
Fermentation Dynamics of Naturally Fermented Palm Beverages of West Bengal and Jharkhand in India



Abstract
1. Introduction
2. Materials and Methods
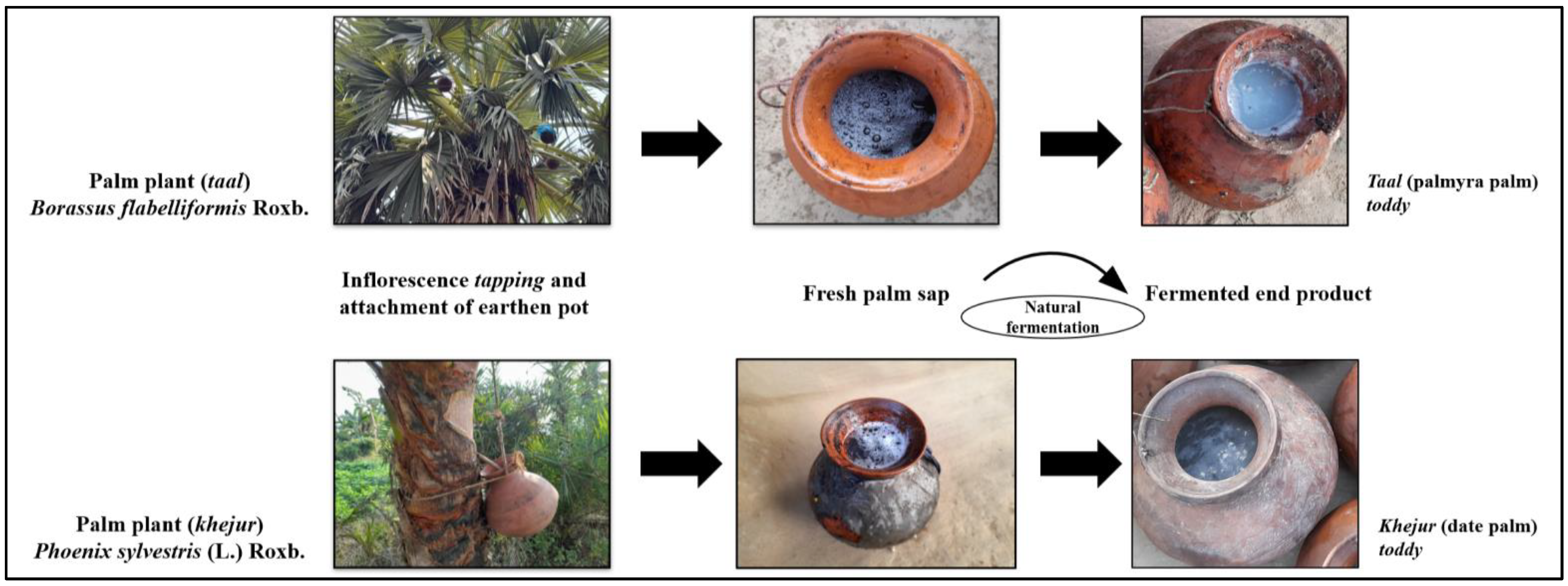
2.1. Collection of Samples
2.2. pH, Total Acidity and Total Alcohol Content
2.3. Proportion Distribution between Ethanol and Methanol
2.4. °BRIX (Degrees BRIX) Profile and Total Sugar Content
2.5. Ester Content
2.6. Total Crude Protein Content
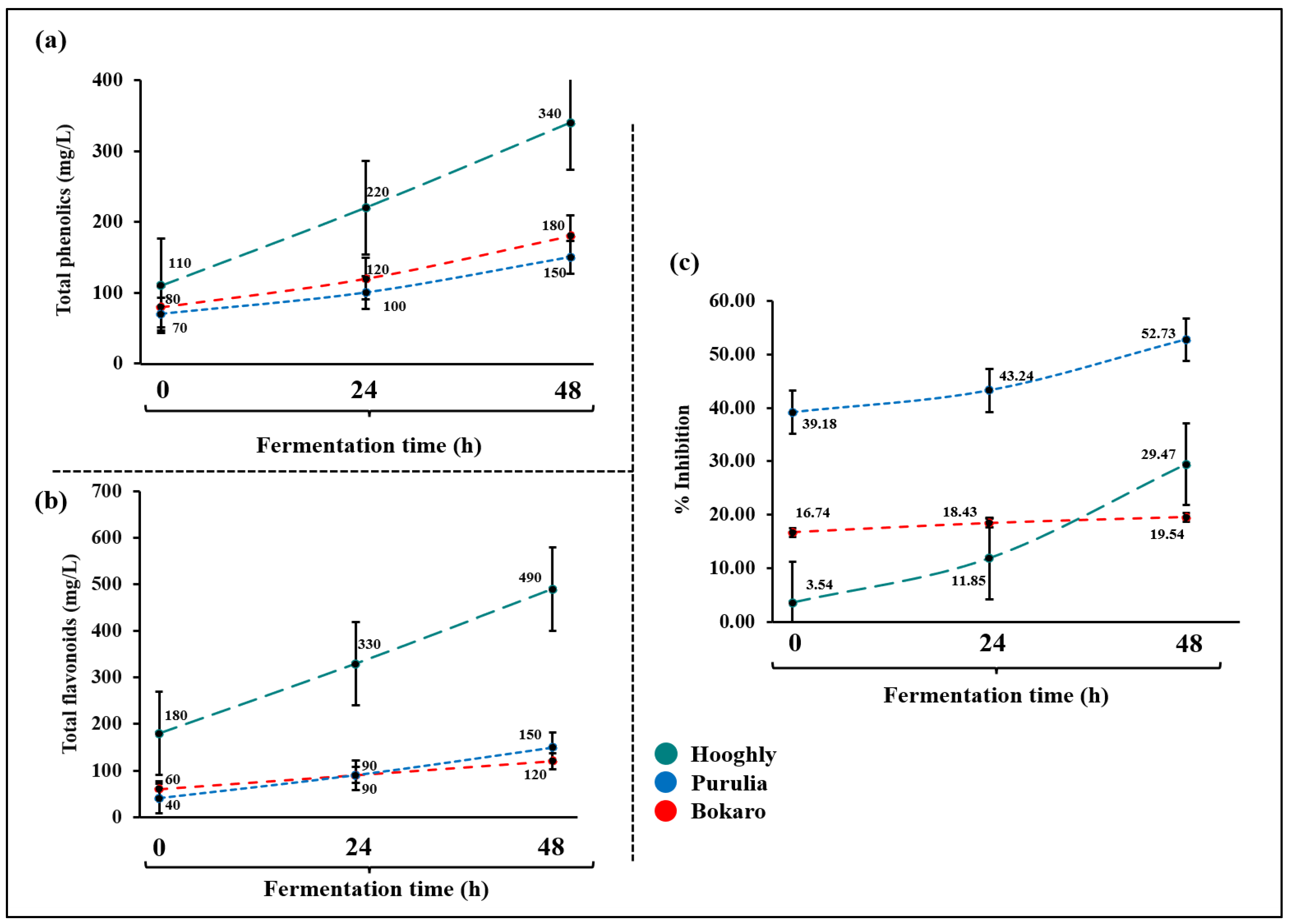
2.7. Total Phenolics and Flavonoids Contents
2.8. Anti-Oxidant Activity
2.9. Microbiological Analysis
2.10. Genomic DNA Isolation
2.11. Repetitive Sequence-Based PCR
2.12. Amplification and DNA Sequencing
2.13. Bioinformatics Analysis
2.14. Statistics
3. Result
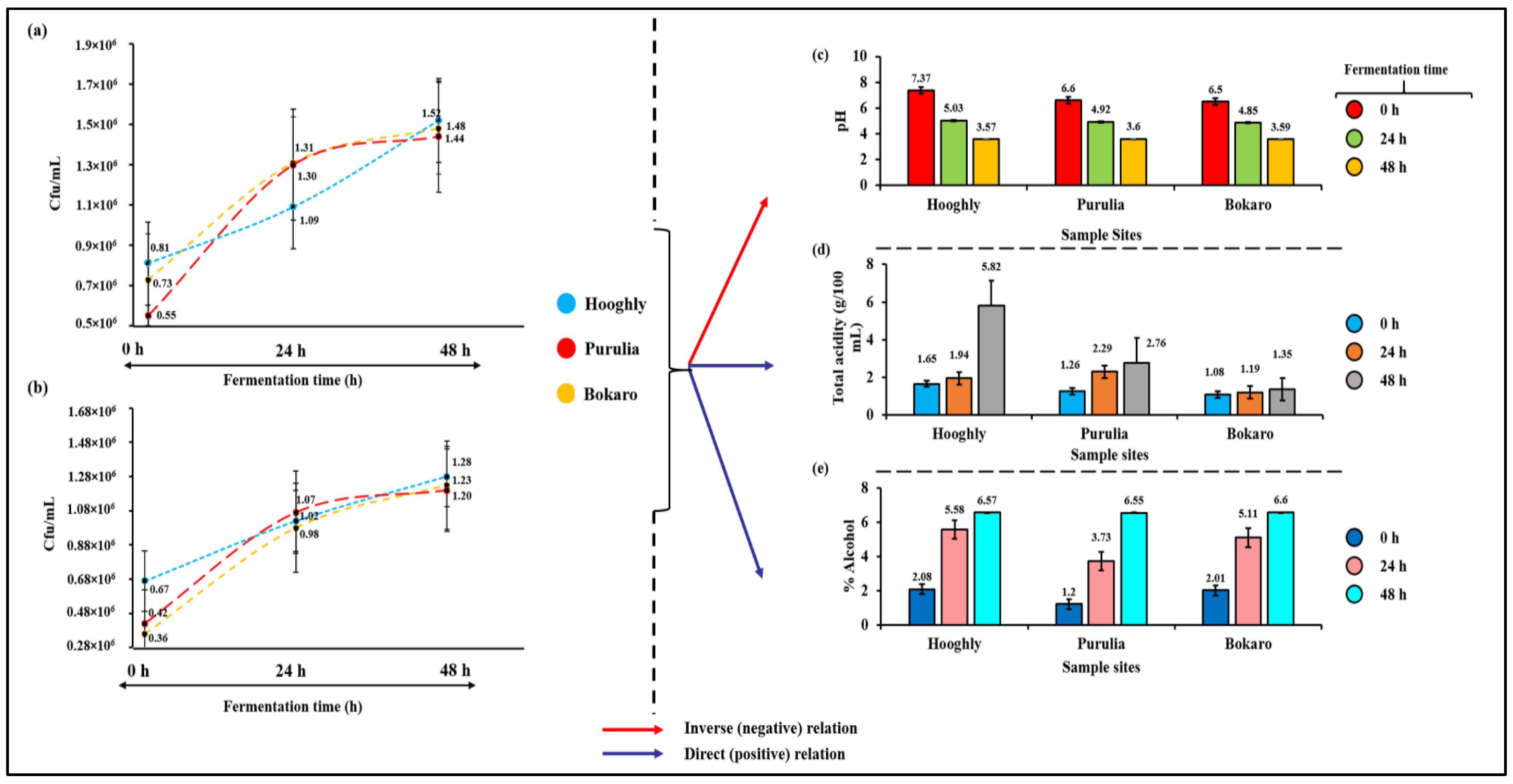
3.1. Microbial Changes
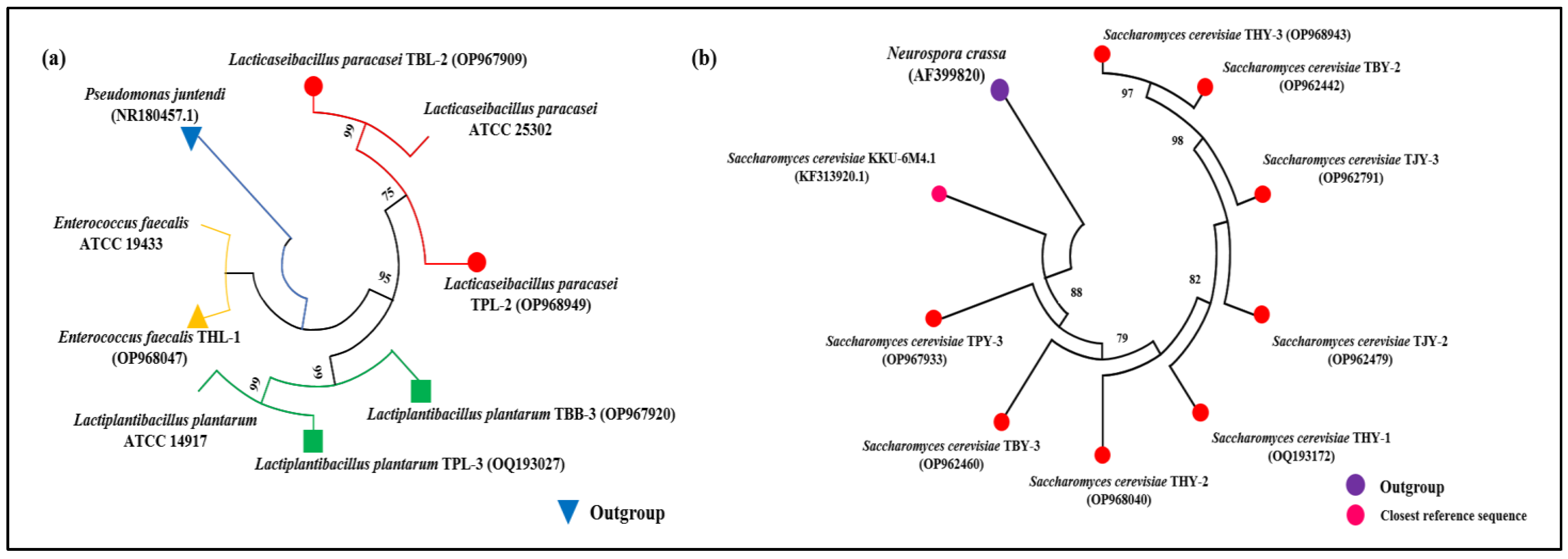
3.2. Microbial Identification
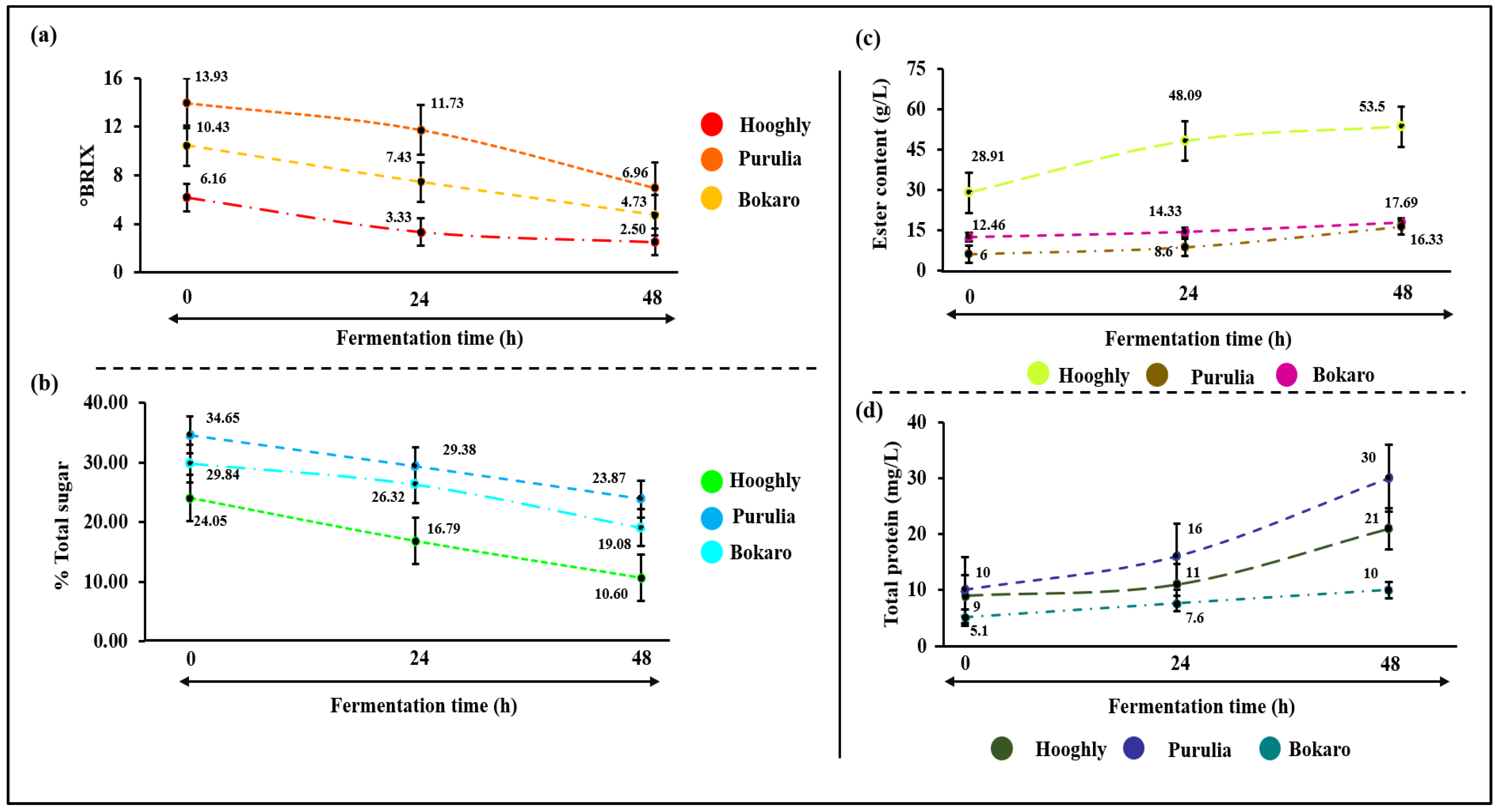
3.3. Physico-Chemical Changes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tamang, J.P. “Ethno-microbiology” of ethnic Indian fermented foods and alcoholic beverages. J. Appl. Microbiol. 2022, 133, 145–161. [Google Scholar] [CrossRef]
- Tamang, J.P. Dietary culture and antiquity of the Himalayan fermented foods and alcoholic fermented beverages. J. Ethnic Foods 2022, 9, 30. [Google Scholar] [CrossRef]
- Tamang, J.P.; Cotter, P.D.; Endo, A.; Han, N.S.; Kort, R.; Liu, S.Q.; Mayo, B.; Westerik, N.; Hutkins, R. Fermented foods in a global age: East meets West. Comp. Rev. Food Sci. Food Saf. 2020, 19, 184–217. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Tamang, J.P. Changes in microbial communities and their predictive functionalities during fermentation of toddy, an alcoholic beverage of India. Microbiol. Res. 2021, 248, 126769. [Google Scholar] [CrossRef]
- Amoikon, T.L.S.; Marcotte, S.; Djeni, T.N.; N’sa, K.M.C.; Grondin, C.; Tinsley, C.; Casaregola, S.; Djè, M.K. A study on the potential of yeasts isolated from palm wines to produce flavouring compounds. LWT 2020, 128, 109506. [Google Scholar] [CrossRef]
- Prathiviraj, R.; Rajeev, R.; Jose, C.M.; Begum, A.; Selvin, J.; Kiran, G.S. Fermentation microbiome and metabolic profiles of Indian palm wine. Gene Rep. 2022, 27, 101543. [Google Scholar] [CrossRef]
- Bandyopadhyay, M.; Chakraborti, K.; Samanta, A.; Bandyopadhyay, A. Date-sugar-palm based folklore of Bengal. Ethnobot. Med. Plants 2012, 2, 689–695. [Google Scholar]
- Djeni, T.N.; Keisam, S.; Kouame, K.H.; Assohoun-Djeni, C.N.M.; Aké, M.D.F.; Amoikon, T.L.S.; Tuikhar, N.; Labala, R.K.; Djè, M.K.; Jeyaram, K. Dynamics of microbial populations and metabolites of fermenting saps throughout tapping process of ron and oil palm trees in Côte d’Ivoire. Front. Microbiol. 2022, 13, 4372. [Google Scholar] [CrossRef] [PubMed]
- Djeni, T.N.; Kouame, K.H.; Ake, M.D.F.; Amoikon, T.L.S.; Djè, M.K.; Jeyaram, K. Microbial diversity and metabolite profiles of palm wine produced from three different palm tree species in Côte d’Ivoire. Sci. Rep. 2020, 10, 1715. [Google Scholar] [CrossRef]
- Tra Bi, C.Y.; Amoikon, T.L.S.; Kouakou, C.A.; Noemie, J.; Lucas, M.; Grondin, C.; Legras, J.; N’guessan, F.K.; Djeni, T.N.; Djè, M.K.; et al. Genetic diversity and population structure of Saccharomyces cerevisiae strains isolated from traditional alcoholic beverages of Côte d’Ivoire. Int. J. Food Microbiol. 2019, 297, 1–10. [Google Scholar] [CrossRef]
- Karamoko, D.; Bouatenin, J.P.K.M.; Yoro, D.T.; Djeni, N.T.; Djè, M.K. Dynamics of lactic acid bacteria isolated from palm wine during the exploitation of two varieties of oil palm for the production of new beverages. J. Glob. Biosci. 2019, 8, 5990–6002. [Google Scholar]
- Mahulette, F.; Astuti, D.I. Microbial succession and chemical characteristics in fermentation of Ambonese arrack (sopi), traditional beverage from Maluku. Biosaintifika J. Biol. Biol. Edu. 2020, 12, 147–154. [Google Scholar] [CrossRef]
- Shamala, T.R.; Sreekantiah, K.R. Microbiological and biochemical studies on traditional Indian palm wine fermentation. Food Microbiol. 1988, 5, 157–162. [Google Scholar] [CrossRef]
- Ouoba, L.I.I.; Kando, C.; Parkouda, C.; Sawadogo-Lingani, H.; Diawara, B.; Sutherland, J.P. The microbiology of Bandji, palm wine of Borassus akeassii from Burkina Faso: Identification and genotypic diversity of yeasts, lactic acid and acetic acid bacteria. J. Appl. Microbiol. 2012, 113, 1428–1441. [Google Scholar] [CrossRef] [PubMed]
- Astudillo-Melgar, F.; Ochoa-Leyva, A.; Utrilla, J.; Huerta-Beristain, G. Bacterial diversity and population dynamics during the fermentation of palm wine from Guerrero Mexico. Front. Microbiol. 2019, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health and Family Welfare, Government of India. FSSAI Manual of Methods of Analysis of Foods—Alcoholic Beverages. 2021. Available online: https://fssai.gov.in/upload/uploadfiles/files/Order_Manual_Analysis_Alcoholic_Beverages_20_07_2021.pdf (accessed on 1 February 2022).
- Latimer, G.W. Association of Official Analytical Chemists International, Official Methods of Analysis of AOAC International, 20th ed.; AOAC International: Rockville, MA, USA, 2016; p. 759. [Google Scholar]
- Coldea, T.E.; Socaciu, C.; Fetea, F.; Ranga, F.; Pop, R.M.; Florea, M. Rapid quantitative analysis of ethanol and prediction of methanol content in traditional fruit brandies from Romania, using FTIR spectroscopy and chemometrics. Not. Bot. Horti Agrobot. Cluj Napoca 2013, 41, 143–149. [Google Scholar] [CrossRef]
- Ofoedu, C.E.; Ofoedu, E.O.; Chacha, J.S.; Owuamanam, C.I.; Efekalam, I.S.; Awuchi, C.G. Comparative evaluation of physicochemical, antioxidant, and sensory properties of red wine as markers of its quality and authenticity. Int. J. Food Sci. 2022, 2022, 8368992. [Google Scholar] [CrossRef]
- Pandeya, A.; Rayamajhi, S.; Pokhrel, P.; Giri, B. Evaluation of secondary metabolites, antioxidant activity, and color parameters of Nepali wines. Food Sci. Nutri. 2018, 6, 2252–2263. [Google Scholar] [CrossRef]
- PotdarVrushali, B.; Gauri, R.D.; Anjali, J.M. Optimum parameters for wine production from fig fruit (Ficus Carica) juice. IOSR J. Environ. Sci. Toxicol. Food Technol. 2020, 14, 28–34. [Google Scholar]
- Milinčić, D.D.; Stanisavljević, N.S.; Kostić, A.Ž.; Bajić, S.S.; Kojić, M.O.; Gašić, U.M.; Barać, M.B.; Stanojević, S.P.; Tešić, Ž.L.; Pešić, M.B. Phenolic compounds and biopotential of grape pomace extracts from Prokupac red grape variety. LWT 2021, 138, 110739. [Google Scholar] [CrossRef]
- El Euch, S.K.; Bouajila, J.; Bouzouita, N. Chemical composition, biological and cytotoxic activities of Cistus salviifolius flower buds and leaves extracts. Ind. Crop. Prod. 2015, 76, 1100–1105. [Google Scholar] [CrossRef]
- Shangpliang, H.N.J.; Tamang, J.P. Phenotypic and genotypic characterisation of lactic acid bacteria isolated from exotic naturally fermented milk (cow and yak) products of Arunachal Pradesh, India. Int. Dairy J. 2021, 118, 105038. [Google Scholar] [CrossRef]
- Lama, S.; Tamang, J.P. Isolation of yeasts from some homemade fermented cow-milk products of Sikkim and their probiotic characteristics. Fermentation 2022, 8, 664. [Google Scholar] [CrossRef]
- Guimarães, T.M.; Moriel, D.G.; Machado, I.P.; Picheth, C.M.T.F.; Bonfim, T.M.B. Isolation and characterization of Saccharomyces cerevisiae strains of winery interest. Rev. Bras. Cienc. Farm. 2006, 42, 119–126. [Google Scholar] [CrossRef]
- Cissé, H.; Kagambèga, B.; Sawadogo, A.; Tankoano, A.; Sangaré, G.; Traoré, Y.; Ouoba, I.I.L.; Savadogo, A. Molecular characterization of Bacillus, lactic acid bacteria and yeast as potential probiotic isolated from fermented food. Sci. Afr. 2019, 6, e00175. [Google Scholar] [CrossRef]
- Palla, M.; Blandino, M.; Grassi, A.; Giordano, D.; Sgherri, C.; Quartacci, M.F.; Reyneri, A.; Agnolucci, M.; Giovannetti, M. Characterization and selection of functional yeast strains during sourdough fermentation of different cereal wholegrain flours. Sci. Rep. 2020, 10, 12856. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 1, 2–3. [Google Scholar] [CrossRef]
- Martino, C.; Morton, J.T.; Marotz, C.A.; Thompson, L.R.; Tripathi, A.; Knight, R.; Zengler, K. A novel sparse compositional technique reveals microbial perturbations. MSystems 2019, 4, e00016-19. [Google Scholar] [CrossRef]
- Kaur, J.; Sharma, A.; Lee, S.; Park, Y.S. DNA Profiling of Leuconostoc citreum strains in fermented foods by repetitive element polymerase chain reaction. J. Microbiol. Biotechnol. 2017, 27, 1778–1782. [Google Scholar] [CrossRef]
- Zongo, O.; Tapsoba, F.; Leray, F.; Bideaux, C.; Guillouet, S.; Traore, Y.; Savadogo, A. Nutritional, biochemical and microbiological composition of Borassus aethiopum Mart. sap in Burkina Faso. J. Food Sci. Technol. 2020, 57, 495–504. [Google Scholar] [CrossRef]
- Zongo, O.; Tapsoba, F.; Guira, F.; Zongo, C.; Traore, Y.; Savadogo, A. Evaluation of microbiological and physicochemical quality of Borassus akeassii fresh sap and fermented sap (bandji) produced at Burkina Faso. Int. J. Adv. Res. 2017, 5, 1489–1498. [Google Scholar]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic potential and safety evaluation of enterococcus faecalis ob14 and ob15, isolated from traditional Tunisian testouri cheese and rigouta, using physiological and genomic analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Watcharapong, M.; Phoomjai, S.; Chonticha, R. Lacticaseibacillus spp.; probiotic candidates from Palmyra palm sugar, possess antimicrobial, and anti-biofilm activities against methicillin-resistant Staphylococcus aureus. Vet. World 2022, 15, 299–308. [Google Scholar]
- Zhao, Q.; Tang, S.; Fang, X.; Wang, Z.; Jiang, Y.; Guo, X.; Zhu, J.; Zhang, Y. The effect of Lactiplantibacillus plantarum BX62 alone or in combination with chitosan on the qualitative characteristics of fresh-cut apples during cold storage. Microorganisms 2021, 9, 2404. [Google Scholar] [CrossRef]
- Chang, H.M.; Foo, H.L.; Loh, T.C.; Lim, E.T.C.; Mutalib, N.E.A. Comparative studies of inhibitory and antioxidant activities, and organic acids compositions of postbiotics produced by probiotic Lactiplantibacillus plantarum strains isolated from Malaysian foods. Front. Vet. Sci. 2021, 7, 602280. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Halami, P.M.; Tamang, J.P. Genome analysis of Lactobacillus plantarum isolated from some Indian fermented foods for bacteriocin production and probiotic marker genes. Front. Microbiol. 2020, 11, 40. [Google Scholar] [CrossRef]
- Yilmaz, B.; Bangar, S.P.; Echegaray, N.; Suri, S.; Tomasevic, I.; Lorenzo, J.M.; Melekoglu, E.; Rocha, J.M.; Ozogul, F. The impacts of Lactiplantibacillus plantarum on the functional properties of fermented foods: A review of current knowledge. Microorganisms 2022, 10, 826. [Google Scholar] [CrossRef]
- Gaur, G.; Gänzle, M.G. Conversion of (poly)phenolic compounds in food fermentations by lactic acid bacteria: Novel insights into metabolic pathways and functional metabolites. Cur. Res. Food Sci. 2023, 6, 100448. [Google Scholar] [CrossRef]
- Cázares-Vásquez, M.L.; Rodríguez-Herrera, R.; Aguilar-González, C.N.; Sáenz-Galindo, A.; Solanilla-Duque, J.F.; Contreras-Esquivel, J.C.; Flores-Gallegos, A.C. Microbial exopolysaccharides in traditional Mexican fermented beverages. Fermentation 2021, 7, 249. [Google Scholar] [CrossRef]
- Hu, J.; Vinothkanna, A.; Wu, M.; Ekumah, J.N.; Akpabli-Tsigbe, N.D.K.; Ma, Y. Tracking the dynamic changes of a flavor, phenolic profile, and antioxidant properties of Lactiplantibacillus plantarum- and Saccharomyces cerevisiae-fermented mulberry wine. Food Sci. Nutri. 2021, 9, 6294–6306. [Google Scholar] [CrossRef]
- Shetty, P.; D’Souza, A.; Poojari, S.; Narayana, J.; Rajeeva, P. Study of fermentation kinetics of palm sap from Cocos nucifera. Int. J. Appl. Sci. Biotechnol. 2017, 5, 375–381. [Google Scholar] [CrossRef]
- Paine, A.; Davan, A.D. Defining a tolerable concentration of methanol in alcoholic drinks. Hum. Exp. Toxicol. 2001, 20, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Sirhan, A.Y.; Wong, R.; Abdulra’uf, L.B.; Aljabar, J.A.; Mostafa, A.; Talhouni, A. Simultaneous determination of ethanol and methanol in alcohol free malt beverages, energy drinks and fruit juices by gas chromatography. Asian J. Agric. Biol. 2019, 7, 183–189. [Google Scholar]
- Bucher, T.; Frey, E.; Wilczynska, M.; Deroover, K.; Dohme, S. Consumer perception and behaviour related to low-alcohol wine: Do people overcompensate? Public Health Nutr. 2020, 23, 1939–1947. [Google Scholar] [CrossRef]
- Eiserhardt, W.L.E.; Svenning, J.C.; Kissling, W.D.; Balslev, H. Geographical ecology of the palms (Arecaceae): Determinants of diversity and distributions across spatial scales. Ann. Botany 2011, 108, 1391–1416. [Google Scholar] [CrossRef] [PubMed]
- Alshammari, N.; Alazmi, M.; Alanazi, N.A.; Sulieman, A.M.E.; Veettil, V.N.; Ponce-Alonso, M. A Comparative study on the microbial communities of Rhynchophorus ferrugineus (red palm weevil)-infected and healthy palm trees. Arab. J. Sci. Eng. 2022, 47, 6735–6746. [Google Scholar] [CrossRef]
- Kouamé, H.K.; Aké, M.D.F.; Assohoun, N.M.C.; Djè, M.K.; Djéni, N.T. Dynamics and species diversity of lactic acid bacteria involved in the spontaneous fermentation of various palm tree saps during palm wine tapping in Côte d’Ivoire. World J. Microbiol. Biotechnol. 2020, 36, 64. [Google Scholar] [CrossRef] [PubMed]
- Naknean, P.; Meenune, M.; Roudaut, G. Characterization of palm sap harvested in Songkhla province, Southern Thailand. Int. Food Res. J. 2010, 17, 977–986. [Google Scholar]
- He, M.; Jin, Y.; Zhou, R.; Zhao, D.; Zheng, J.; Wu, C. Dynamic succession of microbial community in Nongxiangxing daqu and microbial roles involved in flavor formation. Food Res. Int. 2022, 159, 111559. [Google Scholar] [CrossRef] [PubMed]
- Afolabi, F.T.; Owoola, A.T. Microbiological and physicochemical changes in palm wine subjected to spontaneous fermentation during storage. Int. J. Biotechnol. 2019, 8, 48–58. [Google Scholar] [CrossRef]
- Muñoz-Bernal, Ó.A.; Coria-Oliveros, A.J.; Vazquez-Flores, A.A.; de la Rosa, L.A.; Núnez-Gastélum, J.A.; Rodrigo-Garcia, J.; Ayala-Zavala, J.F.; Alvarez-Parrilla, E. Evolution of phenolic content, antioxidant capacity and phenolic profile during cold pre-fermentative maceration and subsequent fermentation of Cabernet Sauvignon red wine. S. Afr. J. Enol. Vitic. 2020, 41, 72–82. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Chukwuma, C.I.; Islam, M.S. Raffia palm (Raphia hookeri) wine: Qualitative sugar profile, functional chemistry, and antidiabetic properties. Food Biosci. 2019, 30, 100423. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Period (h) | Taal Toddy | Khejur Toddy | ||||
|---|---|---|---|---|---|---|
| Hooghly | Purulia | Bokaro | ||||
| Bacteria | Yeast | Bacteria | Yeast | Bacteria | Yeast | |
| 0 | Enterococcus faecalis | Saccharomyces cerevisiae | Lacticaseibacillus paracasei | Saccharomyces cerevisiae | Lacticaseibacillus paracasei | Saccharomyces cerevisiae |
| 24 | Lacticaseibacillus paracasei | Saccharomyces cerevisiae | Lactiplantibacillus plantarum | Saccharomyces cerevisiae | Enterococcus faecalis | Saccharomyces cerevisiae |
| 48 | Lactiplantibacillus plantarum | Saccharomyces cerevisiae | Lactiplantibacillus plantarum | Saccharomyces cerevisiae | Lactiplantibacillus plantarum | Saccharomyces cerevisiae |
| Place of Sample Collection | Successional Phase (h) | Total Alcohol % (v/v) | Proportion Distribution between Ethanol and Methanol | |
|---|---|---|---|---|
| Ethanol (%) | Methanol (%) | |||
| Hooghly | 0 | 2.80 ± 0.04 | 2.65 ± 0.04 | 0.15 ± 0.20 |
| 24 | 5.58 ± 0.07 | 5.28 ± 0.16 | 0.30 ± 0.01 | |
| 48 | 6.57 ± 0.12 | 6.19 ± 0.10 | 0.38 ± 0.00 | |
| Purulia | 0 | 1.34 ± 0.02 | 1.20 ± 0.01 | 0.14 ± 0.20 |
| 24 | 3.73 ± 0.02 | 3.36 ± 0.17 | 0.37 ± 0.02 | |
| 48 | 6.55 ± 0.01 | 6.50 ± 0.01 | 0.05 ± 0.35 | |
| Bokaro | 0 | 2.01 ± 0.02 | 1.93 ± 0.02 | 0.08 ± 0.20 |
| 24 | 5.11 ± 0.01 | 4.81 ± 0.01 | 0.30 ± 0.10 | |
| 48 | 6.60 ± 0.01 | 6.20 ± 0.20 | 0.40 ± 0.01 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, S.; Tamang, J.P. Fermentation Dynamics of Naturally Fermented Palm Beverages of West Bengal and Jharkhand in India. Fermentation 2023, 9, 301. https://doi.org/10.3390/fermentation9030301
Das S, Tamang JP. Fermentation Dynamics of Naturally Fermented Palm Beverages of West Bengal and Jharkhand in India. Fermentation. 2023; 9(3):301. https://doi.org/10.3390/fermentation9030301
Chicago/Turabian StyleDas, Souvik, and Jyoti Prakash Tamang. 2023. "Fermentation Dynamics of Naturally Fermented Palm Beverages of West Bengal and Jharkhand in India" Fermentation 9, no. 3: 301. https://doi.org/10.3390/fermentation9030301
APA StyleDas, S., & Tamang, J. P. (2023). Fermentation Dynamics of Naturally Fermented Palm Beverages of West Bengal and Jharkhand in India. Fermentation, 9(3), 301. https://doi.org/10.3390/fermentation9030301