
Effects of Lactic Acid Bacteria Additives on Fatty Acids, Amino Acids and Antioxidant Capacity of Leymus chinensis Silage during Aerobic Exposure
 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silage Preparation
2.2. Aerobic Stability
2.3. Antioxidant Capacity
2.4. Fatty Acid
2.5. Amino Acid
2.6. Statistical Analysis
3. Results
3.1. Silage Raw Material Characteristics
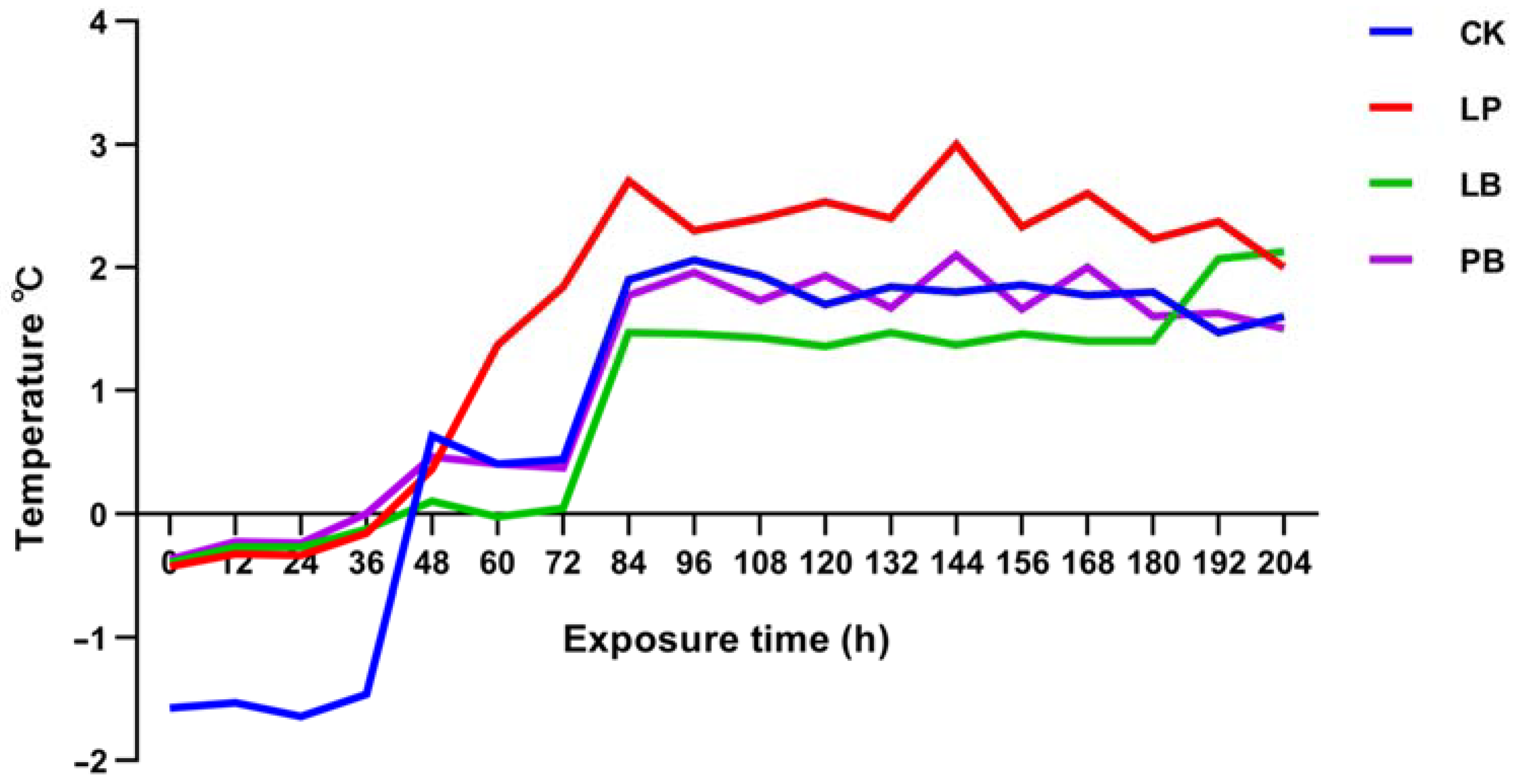
3.2. Temperature Difference between Silage and Room Temperature during Aerobic Exposure
3.3. Changes in Antioxidant Capacity of Silage during Aerobic Exposure
3.4. Changes in Fatty Acids during Aerobic Exposure
3.5. Changes in Amino Acids during Aerobic Exposure
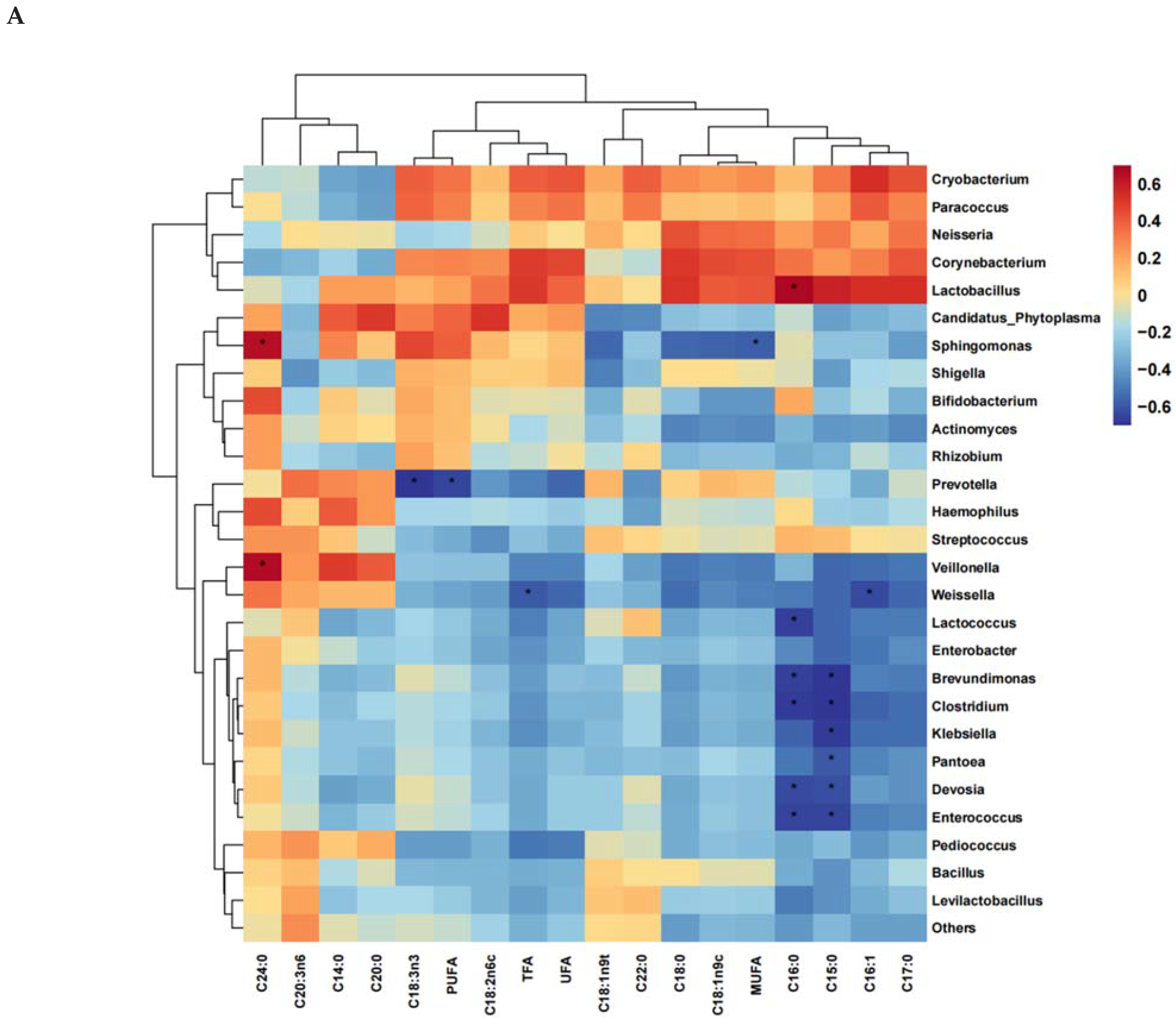
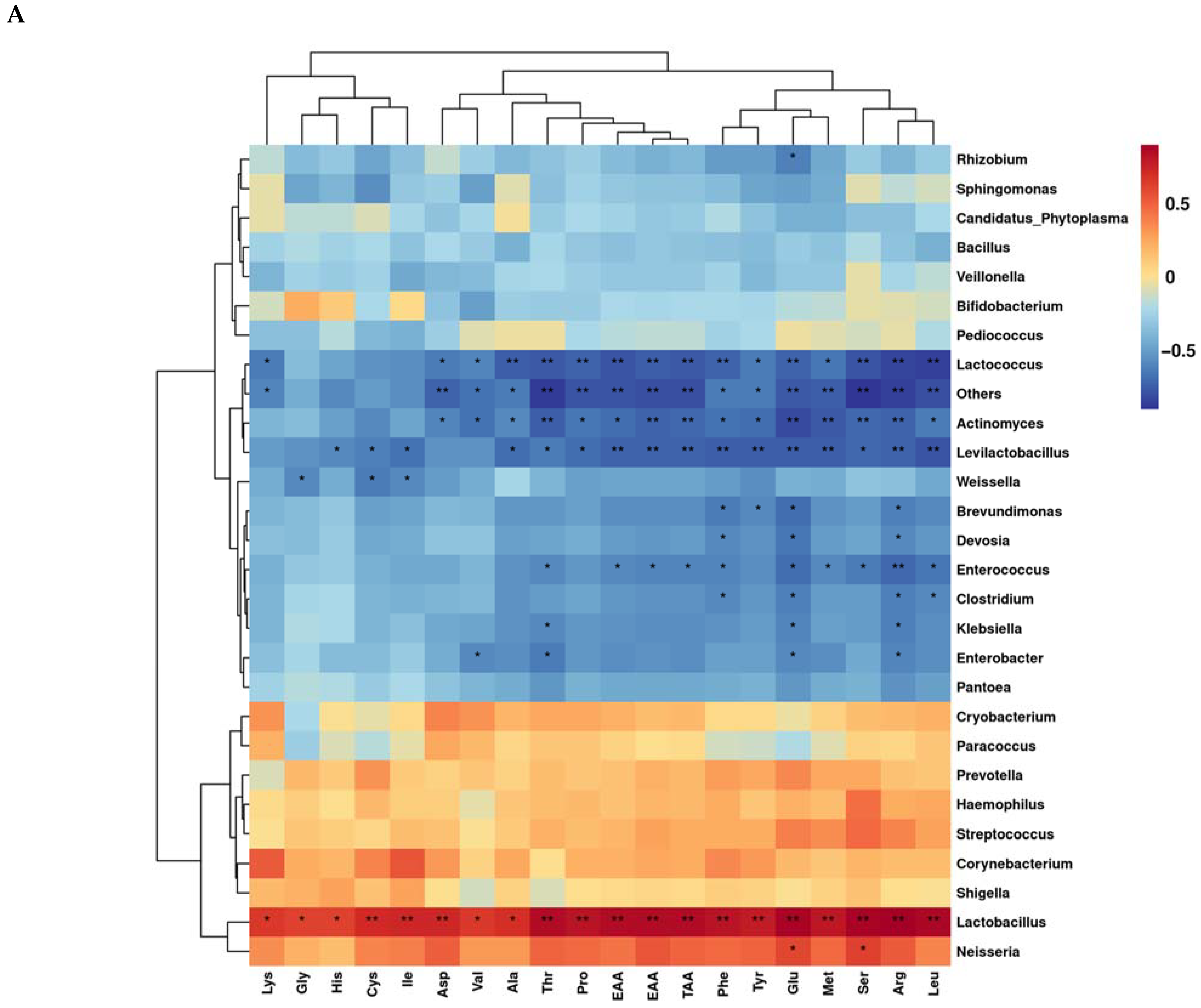
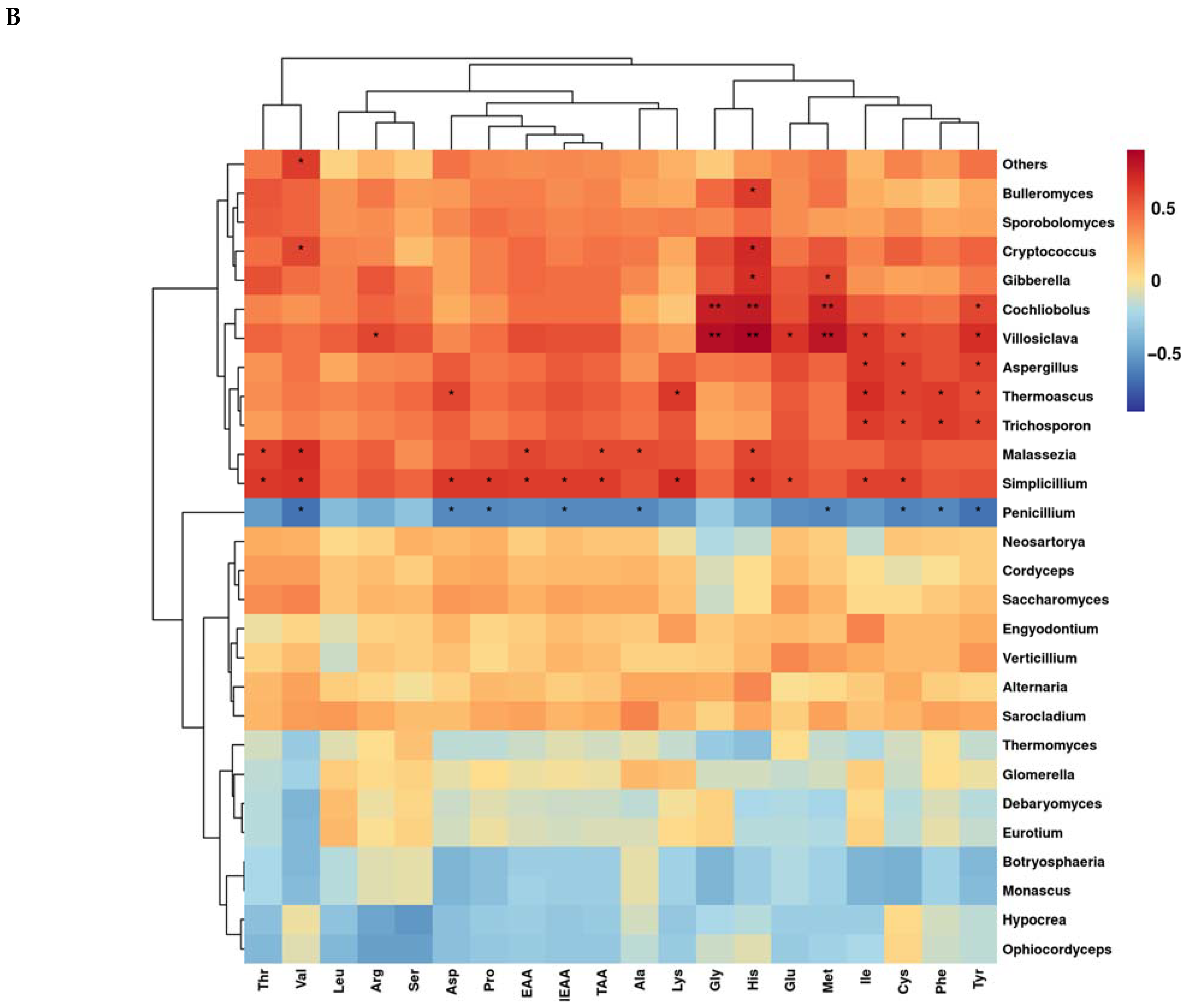
3.6. Correlation of Microbial Genera Levels with Fatty Acids and Amino Acids during Aerobic Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Chen, T.; Nan, Z.; Christensen, M.J. Cattle grazing alters the interaction of seed-borne fungi and two foliar pathogens of Leymus chinensis in a meadow steppe. Eur. J. Plant Pathol. 2019, 155, 207–218. [Google Scholar] [CrossRef]
- Chen, S.; Bobe, G.; Zimmerman, S.; Hammond, E.G.; Luhman, C.M.; Boylston, T.D.; Freeman, A.E.; Beitz, D.C. Physical and sensory properties of dairy products from cows with various milk fatty acid compositions. J. Agric. Food Chem. 2004, 52, 3422–3428. [Google Scholar] [CrossRef]
- Elgersma, A. Grazing increases the unsaturated fatty acid concentration of milk from grass-fed cows: A review of the contributing factors, challenges and future perspectives. Eur. J. Lipid. Sci. Technol. 2015, 117, 1345–1369. [Google Scholar] [CrossRef]
- da Silva Lima, E.; Valente, T.; de Oliveira Roça, R.; Cezário, A.S.; dos Santos, W.; Deminicis, B.B.; Ribeiro, J.C. Effect of whole cottonseed or protected fat dietary additives on carcass characteristics and meat quality of beef cattle: A review. J. Agric. Sci. 2017, 9, 175–189. [Google Scholar]
- Pfeuffer, M.; Watzl, B. Nutrition and health aspects of milk and dairy products and their ingredients. Ernahr. Umschau. 2018, 65, 22–33. [Google Scholar]
- Wang, B.; Zhao, X.; Li, Z.; Luo, H.; Zhang, H.; Guo, Y.; Zhang, C.; Ma, Q. Changes of Metabolites and Gene Expression under Different Feeding Systems Associated with Lipid Metabolism in Lamb Meat. Foods 2021, 10, 2612. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.; Levine, R. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino. Acids 2003, 25, 207–218. [Google Scholar] [CrossRef]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V. Mechanisms of oxidative processes in meat and toxicity induced by postprandial degradation products: A review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 96–123. [Google Scholar] [CrossRef]
- Höhn, A.; König, J.; Grune, T. Protein oxidation in aging and the removal of oxidized proteins. J. Proteomics. 2013, 92, 132–159. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Hussin, G.; Ramli, A.; Miah, G. Fermentation quality and additives: A case of rice straw silage. Biomed. Res. Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kung, L. Silage fermentation and additives. Lat. Am. A Anim. Pro. 2018, 26, 3–4. [Google Scholar]
- Salami, S.; Guinguina, A.; Agboola, J.; Omede, A.; Agbonlahor, E.; Tayyab, U. In vivo and postmortem effects of feed antioxidants in livestock: A review of the implications on authorization of antioxidant feed additives. Animal 2016, 10, 1375–1390. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Xu, D.; Li, F.; Bai, J.; Su, R. Current approaches on the roles of lactic acid bacteria in crop silage. Microb. Biotechnol. 2023, 16, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, L.; Zhou, B.; Zou, C. Research process in silage microorganism and its effect on silage aerobic stability. Chin. J. Anim. Nutr. 2019, 31, 82–89. [Google Scholar]
- Mu, L.; Xie, Z.; Hu, L.; Chen, G.; Zhang, Z. Cellulase interacts with Lactobacillus plantarum to affect chemical composition, bacterial communities, and aerobic stability in mixed silage of high-moisture amaranth and rice straw. Bioresour. Technol. 2020, 315, 123772. [Google Scholar] [CrossRef]
- Martin, B.; Verdier-Metz, I.; Buchin, S.; Hurtaud, C.; Coulon, J.-B. How do the nature of forages and pasture diversity influence the sensory quality of dairy livestock products? Anim. Sci. 2005, 81, 205–212. [Google Scholar] [CrossRef]
- Figueiredo, R.; Rodrigues, A.I.; do Céu Costa, M. Volatile composition of red clover (Trifolium pratense L.) forages in Portugal: The influence of ripening stage and ensilage. Food Chem. 2007, 104, 1445–1453. [Google Scholar] [CrossRef]
- Ranjit, N.K.; Kung Jr, L. The effect of Lactobacillus buchneri, Lactobacillus plantarum, or a chemical preservative on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2000, 83, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Untea, A.; Lupu, A.; Saracila, M.; Panaite, T. Comparison of ABTS, DPPH, phosphomolybdenum assays for estimating antioxidant activity and phenolic compounds in five different plant extracts. Bull. UASVM Anim. Sci. Bio. 2018, 75, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, Y.; Lu, Q.; Sun, L.; Du, S.; Liu, T.; Hou, M.; Ge, G.; Wang, Z.; Jia, Y. Effects of lactic acid bacteria additives on the quality, volatile chemicals and microbial community of leymus chinensis silage during aerobic exposure. Front. Microbiol. 2022, 13, 938153. [Google Scholar] [CrossRef]
- Liu, Q.; Dong, Z.; Shao, T. Effect of additives on fatty acid profile of high moisture alfalfa silage during ensiling and after exposure to air. Anim. Feed Sci. Technol. 2018, 236, 29–38. [Google Scholar] [CrossRef]
- Guo, X.; Ding, W.; Han, J.; Zhou, H. Characterization of protein fractions and amino acids in ensiled alfalfa treated with different chemical additives. Anim. Feed Sci. Technol. 2008, 142, 89–98. [Google Scholar] [CrossRef]
- Muck, R.; Nadeau, E.; McAllister, T.; Contreras-Govea, F.; Santos, M.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- He, L.; Zhou, W.; Wang, C.; Yang, F.; Chen, X.; Zhang, Q. Effect of cellulase and Lactobacillus casei on ensiling characteristics, chemical composition, antioxidant activity, and digestibility of mulberry leaf silage. J. Dairy Sci. 2019, 102, 9919–9931. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Bernardes, T.F.; Copani, G.; Fortunati, P.; Giuberti, G.; Bruschi, S.; Bryan, K.A.; Nielsen, N.G.; Witt, K.L.; Masoero, F. Effect of inoculation with Lactobacillus buchneri LB1819 and Lactococcus lactis O224 on fermentation and mycotoxin production in maize silage compacted at different densities. Anim. Feed Sci. Technol. 2018, 246, 36–45. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123600. [Google Scholar] [CrossRef]
- Amiri, S.; Moghanjougi, Z.M.; Bari, M.R.; Khaneghah, A.M. Natural protective agents and their applications as bio-preservatives in the food industry: An overview of current and future applications. Ital. J. Food Sci. 2021, 33, 55–68. [Google Scholar] [CrossRef]
- Kalogianni, A.I.; Lazou, T.; Bossis, I.; Gelasakis, A.I. Natural phenolic compounds for the control of oxidation, bacterial spoilage, and foodborne pathogens in meat. Foods 2020, 9, 794. [Google Scholar] [CrossRef]
- Mugabe, W.; Shao, T.; Li, J.; Dong, Z.; Yuan, X. Effect of hexanoic acid, Lactobacillus plantarum and their combination on the aerobic stability of napier grass silage. J. Appl. Microbiol. 2020, 129, 823–831. [Google Scholar] [CrossRef]
- Zhang, Y.; Ke, W.; Bai, J.; Li, F.; Xu, D.; Ding, Z.; Guo, X. The effect of Pediococcus acidilactici J17 with high-antioxidant activity on antioxidant, α-tocopherol, β-carotene, fatty acids, and fermentation profiles of alfalfa silage ensiled at two different dry matter contents. Anim. Feed Sci. Technol. 2020, 268, 114614. [Google Scholar] [CrossRef]
- Clarke, H.J.; McCarthy, W.P.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. Oxidative quality of dairy powders: Influencing factors and analysis. Foods 2021, 10, 2315. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Cone, J.; Hendriks, W. Stability of fatty acids in grass and maize silages after exposure to air during the feed out period. Anim. Feed Sci. Technol. 2009, 154, 183–192. [Google Scholar] [CrossRef]
- Winnik, S.; Lohmann, C.; Richter, E.K.; Schäfer, N.; Song, W.-L.; Leiber, F.; Mocharla, P.; Hofmann, J.; Klingenberg, R.; Boren, J. Dietary α-linolenic acid diminishes experimental atherogenesis and restricts T cell-driven inflammation. Eur. heart. J. 2011, 32, 2573–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, S.; Cabrita, A.; Jerónimo, E.; Bessa, R.; Fonseca, A. Effect of ensiling and silage additives on fatty acid composition of ryegrass and corn experimental silages. J. Anim. Sci. 2011, 89, 2537–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibasaki-Kitakawa, N.; Kato, H.; Takahashi, A.; Yonemoto, T. Oxidation kinetics of β-carotene in oleic acid solvent with addition of an antioxidant, α-tocopherol. J. Am. Oil Chem. Soc. 2004, 81, 389–394. [Google Scholar] [CrossRef]
- Ma, J.; Dai, H.; Liu, H.; Du, W. Effects of Cutting Stages and Additives on the Fermentation Quality of Triticale, Rye and Oat Silage in Qinghai-Tibet Plateau. Agronomy 2022, 12, 3113. [Google Scholar] [CrossRef]
- Rathod, N.B.; Ranveer, R.C.; Bhagwat, P.K.; Ozogul, F.; Benjakul, S.; Pillai, S.; Annapure, U.S. Cold plasma for the preservation of aquatic food products: An overview. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4407–4425. [Google Scholar] [CrossRef]
- Dong, J.; Li, S.; Chen, X.; Sun, Z.; Sun, Y.; Zhen, Y.; Qin, G.; Wang, T.; Demelash, N.; Zhang, X. Effects of Lactiplantibacillus plantarum inoculation on the quality and bacterial community of whole-crop corn silage at different harvest stages. Chem. Biol. Technol. Agric. 2022, 9, 57. [Google Scholar] [CrossRef]
- Lim, Y.H.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdullah, N. Comparative studies of versatile extracellular proteolytic activities of lactic acid bacteria and their potential for extracellular amino acid productions as feed supplements. J. Anim. Sci. Biotechnol. 2019, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Mateo, R.D.; Yin, Y.-L.; Wu, G. Functional amino acids and fatty acids for enhancing production performance of sows and piglets. Asian-Australas. J. Anim. Sci. 2007, 20, 295–306. [Google Scholar] [CrossRef]
- Jiao, Y.; Li, X.; Kim, I.H. Changes in growth performance, nutrient digestibility, immune blood profiles, fecal microbial and fecal gas emission of growing pigs in response to zinc aspartic acid chelate. Asian-Australas. J. Anim. Sci. 2020, 33, 597. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, S.; Wu, G. Amino acid metabolism in the kidneys: Nutritional and physiological significance. Amino. Acids. Nutr. Health Amino Acids Syst. Funct. Health 2020, 1265, 71–95. [Google Scholar]
- Qi, B.; Wang, J.; Hu, M.; Ma, Y.; Wu, S.; Qi, G.; Qiu, K.; Zhang, H. Influences of beta-alanine and l-histidine supplementation on growth performance, meat quality, carnosine content, and mRNA expression of carnosine-related enzymes in broilers. Animals 2021, 11, 2265. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.Y.; Beidler, J.; Hooshmand, S.; Figueroa, A.; Kern, M. Watermelon and L-arginine consumption improve serum lipid profile and reduce inflammation and oxidative stress by altering gene expression in rats fed an atherogenic diet. Nutr. Res. 2018, 58, 46–54. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc Rhoads, J.; Carey Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino. Acids. 2009, 37, 153–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, W.; Ding, W.; Xu, D.; Shah, M.N.; Zhang, P.; Guo, X. Influences of addition of malic acid or citric acid, Lactobacillus plantarum and their mixtures on fermentation quality, proteolysis and fatty acid composition of ensiled alfalfa. Arch. Anim. Nutr. 2018, 72, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Zong, C.; Wu, Q.; Wu, A.; Chen, S.; Dong, D.; Zhao, J.; Shao, T.; Liu, Q. Exploring the diversity mechanism of fatty acids and the loss mechanisms of polyunsaturated fatty acids and fat-soluble vitamins in alfalfa silage using different additives. Anim. Feed. Sci. Technol. 2021, 280, 115044. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Mitra, M.; Maiti, M.K. Recent advances in lipid metabolic engineering of oleaginous yeasts. Biotechnol. Adv. 2021, 53, 107722. [Google Scholar] [CrossRef]
- de Andrade Silva, C.A.; da Silva, P.G.P.; da Silva, G.F.A.; Dantas, D.P.; Leite, R.S.R.; Fonseca, G.G. Biotransformation of fruit residues via solid state bioprocess using Lichtheimia ramosa. SN Appl. Sci. 2020, 2, 861. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Yuan, R.; Weng, S.; Wang, G.; Xiong, Z.; Zhang, H.; Song, X.; Liu, W.; Ai, L. Proteolysis, lipolysis, texture and sensory properties of cheese ripened by Monascus fumeus. Food Res. Int. 2020, 137, 109657. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Tang, J.; Zhang, Z.; Wu, Z.; Zhong, A.; Li, Z.; Wang, Y. Correlation between flavor compounds and microorganisms of Chaling natural fermented red sufu. LWT 2022, 154, 112873. [Google Scholar] [CrossRef]
- Zhao, S.; Yang, F.; Wang, Y.; Fan, X.; Feng, C.; Wang, Y. Dynamics of fermentation parameters and bacterial community in high-moisture alfalfa silage with or without lactic acid bacteria. Microorganisms 2021, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- da Silva, T.C.; da Silva, L.D.; Santos, E.M.; Oliveira, J.S.; Perazzo, A.F. Importance of the fermentation to produce high-quality silage. Ferment. Proc. 2017, 2, 1–20. [Google Scholar]
- Neis, E.P.; Dejong, C.H.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Jiang, D.; Zheng, M.; Tian, P.; Zheng, M.; Xu, C. Microbial community dynamics during alfalfa silage with or without clostridial fermentation. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Dong, Z.; Li, J.; Wang, S.; Dong, D.; Shao, T. Diurnal Variation of Epiphytic Microbiota: An Unignorable Factor Affecting the Anaerobic Fermentation Characteristics of Sorghum-Sudangrass Hybrid Silage. Microbiol. Spectrum. 2022, 11, e03404–e03422. [Google Scholar] [CrossRef]
- Gallo, A.; Ghilardelli, F.; Atzori, A.S.; Zara, S.; Novak, B.; Faas, J.; Fancello, F. Co-occurrence of regulated and emerging mycotoxins in corn silage: Relationships with fermentation quality and bacterial communities. Toxins 2021, 13, 232. [Google Scholar] [CrossRef]
- Sommer, B.; Overy, D.P.; Haltli, B.; Kerr, R.G. Secreted lipases from Malassezia globosa: Recombinant expression and determination of their substrate specificities. Microbiology 2016, 162, 1069–1079. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Lima, N. Thermophilic fungi to dominate aflatoxigenic/mycotoxigenic fungi on food under global warming. Int. J. Environ. Res. Public Health 2017, 14, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wali, A.; Hou, J.; Tsuruta, T.; Nishino, N. Bacterial and fungal microbiota of total mixed ration silage stored at various temperatures. J. Appl. Microbiol. 2022, 133, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Xue, M.; Shen, Z.; Jia, X.; Hou, X.; Lai, D.; Zhou, L. Phytotoxic secondary metabolites from fungi. Toxins 2021, 13, 261. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Sample | SEM | |
|---|---|---|---|
| Fatty Acids (%) | TFA | 1.4839 | 0.0120 |
| SFA | 0.4843 | 0.0056 | |
| PUFA | 0.7044 | 0.0179 | |
| MUFA | 0.2952 | 0.0022 | |
| C14:0 | 0.0509 | 0.0005 | |
| C15:0 | 0.0056 | 0.0000 | |
| C16:0 | 0.3602 | 0.0055 | |
| C16:1 | 0.0423 | 0.0004 | |
| C17:0 | 0.0048 | 0.0003 | |
| C18:0 | 0.0433 | 0.0008 | |
| C18:1n9t | 0.0245 | 0.0001 | |
| C18:1n9c | 0.2284 | 0.0018 | |
| C18:2n6c | 0.2329 | 0.0045 | |
| C20:0 | 0.0161 | 0.0003 | |
| C18:3n3 | 0.4715 | 0.0222 | |
| C22:0 | 0.0000 | 0.0000 | |
| C20:3n6 | 0.0000 | 0.0000 | |
| C24:0 | 0.0035 | 0.0003 | |
| Amino Acids (%) | TAA | 12.9613 | 0.0154 |
| EAA | 6.1690 | 0.0570 | |
| NEAA | 6.7923 | 0.0426 | |
| Arg * | 0.7243 | 0.0146 | |
| His * | 0.4057 | 0.0215 | |
| Ile * | 0.5631 | 0.0109 | |
| Leu * | 1.1381 | 0.0260 | |
| Lys * | 0.7260 | 0.0116 | |
| Met * | 0.1451 | 0.0021 | |
| Phe * | 0.7991 | 0.0116 | |
| Thr * | 0.6157 | 0.0184 | |
| Val * | 1.0517 | 0.0105 | |
| Ala | 1.1814 | 0.0217 | |
| Asp | 1.7258 | 0.0614 | |
| Cys | 0.0894 | 0.0018 | |
| Glu | 1.4910 | 0.0425 | |
| Gly | 0.6557 | 0.0145 | |
| Pro | 0.6928 | 0.0085 | |
| Ser | 0.5414 | 0.0105 | |
| Tyr | 0.4147 | 0.0074 | |
| Antioxidant capacity (%) | Total Phenol | 0.3732 | 0.0146 |
| Inhibition Rate | 72.9067 | 0.4491 |
| Items | Time (d) | Treatments (%) | |||
|---|---|---|---|---|---|
| CK | LP | LB | PB | ||
| Total Phenol | 0 | 0.1772 ± 0.0048 Da | 0.3213 ± 0.0093 Ba | 0.3358 ± 0.01 Aa | 0.3037 ± 0.0048 Ca |
| 4 | 0.1015 ± 0.0065 Cb | 0.1907 ± 0.0082 Bb | 0.2176 ± 0.0065 Ab | 0.2083 ± 0.0078 Ab | |
| 8 | 0.0693 ± 0.0082 Cc | 0.0953 ± 0.0078 Bc | 0.1741 ± 0.0036 Ac | 0.1668 ± 0.0048 Ac | |
| Inhibition Rate | 0 | 65.616 ± 0.0955 Ca | 68.6406 ± 0.1988 Ba | 71.0602 ± 0.2527 Aa | 68.4814 ± 0.4963 Ba |
| 4 | 64.7883 ± 0.526 Ca | 63.6103 ± 0.4963 Db | 67.972 ± 0.4307 Ab | 66.794 ± 0.2918 Bb | |
| 8 | 63.5466 ± 0.526 Cb | 62.8462 ± 0.5054 Cb | 66.9214 ± 0.1459 Ac | 65.425 ± 0.2527 Bc | |
| Items | Time (d) | Treatments (%) | |||
|---|---|---|---|---|---|
| CK | LP | LB | PB | ||
| TFA | 0 | 0.9705 ± 0.1504 Aa | 1.0861 ± 0.1903 Aa | 1.2443 ± 0.0871 Aa | 1.0652 ± 0.1592 Aa |
| 4 | 0.9042 ± 0.0842 Aab | 0.8636 ± 0.0821 Aab | 1.1991 ± 0.2884 Aa | 0.9391 ± 0.2053 Aa | |
| 8 | 0.7358 ± 0.0831 Cb | 0.8214 ± 0.0569 BCb | 1.1802 ± 0.0328 Aa | 0.8959 ± 0.1191 Ba | |
| SFA | 0 | 0.3917 ± 0.0805 Aa | 0.4413 ± 0.1037 Aa | 0.422 ± 0.0277 Aa | 0.3962 ± 0.0264 Aa |
| 4 | 0.3783 ± 0.0374 Aa | 0.356 ± 0.0263 Aa | 0.4033 ± 0.0751 Aa | 0.3689 ± 0.076 Aa | |
| 8 | 0.2948 ± 0.0383 Ba | 0.3686 ± 0.0115 Aa | 0.3885 ± 0.0188 Aa | 0.4212 ± 0.0589 Aa | |
| PUFA | 0 | 0.3967 ± 0.0516 Ba | 0.3528 ± 0.0904 Ba | 0.6464 ± 0.0884 Aa | 0.5365 ± 0.1556 ABa |
| 4 | 0.3631 ± 0.0323 Ba | 0.3551 ± 0.0184 Ba | 0.5926 ± 0.1448 Aa | 0.4118 ± 0.0667 Ba | |
| 8 | 0.3558 ± 0.0448 Ba | 0.3681 ± 0.0582 Ba | 0.7014 ± 0.0165 Aa | 0.374 ± 0.0364 B | |
| MUFA | 0 | 0.1821 ± 0.1201 Aa | 0.2921 ± 0.1585 Aa | 0.176 ± 0.0632 Aa | 0.1324 ± 0.0204 Aa |
| 4 | 0.1627 ± 0.0452 Aa | 0.1525 ± 0.0426 Aab | 0.2032 ± 0.0808 Aa | 0.1584 ± 0.0767 Aa | |
| 8 | 0.0852 ± 0.0013 A | 0.0847 ± 0.0116 Ab | 0.0903 ± 0.0143 Aa | 0.1007 ± 0.0246 Aa | |
| C14:0 | 0 | 0.0415 ± 0.0045 Aa | 0.0287 ± 0.0061 Bb | 0.0371 ± 0.0019 Aa | 0.0267 ± 0.0034 Bc |
| 4 | 0.0262 ± 0.0018 Bb | 0.0354 ± 0.0019 ABab | 0.0253 ± 0.0062 Bb | 0.0399 ± 0.0085 Ab | |
| 8 | 0.0277 ± 0.0022 Cb | 0.0422 ± 0.0065 Ba | 0.0425 ± 0.0023 Ba | 0.0524 ± 0.0034 Aa | |
| C15:0 | 0 | 0.002 ± 0.0004 Aa | 0.0022 ± 0.0006 Aa | 0.002 ± 0.0003 Aa | 0.002 ± 0.0006 Aa |
| 4 | 0.0022 ± 0.0006 Aa | 0.002 ± 0.0001 Aa | 0.0022 ± 0.0004 Aa | 0.0018 ± 0.0002 Aa | |
| 8 | 0.0015 ± 0.0004 Aa | 0.0018 ± 0 Aa | 0.002 ± 0.0001 Aa | 0.0022 ± 0.0006 Aa | |
| C16:0 | 0 | 0.2778 ± 0.0448 Aa | 0.3041 ± 0.0513 Aa | 0.2976 ± 0.0137 Aa | 0.2981 ± 0.0115 Aa |
| 4 | 0.2749 ± 0.0171 Aa | 0.2544 ± 0.0114 Aa | 0.2899 ± 0.0473 Aa | 0.2548 ± 0.0468 Aa | |
| 8 | 0.2232 ± 0.0327 Ba | 0.2726 ± 0.0084 ABa | 0.2898 ± 0.013 Aa | 0.3079 ± 0.041 Aa | |
| C16:1 | 0 | 0.025 ± 0.0081 Aa | 0.0299 ± 0.0075 Aa | 0.0271 ± 0.001 Aa | 0.0292 ± 0.002 Aa |
| 4 | 0.0267 ± 0.0041 Aa | 0.0223 ± 0.0022 Aa | 0.0324 ± 0.0091 Aa | 0.0225 ± 0.004 Aa | |
| 8 | 0.0179 ± 0.0019 Ba | 0.0214 ± 0.0008 ABa | 0.0247 ± 0.0017 Aa | 0.0251 ± 0.0044 Aa | |
| C17:0 | 0 | 0.0043 ± 0.0022 Aa | 0.0064 ± 0.0043 Aa | 0.0046 ± 0.0012 Aab | 0.0038 ± 0.0009 Aa |
| 4 | 0.0044 ± 0.0011 ABa | 0.0035 ± 0.0007 Ba | 0.0057 ± 0.0006 Aa | 0.0037 ± 0.0011 Ba | |
| 8 | 0.0018 ± 0.0016 Aa | 0.003 ± 0.0002 Aa | 0.0032 ± 0.0002 Ab | 0.0036 ± 0.0009 Aa | |
| C18:0 | 0 | 0.0551 ± 0.0302 Aa | 0.0934 ± 0.0479 Aa | 0.0684 ± 0.0156 Aa | 0.0529 ± 0.0117 Aa |
| 4 | 0.0591 ± 0.0149 Aa | 0.0496 ± 0.0166 Aa | 0.0694 ± 0.0244 Aa | 0.0563 ± 0.0206 Aa | |
| 8 | 0.0311 ± 0.0022 Aa | 0.0365 ± 0.0035 Aa | 0.039 ± 0.0052 Aa | 0.0413 ± 0.0125 Aa | |
| C18:1n9t | 0 | 0.0107 ± 0.0046 Aab | 0.0114 ± 0.0065 Aa | 0.0048 ± 0.0028 Aab | 0.0072 ± 0.0038 Aa |
| 4 | 0.0128 ± 0.0028 Aa | 0.0076 ± 0.0015 Aa | 0.01 ± 0.0043 Aa | 0.0072 ± 0.0032 Aa | |
| 8 | 0.0055 ± 0.0014 Bb | 0.0071 ± 0.0007 Ba | 0.0024 ± 0.0005 Cb | 0.0108 ± 0.0019 Aa | |
| C18:1n9c | 0 | 0.1464 ± 0.1074 Aa | 0.2508 ± 0.1468 Aab | 0.1441 ± 0.0595 Aa | 0.096 ± 0.0192 Aa |
| 4 | 0.1232 ± 0.0389 Aa | 0.1226 ± 0.0407 Aa | 0.1607 ± 0.0723 Aa | 0.1287 ± 0.07 Aa | |
| 8 | 0.0618 ± 0.0034 Aa | 0.0562 ± 0.0107 Ab | 0.0631 ± 0.013 Aa | 0.0649 ± 0.0185 Aa | |
| C18:2n6c | 0 | 0.1679 ± 0.0357 ABa | 0.1395 ± 0.0549 Ba | 0.2422 ± 0.0484 Aa | 0.1706 ± 0.0451 ABa |
| 4 | 0.1245 ± 0.0068 Ba | 0.1544 ± 0.02 ABa | 0.1791 ± 0.031 Aa | 0.171 ± 0.028 Aa | |
| 8 | 0.1235 ± 0.0157 Ba | 0.1267 ± 0.0102 Ba | 0.2194 ± 0.0105 Aa | 0.1322 ± 0.0086 Ba | |
| C18:3n3 | 0 | 0.2258 ± 0.0222 Ba | 0.211 ± 0.0361 Ba | 0.4042 ± 0.0483 Aa | 0.3645 ± 0.1117 Aa |
| 4 | 0.2354 ± 0.0252 Ba | 0.199 ± 0.0134 Ba | 0.412 ± 0.1139 Aa | 0.2389 ± 0.0396 Ba | |
| 8 | 0.2306 ± 0.0302 Ba | 0.2386 ± 0.0483 Ba | 0.4814 ± 0.0056 Aa | 0.2392 ± 0.0301 Ba | |
| C20:0 | 0 | 0.0102 ± 0.0017 ABa | 0.0064 ± 0.0016 Cb | 0.0113 ± 0.0026 Aa | 0.0074 ± 0.0014 BCb |
| 4 | 0.0071 ± 0.0006 Ba | 0.0103 ± 0.0004 Aa | 0.0068 ± 0.0016 Bb | 0.0107 ± 0.0006 Aa | |
| 8 | 0.0078 ± 0.0019 Ba | 0.0097 ± 0.0004 ABa | 0.0098 ± 0.0006 ABab | 0.0113 ± 0.0022 Aa | |
| C20:3n6 | 0 | 0.0031 ± 0.0004 Aa | 0.0023 ± 0.0006 ABa | 0 | 0.0014 ± 0.0014 Ba |
| 4 | 0.0032 ± 0.0006 Aa | 0.0017 ± 0.0015 Aa | 0.0016 ± 0.0014 Aa | 0.002 ± 0.0003 Aa | |
| 8 | 0.0016 ± 0.0014 Aa | 0.0028 ± 0.0001 Aa | 0.0005 ± 0.0009 Aa | 0.0027 ± 0.0023 Aa | |
| C22:0 | 0 | 0 | 0 | 0 | 0.0036 ± 0.0062 |
| 4 | 0.0039 ± 0.0067 | 0 | 0.0029 ± 0.0049 | 0 | |
| 8 | 0 | 0 | 0 | 0 | |
| C24:0 | 0 | 0.0008 ± 0.0014 ABa | 0 | 0.0011 ± 0.001 ABa | 0.0018 ± 0.0002 Aa |
| 4 | 0.0006 ± 0.001 Aa | 0.0008 ± 0.0014 ABb | 0.0011 ± 0.001 Aa | 0.0016 ± 0.0014 Aa | |
| 8 | 0.0017 ± 0.0003 Ba | 0.0027 ± 0.0005 Aa | 0.0023 ± 0.0004 ABa | 0.0025 ± 0.0005 ABa | |
| Items | Time (d) | Treatments (%) | |||
|---|---|---|---|---|---|
| CK | LP | LB | PB | ||
| TAA | 0 | 11.7488 ± 0.1357 Ca | 14.0573 ± 0.0045 Aa | 13.3961 ± 0.0775 Ba | 13.4761 ± 0.1161 Ba |
| 4 | 10.5513 ± 0.1429 Cb | 12.6188 ± 0.3121 ABb | 12.9406 ± 0.1365 Ab | 12.5660 ± 0.0550 Bb | |
| 8 | 9.8805 ± 0.1025 Dc | 11.1348 ± 0.1034 Cc | 12.0638 ± 0.0189 Ac | 11.7564 ± 0.2185 Bc | |
| EAA | 0 | 5.6304 ± 0.0154 Ca | 6.5886 ± 0.1274 Aa | 6.39 ± 0.0178 Ba | 6.4056 ± 0.0625 Ba |
| 4 | 4.9617 ± 0.0523 Cb | 5.9481 ± 0.1222 Bb | 6.1855 ± 0.1317 Ab | 5.8945 ± 0.0836 Bb | |
| 8 | 4.7032 ± 0.0427 Cc | 5.2953 ± 0.0510 Bc | 5.7079 ± 0.0665 Ac | 5.6156 ± 0.1282 Ac | |
| NEAA | 0 | 6.1184 ± 0.1491 Ca | 7.4687 ± 0.1320 Aa | 7.0061 ± 0.0721 Ba | 7.0705 ± 0.0845 Ba |
| 4 | 5.5896 ± 0.0909 Bb | 6.6707 ± 0.1904 Ab | 6.7552 ± 0.0495 Ab | 6.6716 ± 0.1385 Ab | |
| 8 | 5.1772 ± 0.0598 Dc | 5.8395 ± 0.0536 Cc | 6.3559 ± 0.0661 Ac | 6.1408 ± 0.0942 Bc | |
| Arg * | 0 | 0.5111 ± 0.0103 Ba | 0.6931 ± 0.0258 Aa | 0.6659 ± 0.0243 Aa | 0.7021 ± 0.0056 Aa |
| 4 | 0.4678 ± 0.0164 Cb | 0.6065 ± 0.0062 Bb | 0.6395 ± 0.0244 Aa | 0.5972 ± 0.0149 Bb | |
| 8 | 0.3899 ± 0.0073 Cc | 0.5620 ± 0.0168 Bc | 0.5916 ± 0.0065 Ab | 0.5704 ± 0.0110 Bc | |
| His * | 0 | 0.3469 ± 0.0127 Ca | 0.3761 ± 0.0167 Ba | 0.3882 ± 0.0046 Ba | 0.4186 ± 0.0109 Aa |
| 4 | 0.3114 ± 0.0100 Bb | 0.3431 ± 0.0066 Ab | 0.3493 ± 0.0041 Ab | 0.3362 ± 0.0140 Ab | |
| 8 | 0.3215 ± 0.0096 Bb | 0.3230 ± 0.0060 ABb | 0.3381 ± 0.0081 Ab | 0.3338 ± 0.0071 ABb | |
| Ile * | 0 | 0.4947 ± 0.0161 Ca | 0.5784 ± 0.0254 Aa | 0.5358 ± 0.0206 Ba | 0.5440 ± 0.0128 ABa |
| 4 | 0.4717 ± 0.0058 Bb | 0.4956 ± 0.0084 ABb | 0.5095 ± 0.0318 Aa | 0.4893 ± 0.0114 ABb | |
| 8 | 0.4513 ± 0.0091 Bb | 0.4580 ± 0.0169 Bc | 0.5133 ± 0.0166 Aa | 0.5038 ± 0.0184 Ab | |
| Leu * | 0 | 1.0950 ± 0.0508 Ba | 1.2528 ± 0.0149 Aa | 1.2301 ± 0.0377 Aa | 1.2530 ± 0.0460 Aa |
| 4 | 0.9379 ± 0.0221 Bb | 1.1472 ± 0.0421 Ab | 1.2167 ± 0.0477 Aa | 1.1412 ± 0.0424 Ab | |
| 8 | 0.9096 ± 0.0050 Cb | 1.0653 ± 0.0470 Bc | 1.1660 ± 0.0438 Aa | 1.1922 ± 0.0312 Aab | |
| Lys * | 0 | 0.6657 ± 0.0195 Ba | 0.7865 ± 0.0108 Aa | 0.7665 ± 0.0342 Aa | 0.6877 ± 0.0384 Ba |
| 4 | 0.6216 ± 0.0130 Cb | 0.6890 ± 0.0277 Bb | 0.7483 ± 0.0211 Aa | 0.6849 ± 0.0069 Ba | |
| 8 | 0.6106 ± 0.0214 Cb | 0.6483 ± 0.0206 Bb | 0.7180 ± 0.0106 Aa | 0.6676 ± 0.0095 Ba | |
| Met * | 0 | 0.1138 ± 0.0029 Ca | 0.1394 ± 0.0087 Aa | 0.1270 ± 0.0056 Ba | 0.1500 ± 0.0041 Aa |
| 4 | 0.1037 ± 0.0030 Ab | 0.1233 ± 0.0060 Ab | 0.1227 ± 0.0059 Aab | 0.1213 ± 0.0267 Aab | |
| 8 | 0.0921 ± 0.0005 Bc | 0.1063 ± 0.0083 ABc | 0.1132 ± 0.0067 Ab | 0.1096 ± 0.0134 Ab | |
| Phe * | 0 | 0.7860 ± 0.0185 Ba | 0.9297 ± 0.0296 Aa | 0.8080 ± 0.0251 Ba | 0.8350 ± 0.0305 Ba |
| 4 | 0.6555 ± 0.0103 Cb | 0.7436 ± 0.0251 Bb | 0.7666 ± 0.0245 ABab | 0.7913 ± 0.0245 Aab | |
| 8 | 0.5844 ± 0.0033 Cc | 0.6846 ± 0.0251 Bc | 0.7608 ± 0.0130 Ab | 0.7239 ± 0.0483 ABb | |
| Thr * | 0 | 0.5222 ± 0.0114 Ba | 0.6331 ± 0.0329 Aa | 0.6669 ± 0.0213 Aa | 0.6497 ± 0.0211 Aa |
| 4 | 0.4651 ± 0.0159 Bb | 0.6264 ± 0.0226 Aa | 0.6358 ± 0.0355 Aa | 0.6117 ± 0.0154 Aab | |
| 8 | 0.4341 ± 0.0192 Bb | 0.5166 ± 0.0113 Ab | 0.5308 ± 0.0120 Ab | 0.5518 ± 0.0454 Ab | |
| Val * | 0 | 1.0950 ± 0.0422 Ba | 1.1995 ± 0.0470 Aa | 1.2016 ± 0.0360 Aa | 1.1656 ± 0.0133 Aa |
| 4 | 0.9270 ± 0.0352 Cb | 1.1733 ± 0.0532 ABa | 1.1971 ± 0.0282 Aa | 1.1213 ± 0.0179 Bb | |
| 8 | 0.9097 ± 0.0328 Bb | 0.9313 ± 0.0224 ABb | 0.9762 ± 0.0337 Ab | 0.9626 ± 0.0174 ABc | |
| Ala | 0 | 1.1794 ± 0.0104 Ca | 1.2946 ± 0.0580 Aa | 1.2611 ± 0.0297 ABa | 1.2058 ± 0.0430 BCa |
| 4 | 0.9946 ± 0.0517 Bb | 1.1938 ± 0.0146 Ab | 1.2281 ± 0.0185 Aa | 1.1952 ± 0.0423 Aa | |
| 8 | 0.9667 ± 0.0387 Db | 1.1441 ± 0.0331 Bb | 1.2220 ± 0.0295 Aa | 1.0394 ± 0.0386 Cb | |
| Asp | 0 | 1.7929 ± 0.0667 Ca | 2.1696 ± 0.0199 Aa | 2.0194 ± 0.0192 Ba | 1.9809 ± 0.0777 Ba |
| 4 | 1.7277 ± 0.0778 Ca | 1.9231 ± 0.0265 Bb | 2.0379 ± 0.0392 Aa | 1.7277 ± 0.0778 ABa | |
| 8 | 1.6787 ± 0.0651 Ba | 1.7047 ± 0.0550 Bc | 1.8970 ± 0.0903 Ab | 1.8695 ± 0.0287 Ab | |
| Cys | 0 | 0.0767 ± 0.0022 Ba | 0.0862 ± 0.0020 Aa | 0.0779 ± 0.0022 Ba | 0.0789 ± 0.0011 Ba |
| 4 | 0.0627 ± 0.0016 Bb | 0.0685 ± 0.0033 Ab | 0.0698 ± 0.0019 Ab | 0.0686 ± 0.0034 Aa | |
| 8 | 0.0599 ± 0.0007 Bb | 0.0604 ± 0.0013 Bc | 0.0678 ± 0.0009 Ab | 0.0645 ± 0.0032 Ab | |
| Glu | 0 | 1.1907 ± 0.0664 Ca | 1.5667 ± 0.0489 Aa | 1.3721 ± 0.0291 Ba | 1.5129 ± 0.0491 Aa |
| 4 | 1.0967 ± 0.0541 Ca | 1.2873 ± 0.0183 ABb | 1.2181 ± 0.0386 Bb | 1.3052 ± 0.0307 Ab | |
| 8 | 0.8386 ± 0.0172 Cb | 1.0834 ± 0.0206 Bc | 1.1535 ± 0.0518 Ab | 1.1410 ± 0.0144 Ac | |
| Gly | 0 | 0.6498 ± 0.0320 Ba | 0.6817 ± 0.0261 Aa | 0.6901 ± 0.0116 Ba | 0.6827 ± 0.0526 Ba |
| 4 | 0.6295 ± 0.0187 Ba | 0.6077 ± 0.0073 Aab | 0.6273 ± 0.0054 Ab | 0.6285 ± 0.0063 Ab | |
| 8 | 0.6194 ± 0.0073 Ba | 0.6055 ± 0.0223 Bb | 0.6226 ± 0.0053 Ab | 0.6132 ± 0.0155 Ac | |
| Pro | 0 | 0.5752 ± 0.0135 Ca | 0.7121 ± 0.0145 Aa | 0.7033 ± 0.0224 ABa | 0.6749 ± 0.0167 Ba |
| 4 | 0.4914 ± 0.0109 Bb | 0.6553 ± 0.0277 Ab | 0.6811 ± 0.0228 Ab | 0.6588 ± 0.0210 Aa | |
| 8 | 0.4792 ± 0.0319 Bb | 0.5247 ± 0.0354 Bc | 0.6177 ± 0.0191 Ab | 0.5899 ± 0.0308 Ab | |
| Ser | 0 | 0.2796 ± 0.0073 Ca | 0.4831 ± 0.0169 Aa | 0.4238 ± 0.0209 Ba | 0.4588 ± 0.0131 Aa |
| 4 | 0.2470 ± 0.0082 Cb | 0.3815 ± 0.0275 Bb | 0.4187 ± 0.0156 Aa | 0.4263 ± 0.0057 Ab | |
| 8 | 0.2343 ± 0.0090 Cb | 0.3596 ± 0.0059 Bb | 0.3931 ± 0.0105 Aa | 0.3739 ± 0.0194 ABc | |
| Tyr | 0 | 0.3742 ± 0.0046 Ba | 0.4413 ± 0.0102 Aa | 0.3909 ± 0.0087 Ba | 0.4254 ± 0.0116 Aa |
| 4 | 0.3399 ± 0.0125 Bb | 0.3769 ± 0.0089 Ab | 0.3742 ± 0.0139 Aab | 0.3798 ± 0.0179 Ab | |
| 8 | 0.3004 ± 0.0127 Cc | 0.3238 ± 0.0094 Bc | 0.3654 ± 0.0091 Ab | 0.3488 ± 0.0033 Ac | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Bao, J.; Si, Q.; Liu, M.; Bai, B.; Fu, Z.; Ge, G.; Jia, Y.; Wang, Z. Effects of Lactic Acid Bacteria Additives on Fatty Acids, Amino Acids and Antioxidant Capacity of Leymus chinensis Silage during Aerobic Exposure. Fermentation 2023, 9, 323. https://doi.org/10.3390/fermentation9040323
Liu Y, Bao J, Si Q, Liu M, Bai B, Fu Z, Ge G, Jia Y, Wang Z. Effects of Lactic Acid Bacteria Additives on Fatty Acids, Amino Acids and Antioxidant Capacity of Leymus chinensis Silage during Aerobic Exposure. Fermentation. 2023; 9(4):323. https://doi.org/10.3390/fermentation9040323
Chicago/Turabian StyleLiu, Yichao, Jian Bao, Qiang Si, Mingjian Liu, Baochao Bai, Zhihui Fu, Gentu Ge, Yushan Jia, and Zhijun Wang. 2023. "Effects of Lactic Acid Bacteria Additives on Fatty Acids, Amino Acids and Antioxidant Capacity of Leymus chinensis Silage during Aerobic Exposure" Fermentation 9, no. 4: 323. https://doi.org/10.3390/fermentation9040323