

Optimization of Coconut Milk Kefir Beverage by RSM and Screening of Its Metabolites and Peptides
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Coconut Milk Kefir Production
2.2. Experimental Design
2.3. pH
2.4. Antibacterial Activity
2.5. Antioxidant Assays
2.5.1. Diphenyl-2-picryl-hydrazyl Assay
2.5.2. 2,2′-Azino-bis (3-Ethylbenzothiazoline-6-sulfonic Acid) Diammonium Salt Assay
2.5.3. Ferric Reducing Antioxidant Power Assay
2.6. 1H-NMR Metabolomics Analysis
2.7. Identification of Peptides
2.8. Statistical Analysis
3. Results and Discussion
3.1. Coconut Milk Kefir Optimization
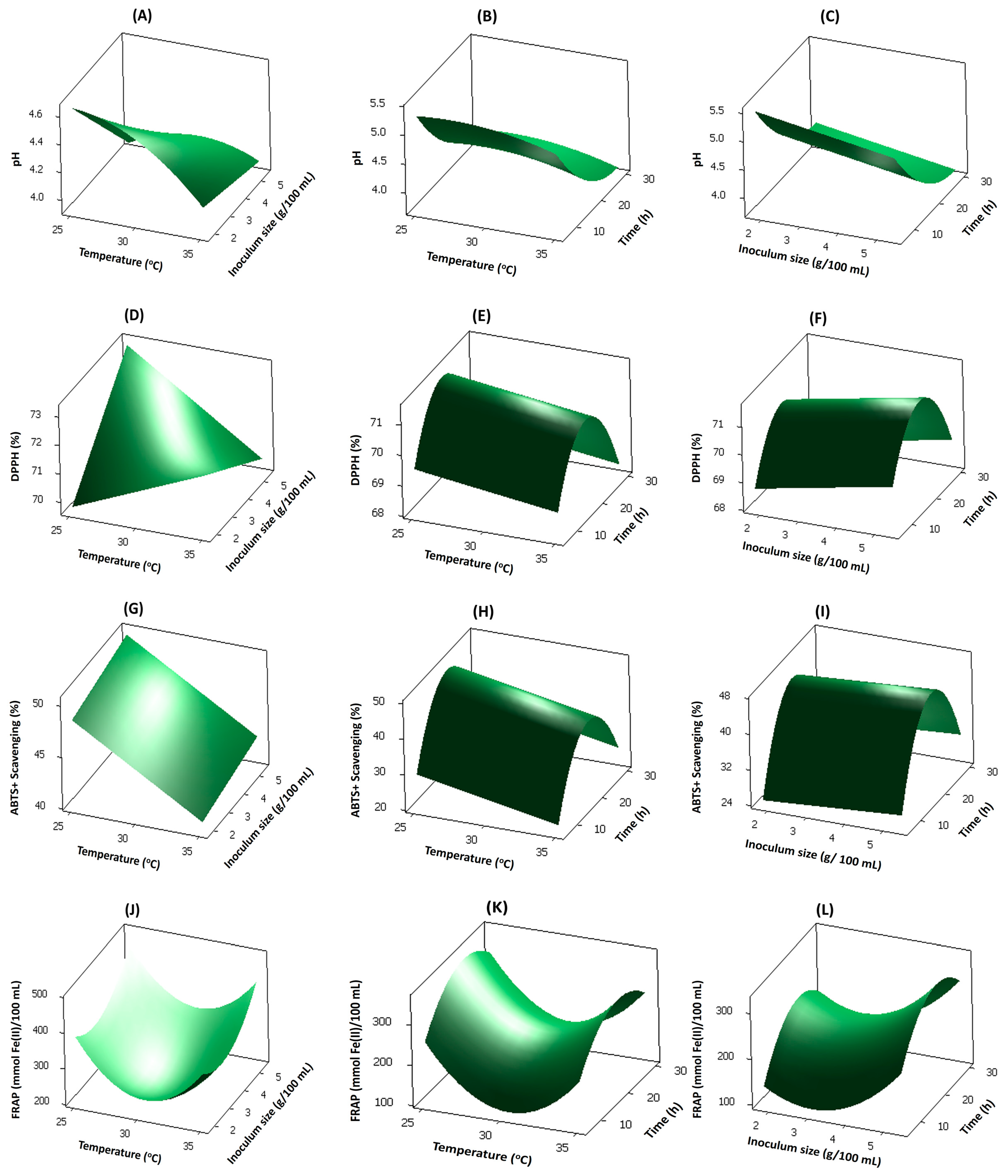
3.2. Effect of Process Variables on Response Variables
3.2.1. Effect of Process Variables on pH
3.2.2. Effect of Process Variables on Antioxidants
3.2.3. Effect of Process Variables on Antimicrobial Properties
3.3. Validation and Verification of the Predictive Model
3.4. Peptide Sequencing

{kind=link}
{kind=link}
{kind=link}
| Peptides | MW (Da) | Net Charge | Activity | Database Similarity | Reference |
|---|---|---|---|---|---|
| LTRGDEVAIFTPR | 1475 | 1 | Anti-Gram+ and Gram− | GSEIQPR (38.46%) | [32] |
| DEVAIFTPR | 1047 | 0 | Anti-Gram+ and Gram− | GSEIQPR (40%) | [32] |
| GDEVAIFTPR | 1104 | −1 | Anti-Gram+ and Gram− | RLGDGCTR (33.33%) | [33] |
| RVKLTRGDEVAIFTPR | 1858 | 3 | Anticancer | RLGDGCTR (37.5%) | [33] |
| LNALEPTR | 913 | 1 | Anticancer | RLGDGCTR (33.33%) | [33] |
| Peptides | MW (Da) | Net Charge | Activity | Database Similarity | Reference |
|---|---|---|---|---|---|
| LTRGDEVAIFAPR | 1445 | 1 | Anti-Gram+ and Gram− | GSEIQPR (38.46%) | [32] |
| GDEVAIFAPR | 1074 | 0 | Anti-Gram+ and Gram− | RLGDGCTR (33.33%) | [32] |
| RVKLTRGDEVAIFAPR | 1828 | 3 | Anticancer | RLGDGCTR(37.5%) | [32] |
| IERLNALEPTR | 1311 | 1 | Anti-Gram− | IVRVAVALRRIR (41.66%) | [34] |
| AMVSSIVGK | 891 | 1 | Anti-Gram+ and Gram−, Anti-MRSA, anti-sepsis | KVTKSVKSIPVKI (40%) | [35] |
3.5. Bioactive Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abadl, M.M.; Mohsin, A.Z.; Sulaiman, R.; Abas, F.; Muhialdin, B.J.; Hussin, A.S. Biological activities and physiochemical properties of low-fat and high-fat coconut-based kefir. Int. J. Gastron. Food Sci. 2022, 30, 100624. [Google Scholar] [CrossRef]
- de Oliveira Leite, A.M.; Miguel, M.A.L.; Peixoto, R.S.; Rosado, A.S.; Silva, J.T.; Paschoalin, V.M.F. Microbiological, technological and therapeutic properties of kefir: A natural probiotic beverage. Braz. J. Microbiol. 2013, 44, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.D.; Dias, M.M.S.; Grześkowiak, Ł.M.; Reis, S.A.; Conceição, L.L.; Maria do Carmo, G.P. Milk kefir: Nutritional, microbiological and health benefits. Nutr. Res. Rev. 2017, 30, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Wang, Y.; Ahmad, A.; Khan, S.T.; Nisa, M.; Ahmad, H.; Afreen, A. Kefir and Health: A Contemporary Perspective. Crit. Rev. Food Sci. Nutr. 2013, 53, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Tóth, A.G.; Csabai, I.; Maróti, G.; Jerzsele, Á.; Dubecz, A.; Patai, Á.V.; Judge, M.F.; Nagy, S.Á.; Makrai, L.; Bányai, K.; et al. A glimpse of antimicrobial resistance gene diversity in kefir and yoghurt. Sci. Rep. 2020, 10, 22458. [Google Scholar] [CrossRef]
- Rethinam, P. International scenario of coconut sector. In The Coconut Palm (Cocos nucifera L.)-Research and Development Perspectives; Springer: Singapore, 2019; pp. 21–56. [Google Scholar]
- Lappano, R.; Sebastiani, A.; Cirillo, F.; Rigiracciolo, D.C.; Galli, G.R.; Curcio, R.; Malaguarnera, R.; Belfiore, A.; Cappello, A.R.; Maggiolini, M. The lauric acid-activated signaling prompts apoptosis in cancer cells. Cell Death Discov. 2017, 3, 17063. [Google Scholar] [CrossRef]
- DebMandal, M.; Mandal, S. Coconut (Cocos nucifera L.: Arecaceae): In health promotion and disease prevention. Asian Pac. J. Trop. Med. 2011, 3, 241–247. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. New perspectives in food allergy. Int. J. Mol. Sci. 2020, 21, 1474. [Google Scholar] [CrossRef]
- Iguacel, I.; Huybrechts, I.; Moreno, L.A.; Michels, N. Vegetarianism and veganism compared with mental health and cognitive outcomes: A systematic review and meta-analysis. Nutr. Rev. 2021, 79, 361–381. [Google Scholar] [CrossRef]
- dos Santos, D.C.; de Oliveira Filho, J.G.; Santana, A.C.A.; de Freitas, B.S.M.; Silva, F.G.; Takeuchi, K.P.; Egea, M.B. Optimization of soymilk fermentation with kefir and the addition of inulin: Physicochemical, sensory and technological characteristics. LWT 2019, 104, 30–37. [Google Scholar] [CrossRef]
- Straits Research. Coconut Milk Market: Information by Packaging Type (Plastic Bottles, Tetra Packs), Category (Organic, Conventional), End-Users (Household, Food Services), and Region—Forecast till 2030. 2022. Available online: https://straitsresearch.com/report/coconut-milk-market (accessed on 14 March 2023).
- The Brainy Insights. Global Kefir Market, Industry Analysis, Trends, Analysis and Forecast, 2022 to 2030. 2022. Available online: https://www.thebrainyinsights.com/report/kefir-market-12895#summary (accessed on 14 March 2023).
- Rodrigues, K.L.; Caputo, L.R.G.; Carvalho, J.C.T.; Evangelista, J.; Schneedorf, J.M. Antimicrobial and healing activity of kefir and kefiran extract. Int. J. Antimicrob. Agents 2005, 25, 404. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Kadum, H.; Hamid, A.A.; Abas, F.; Ramli, N.S.; Mohammed, A.K.S.; Muhialdin, B.J.; Jaafar, A.H. Bioactive Compounds Responsible for Antioxidant Activity of Different Varieties of Date (Phoenix dactylifera L.) Elucidated by 1H-NMR Based Metabolomics. Int. J. Food Prop. 2019, 22, 462–476. [Google Scholar] [CrossRef]
- Hussein, F.A.; Chay, S.Y.; Zarei, M.; Auwal, S.M.; Hamid, A.A.; Wan Ibadullah, W.Z.; Saari, N. Whey protein concentrate as a novel source of bifunctional peptides with angiotensin-I converting enzyme inhibitory and antioxidant properties: RSM study. Foods 2020, 9, 64. [Google Scholar] [CrossRef]
- Koh, W.Y.; Utra, U.; Rosma, A.; Effarizah, M.E.; Rosli, W.I.; Park, Y.H. Development of a novel fermented pumpkin-based beverage inoculated with water kefir grains: A response surface methodology approach. Food Sci. Biotechnol. 2018, 27, 525–535. [Google Scholar] [CrossRef]
- Harrell, F.E., Jr.; Harrell, F.E. Multivariable modeling strategies. In Regression Modeling Strategies: With Applications to Linear Models, Logistic and Ordinal Regression, and Survival Analysis; Springer: Cham, Switzerland, 2015; pp. 63–102. [Google Scholar]
- Marzlan, A.A.; Muhialdin, B.J.; Abedin, N.H.Z.; Mohammed, N.K.; Abadl, M.M.T.; Roby, B.H.M.; Hussin, A.S.M. Optimized supercritical CO2 extraction conditions on yield and quality of torch ginger (Etlingera elatior (Jack) RM Smith) inflorescence essential oil. Ind. Crops Prod. 2020, 154, 112581. [Google Scholar] [CrossRef]
- Pandey, S.M.; Mishra, H.N. Optimization of the prebiotic & probiotic concentration and incubation temperature for the preparation of synbiotic soy yoghurt using response surface methodology. LWT-Food Sci. Technol. 2015, 62, 458–467. [Google Scholar]
- Wu, S.; Li, D.; Li, S.-J.; Bhandari, B.; Yang, B.-l.; Chen, X.D.; Mao, Z.-H. Effects of incubation temperature, starter culture level and total solids content on the rheological properties of yogurt. Int. J. Food Eng. 2009, 5, 1–17. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Bąbelewski, P. Phenolic and carotenoid profile of new goji cultivars and their anti-hyperglycemic, anti-aging and antioxidant properties. J. Funct. Foods 2018, 48, 632–642. [Google Scholar] [CrossRef]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of Milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, T.; Sahin, S.; Akpinar-Bayizit, A.; Yilmaz-Ersan, L. Assessment of antioxidant capacity by method comparison and amino acid characterisation in buffalo milk kefir. Int. J. Dairy Technol. 2019, 72, 65–73. [Google Scholar] [CrossRef]
- Guzel-Seydim, Z.B.; Kok-Tas, T.; Greene, A.K.; Seydim, A.C. Review: Functional Properties of Kefir. Crit. Rev. Food Sci. Nutr. 2011, 51, 261–268. [Google Scholar] [CrossRef] [PubMed]
- John, S.M.; Deeseenthum, S. Properties and benefits of kefir-A review. Songklanakarin J. Sci. Technol. 2015, 37, 275–282. [Google Scholar]
- Santos, A.; San Mauro, M.; Sanchez, A.; Torres, J.; Marquina, D. The antimicrobial properties of different strains of Lactobacillus spp. isolated from kefir. Syst. Appl. Microbiol. 2003, 26, 434–437. [Google Scholar] [CrossRef]
- Kim, D.-H.; Jeong, D.; Kim, H.; Kang, I.-B.; Chon, J.-W.; Song, K.-Y.; Seo, K.-H. Antimicrobial Activity of Kefir against Various Food Pathogens and Spoilage Bacteria. Korean J. Food Sci. Anim. Resour. 2016, 36, 787–790. [Google Scholar] [CrossRef]
- Stoffels, G.; Nissen-Meyer, J.; Gudmundsdottir, A.; Sletten, K.; Holo, H.; Nes, I. Purification and characterization of a new bacteriocin isolated from a Carnobacterium sp. Appl. Environ. Microbiol. 1992, 58, 1417–1422. [Google Scholar] [CrossRef]
- Mishra, A.; Gauri, S.S.; Mukhopadhyay, S.K.; Chatterjee, S.; Das, S.S.; Mandal, S.M.; Dey, S. Identification and structural characterization of a new pro-apoptotic cyclic octapeptide cyclosaplin from somatic seedlings of Santalum album L. Peptides 2014, 54, 148–158. [Google Scholar] [CrossRef]
- Hilpert, K.; Elliott, M.; Jenssen, H.; Kindrachuk, J.; Fjell, C.D.; Körner, J.; Ulrich, A.S. Screening and characterization of surface-tethered cationic peptides for antimicrobial activity. Chem. Biol. 2009, 16, 58–69. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, E.; Yuan, C.; Zhang, L.; Yousef, A.E. Isolation of a Paenibacillus sp. strain and structural elucidation of its broad-spectrum lipopeptide antibiotic. Appl. Environ. Microbiol. 2012, 78, 3156–3165. [Google Scholar] [CrossRef] [PubMed]
- Arulrajah, B.; Muhialdin, B.J.; Zarei, M.; Hasan, H.; Saari, N. Lacto-fermented Kenaf (Hibiscus cannabinus L.) seed protein as a source of bioactive peptides and their applications as natural preservatives. Food Control 2020, 110, 106969. [Google Scholar] [CrossRef]
- Hancock, R.E.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Muhialdin, B.J.; Hassan, Z.; Abu Bakar, F.; Algboory, H.L.; Saari, N. Novel antifungal peptides produced by Leuconostoc mesen-teroides DU15 effectively inhibit growth of Aspergillus niger. J. Food Sci. 2015, 80, M1026–M1030. [Google Scholar] [CrossRef] [PubMed]
- Guilhelmelli, F.; Vilela, N.; Albuquerque, P.; Derengowski, L.; Silva-Pereira, I.; Kyaw, C. Antibiotic development challenges: The various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front. Microbiol. 2013, 4, 353. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Feng, L.; Zhao, S.; Chen, G.; Jiang, W.; Liu, Y.; Jiang, J.; Zhou, X. Antioxidant status of serum, muscle, intestine and hepatopancreas for fish fed graded levels of biotin. Fish Physiol. Biochem. 2014, 40, 499–510. [Google Scholar] [CrossRef]
- Qudah, K.M.; Ismail, Z.B. The relationship between serum biotin and oxidant/antioxidant activities in bovine lameness. Res. Vet. Sci. 2012, 92, 138–141. [Google Scholar] [CrossRef]
- Ma, N.; Abaker, J.A.; Bilal, M.S.; Dai, H.; Shen, X. Sodium butyrate improves antioxidant stability in sub-acute ruminal acidosis in dairy goats. BMC Vet. Res. 2018, 14, 275. [Google Scholar] [CrossRef]
- Manyam, B.V.; Katz, L.; Hare, T.A.; Kaniefski, K.; Tremblay, R.D. Isoniazid-induced elevation of CSF GABA levels and effects on chorea in Huntington’s disease. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1981, 10, 35–37. [Google Scholar] [CrossRef]
- Bown, A.W.; Shelp, B.J. The Metabolism and Functions of [gamma]-Aminobutyric Acid. Plant Physiol. 1997, 115, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [PubMed]
- Muhialdin, B.J.; Kadum, H.; Zarei, M.; Hussin, A.S. Effects of metabolite changes during lacto-fermentation on the biological activity and consumer acceptability for dragon fruit juice. LWT 2020, 121, 108992. [Google Scholar] [CrossRef]
- Del Valle, M.J.; Laiño, J.E.; de LeBlanc, A.D.M.; de Giori, G.S.; LeBlanc, J.G. (Soyamilk fermented with riboflavin-producing Lactobacillus plantarum CRL 2130 reverts and prevents ariboflavinosis in murine models. Br. J. Nutr. 2016, 116, 1229–1235. [Google Scholar] [CrossRef]
- Zhu, Y.-Y.; Thakur, K.; Feng, J.-Y.; Cai, J.-S.; Zhang, J.-G.; Hu, F.; Wei, Z.-J. Riboflavin-overproducing lactobacilli for the enrich-ment of fermented soymilk: Insights into improved nutritional and functional attributes. Appl. Microbiol. Biotechnol. 2020, 104, 5759–5772. [Google Scholar] [CrossRef] [PubMed]


| Factors | Code | Variable Levels | ||
|---|---|---|---|---|
| −1 | 0 | +1 | ||
| Temperature (°C) | x1 | 25 | 30 | 35 |
| Inoculum size (g/100 mL) | x2 | 1.8 | 3.6 | 5.4 |
| incubation time (h) | x3 | 6 | 18 | 30 |
| Run | x1 | x2 | x3 | Responses | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | A (°C) | C | A (g/100 mL) | C | A (h) | y1 | y2 (%) | y3 (%) | y4 (mmol Fe(II)/10 mL) | y5 (%) | y6 (%) | y7 (%) | y8 (%) | |
| 1 | −1 | 25 | −1 | 1.8 | 0 | 18 | 4.65 | 70.232 | 45.611 | 384.648 | 85.90 | 87.78 | 95.65 | 84.61 |
| 2 | +1 | 35 | −1 | 1.8 | 0 | 18 | 4.04 | 71.370 | 40.214 | 296.685 | 99.64 | 99.28 | 100.00 | 96.22 |
| 3 | −1 | 25 | +1 | 5.4 | 0 | 18 | 4.00 | 72.652 | 60.113 | 445.389 | 100.00 | 99.21 | 99.65 | 100.00 |
| 4 | +1 | 35 | +1 | 5.4 | 0 | 18 | 3.98 | 68.663 | 32.806 | 491.685 | 94.86 | 99.97 | 99.97 | 95.61 |
| 5 | −1 | 25 | 0 | 3.6 | −1 | 6 | 5.22 | 68.014 | 24.435 | 217.796 | 62.43 | 67.54 | 68.15 | 65.04 |
| 6 | +1 | 35 | 0 | 3.6 | −1 | 6 | 5.12 | 69.254 | 27.153 | 211.130 | 83.41 | 78.70 | 71.76 | 82.75 |
| 7 | −1 | 25 | 0 | 3.6 | +1 | 30 | 3.99 | 68.877 | 25.481 | 365.944 | 91.29 | 98.62 | 99.73 | 89.22 |
| 8 | +1 | 35 | 0 | 3.6 | +1 | 30 | 3.70 | 67.989 | 25.241 | 257.796 | 82.43 | 85.02 | 92.91 | 79.31 |
| 9 | 0 | 30 | −1 | 1.8 | 0 | 6 | 5.60 | 67.943 | 24.559 | 144.093 | 76.40 | 70.36 | 69.22 | 74.68 |
| 10 | 0 | 30 | +1 | 5.4 | 0 | 6 | 4.99 | 71.421 | 26.574 | 245.389 | 83.06 | 89.74 | 99.88 | 85.67 |
| 11 | 0 | 30 | −1 | 1.8 | +1 | 30 | 4.15 | 68.503 | 34.111 | 237.611 | 88.42 | 100.00 | 99.82 | 86.59 |
| 12 | 0 | 30 | +1 | 5.4 | +1 | 30 | 3.81 | 68.972 | 32.816 | 201.130 | 81.77 | 98.58 | 98.08 | 80.68 |
| 13 | 0 | 30 | 0 | 3.6 | 0 | 18 | 4.33 | 71.332 | 47.574 | 219.352 | 98.51 | 98.57 | 99.80 | 99.65 |
| 14 | 0 | 30 | 0 | 3.6 | 0 | 18 | 4.28 | 72.354 | 45.639 | 200.574 | 98.93 | 99.99 | 99.97 | 98.39 |
| 15 | 0 | 30 | 0 | 3.6 | 0 | 18 | 4.25 | 71.685 | 47.287 | 227.611 | 97.84 | 99.32 | 100.00 | 98.63 |
| y1 | y2 (%) | y3 (%) | y4 (mmol Fe(II)/10 mL) | y5 (%) | y6 (%) | y7 (%) | y8 (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RC | PV | RC | PV | RC | PV | RC | PV | RC | PV | RC | PV | RC | PV | RC | PV | |
| Intercept | ||||||||||||||||
| Constant | 4.552 | 51.989 | 22.889 | 4711.20 | −86.944 | −211.86 | 23.730 | −215.96 | ||||||||
| Linear | ||||||||||||||||
| x1 | 0.213 | 0.053 | 0.450 | 0.082 | −0.756 | 0.105 | −297.75 | 0.001 | 4.644 | 0.002 | 14.438 | 0.011 | 0.037 | 0.935 | 12.992 | 0.008 |
| x2 | −0.608 | 0.001 | 4.527 | 0.035 | 0.543 | 0.655 | −125.12 | 0.066 | 16.377 | 0.039 | 6.421 | 0.004 | 9.034 | 0.011 | 18.232 | 0.006 |
| x3 | −0.141 | 0.000 | 0.554 | 0.004 | 4.671 | 0.000 | 21.32 | 0.013 | 7.976 | 0.000 | 7.760 | 0.000 | 5.161 | 0.000 | 8.428 | 0.000 |
| Quadratic | ||||||||||||||||
| x1x1 | −0.005 | 0.012 | 4.90 | 0.002 | −0.206 | 0.022 | −0.149 | 0.032 | ||||||||
| x2x2 | 20.47 | 0.035 | ||||||||||||||
| x3x3 | 0.0024 | 0.000 | −0.016 | 0.004 | −0.125 | 0.000 | −0.52 | 0.020 | −0.107 | 0.000 | −0.084 | 0.000 | −0.0823 | 0.005 | −0.111 | 0.000 |
| Interaction | ||||||||||||||||
| x1x2 | 0.0164 | 0.004 | −0.1424 | 0.042 | −0.524 | 0.045 | −0.444 | 0.021 | ||||||||
| x1x3 | −0.124 | 0.006 | −0.103 | 0.008 | −0.115 | 0.002 | ||||||||||
| x2x3 | −0.241 | 0.018 | −0.375 | 0.027 | −0.196 | 0.017 | ||||||||||
| R2 | 99.04% | 73.70% | 79.13% | 85.67% | 91.99% | 95.55% | 83.80% | 97.34% | ||||||||
| R2-adjust | 98.33% | 59.09% | 70.79% | 74.92% | 85.99% | 91.10% | 74.80% | 93.80% | ||||||||
| p-value | 0.000 | 0.018 | 0.002 | 0.005 | 0.001 | 0.000 | 0.002 | 0.000 | ||||||||
| F value | 138.22 | 5.04 | 9.48 | 7.97 | 15.32 | 21.47 | 9.31 | 27.47 | ||||||||
| Lack of fit (p-value) | 0.223 | 0.167 | 0.024 | 0.056 | 0.014 | 0.031 | 0.000 | 0.044 | ||||||||
| Lack of fit (F-value) | 3.80 | 5.32 | 41.08 | 17.10 | 69.01 | 31.30 | 4161.17 | 21.81 | ||||||||
| Response Variables | Goal | Maximum/Minimum Values | |
|---|---|---|---|
| Predicted | Experimental (n = 3) | ||
| pH | In range | 4.2 | 4.17 ± 0.21 |
| DPPH scavenging activity (%) | Maximize | 72.6 | 71.17 ± 3.67 |
| ABTS+ scavenging activity (%) | Maximize | 47.06 | 43.63 ± 3.42 |
| FRAP (mmol Fe(II)/10 mL) | Maximize | 437.045 | 419.6 ± 17.7 |
| Antimicrobial activity against B. subtilis (%) | Maximize | 93.1 | 90.4 ± 1.35 |
| Antimicrobial activity against Salmonella Typhimurium (%) | Maximize | 97.3 | 97.2 ± 1.54 |
| Antimicrobial activity against E. coli (%) | Maximize | 100 | 99.6 ± 0.64 |
| Antimicrobial activity against S. aureus (%) | Maximize | 94.8 | 94.7 ± 1.65 |
| Metabolites | 1H-NMR Characteristic Signals | CMK | CM |
|---|---|---|---|
| Butyrate | δ 0.88 (t) | 0.0557 | ND |
| Threonine | δ 1.316 (d) | ND | 0.2543 |
| Lactic acid | δ 1.32 (d), δ 4.14 (q) | 1.0271 | ND |
| Alanine | δ 1.46 (d) | 0.0442 | 0.1167 |
| Biotin | δ 1.61 (dd) | 0.0816 | ND |
| Caprylic acid | δ 1.61 (br. S.) | 0.6683 | ND |
| 4-Aminobutyrate (GABA) | δ 1.89 (m), δ 2.28 (t), δ 3.00 (t) | 0.1642 | 0.0657 |
| gamma-Butyrolactone | δ 2.24 (m) | 0.3383 | ND |
| 3-Hydroxybutyrate | δ 2.314 (m), δ 2.414 (m) | ND | 0.3137 |
| Malic acid | δ 2.36 (dd), δ 2.66 (dd) | ND | 0.2602 |
| Riboflavin | δ 2.5 (t) | 0.1440 | ND |
| Lysine | δ 3.02 (t) | 0.0708 | 0.1281 |
| Choline | δ 3.189 (s), δ 3.507 (dd), δ 4.056 (m) | 0.0213 | 0.0157 |
| Carnitine | δ 3.22 (s) | 0.0101 | ND |
| Glucuronate | δ 3.289 (t) | 2.5208 | 5.2900 |
| Xylose | δ 3.42 (t), δ 3.51 (dd) | 2.3029 | 4.7923 |
| Sucrose | δ 3.67 (s), δ 3.87 (dd), δ 4.04 (t), δ 4.21 (d), 5.4 (d) | 2.0539 | 2.9443 |
| 2-Aminobutyrate | δ 3.718 (dd) | 1.8177 | ND |
| Leucine | δ 3.722 (m) | ND | 1.8325 |
| Guanidoacetate | δ 3.78 (s) | 0.2967 | 0.9874 |
| Ethanolamine | δ 3.81 (d) | 0.262 | 1.5021 |
| Gluconate | 4.15 (d) | 0.3070 | 1.5317 |
| O-Acetylcarnitine | δ 5.57 (q) | ND | 0.0176 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abadl, M.M.T.; Marzlan, A.A.; Sulaiman, R.; Abas, F.; Meor Hussin, A.S. Optimization of Coconut Milk Kefir Beverage by RSM and Screening of Its Metabolites and Peptides. Fermentation 2023, 9, 430. https://doi.org/10.3390/fermentation9050430
Abadl MMT, Marzlan AA, Sulaiman R, Abas F, Meor Hussin AS. Optimization of Coconut Milk Kefir Beverage by RSM and Screening of Its Metabolites and Peptides. Fermentation. 2023; 9(5):430. https://doi.org/10.3390/fermentation9050430
Chicago/Turabian StyleAbadl, Muna Mahmood Taleb, Anis Asyila Marzlan, Rabiha Sulaiman, Faridah Abas, and Anis Shobirin Meor Hussin. 2023. "Optimization of Coconut Milk Kefir Beverage by RSM and Screening of Its Metabolites and Peptides" Fermentation 9, no. 5: 430. https://doi.org/10.3390/fermentation9050430
APA StyleAbadl, M. M. T., Marzlan, A. A., Sulaiman, R., Abas, F., & Meor Hussin, A. S. (2023). Optimization of Coconut Milk Kefir Beverage by RSM and Screening of Its Metabolites and Peptides. Fermentation, 9(5), 430. https://doi.org/10.3390/fermentation9050430