 132, 2661–2672. [Google Scholar] [CrossRef]
- Cuchillo-Hilario, M.; Wrage-Mönnig, N.; Isselstein, J. Forage selectivity by cattle and sheep co-grazing swards differing in plant species diversity. Grass Forag Sci. 2018, 73, 320–329. [Google Scholar] [CrossRef]
- Grant, S.A.; Suckling, D.E.; Smith, H.K.; Torvell, L.; Forbes, T.D.A.; Hodgson, J. Comparative studies of diet selection by sheep and cattle: The hill grasslands. J. Ecol. 1985, 73, 987–1004. [Google Scholar] [CrossRef]
- Rutter, S.M. Grazing preferences in sheep and cattle: Implications for production, the environment and animal welfare. Can. J. Anim. Sci. 2010, 90, 285–293. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Concentration, g kg DM−1 |
|---|---|
| Crude Protein | 143.5 |
| Acid Detergent Fiber | 353.9 |
| Neutral Detergent Fiber | 578.4 |
| Water-Soluble Carbohydrate | 91.5 |
| Ethanol-Soluble Carbohydrate | 74.1 |
| Fructan/Sugar | Concentration, μg mL−1 |
|---|---|
| Long-chain † | 444.1 |
| Short-chain † | 30.0 |
| Bifurcose | 11.3 |
| 1-Nystose | 0.0 |
| 6-Nystose | 7.6 |
| 1-Kestose | 0.0 |
| 6-Kestose | 3.4 |
| Neokestose | 0.0 |
| Raffinose | 7.5 |
| Free Sugars ‡ | 1,064.8 |
| Time, h | p-Values | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fructan/Sugar, μg mL−1 | 0 | 2 | 4 | 8 | SE | Species | Time | Species × Time | ||||
| Bovine | Ovine | Bovine | Ovine | Bovine | Ovine | Bovine | Ovine | |||||
| Long-chain † | 408.2 α | 421.7 α | 383.1 x,α | 349.0 y,β | 0.0 b,β | 50.2 a,γ | 0.0 β | 0.0 δ | 16.8 | 0.44 | <0.01 | 0.04 |
| Short-chain † | 53.7 β | 85.8 β | 185.4 b,α | 255.9 a,α | 19.0 b,β | 249.4 a,α | 4.7 β | 11.2 γ | 21.5 | <0.01 | <0.01 | <0.01 |
| Bifurcose | 12.2 β | 15.4 γ | 40.1 α | 48.9 β | 2.1 β | 85.8 α | 0.0 β | 4.8 γ | 9.6 | <0.01 | <0.01 | <0.01 |
| 1-Nystose | 1.7 β | 3.4 γ | 14.1 α | 17.9 β | 0.7 a,β | 30.3 b,α | 0.0 β | 0.0 γ | 3.4 | <0.01 | <0.01 | <0.01 |
| 6-Nystose | 7.9 | 8.1 | 11.3 | 10.8 | 8.5 | 8.4 | 0.7 | 1.5 | 1.4 | 0.91 | <0.01 | 0.96 |
| 1-Kestose | 0.3 b | 8.9 a | 4.1 | 0.4 | 0.0 | 2.2 | 0.0 | 0.0 | 2.2 | 0.40 | 0.15 | 0.06 |
| 6-Kestose | 6.0 | 7.0 | 9.7 | 8.6 | 7.9 | 10.8 | 0.0 | 0.7 | 1.8 | 0.51 | <0.01 | 0.74 |
| Neokestose | 0.00 | 0.0 | 0.7 | 0.3 | 0.0 | 0.5 | 0.0 | 0.0 | 0.2 | 0.78 | 0.05 | 0.20 |
| Raffinose | 5.5 | 4.4 | 5.8 | 5.2 | 0.0 | 0.9 | 0.0 | 0.0 | 1.0 | 0.77 | <0.01 | 0.70 |
| Free Sugars ‡ | 1104.9 | 1084.0 | 908.6 | 750.2 | 83.4 | 230.6 | 43.1 | 49.6 | 65.0 | 0.85 | <0.01 | 0.11 |
| SCFA, mmol L−1 | Species | Statistics | ||
|---|---|---|---|---|
| Bovine | Ovine | SE | p-Value | |
| Lactate | 21.03 | 17.23 | 1.46 | 0.12 |
| Succinate | 1.82 | 1.19 | 0.75 | 0.58 |
| Acetate | 17.70 a | 14.42 b | 0.59 | <0.01 |
| Propionate | 8.25 | 8.00 | 0.73 | 0.82 |
| Butyrate | 1.91 | 1.78 | 0.11 | 0.44 |
| Total SCFA | 50.70 a | 42.62 b | 2.05 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinert-Nelson, J.R.; Kagan, I.A.; Ely, D.G.; Flythe, M.D.; Davis, B.E. Fructan Catabolism by Rumen Microbiota of Cattle and Sheep. Fermentation 2023, 9, 925. https://doi.org/10.3390/fermentation9110925
Weinert-Nelson JR, Kagan IA, Ely DG, Flythe MD, Davis BE. Fructan Catabolism by Rumen Microbiota of Cattle and Sheep. Fermentation. 2023; 9(11):925. https://doi.org/10.3390/fermentation9110925
Chicago/Turabian StyleWeinert-Nelson, Jennifer R., Isabelle A. Kagan, Donald G. Ely, Michael D. Flythe, and Brittany E. Davis. 2023. "Fructan Catabolism by Rumen Microbiota of Cattle and Sheep" Fermentation 9, no. 11: 925. https://doi.org/10.3390/fermentation9110925
APA StyleWeinert-Nelson, J. R., Kagan, I. A., Ely, D. G., Flythe, M. D., & Davis, B. E. (2023). Fructan Catabolism by Rumen Microbiota of Cattle and Sheep. Fermentation, 9(11), 925. https://doi.org/10.3390/fermentation9110925