

Evaluation of the Amino Acid Composition and Content of Organic Acids of Complex Postbiotic Substances Obtained on the Basis of Metabolites of Probiotic Bacteria Lacticaseibacillus paracasei ABK and Lactobacillus helveticus H9
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Postbiotic Substances
2.2. Determination of Amino Acid Composition, Free Amino and Organic Acids Content
2.3. Genome Analysis
2.4. Statistical Data Analysis
3. Results and Discussion
3.1. Composition of Total Amino Acids
3.2. Composition of Free Amino Acids

3.3. Content of Organic Acids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kudryasheva, A.A. The impact of nutrition on human health. Food Ind. 2004, 12, 88–90. [Google Scholar]
- Shenderov, B.A. Metabiotics is a new technology for the prevention of diseases associated with human microecological imbalance. J. Restor. Med. 2017, 4, 40. [Google Scholar]
- Ardatskaya, M.D.; Stolyarova, L.G.; Arkhipova, E.V.; Filimonova, O.Y.U. Metabiotics as a natural development of the probiotic concept. Difficult Patient 2020, 15, 35–39. [Google Scholar]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Factories 2020, 19, 1–22. [Google Scholar] [CrossRef]
- Oleskin, A.V.; Shenderov, B.A. Microbial Communication and Microbiota-Host Interactivity: Neurophysiological, Biotechnological, and Biopolitical Implications; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2020; pp. 1–389. [Google Scholar]
- Aggeletopoulou, I.; Konstantakis, C.; Assimakopoulos, S.F.; Triantos, C. The role of the gut microbiota in the treatment of inflammatory bowel diseases. Microb. Pathog. 2019, 137, 103774. [Google Scholar] [CrossRef]
- Begunova, A.V. Biologically active metabolites of lactic acid bacteria. Food Ind. 2022, 6, 21–25. [Google Scholar]
- Newburg, D.S.; Ko, J.S.; Leone, S.; Nanthakumar, N.N. Human milk oligosaccharides and synthetic galactosyloligosaccharides contain 3′-, 4-, and 6′-galactosyllactose and attenuate inflammation in human T84, NCM-460, and H4 cells and intestinal tissue ex vivo. J. Nutr. 2016, 146, 358–367. [Google Scholar] [CrossRef]
- Pederson, J.A.; Steele, J.L.; Christensen, J.E.; Dudley, E.G. Peptidases and amino acid catabolism in lactic acid bacteria. In Lactic Acid Bacteria: Genetics, Metabolism and Applications, Proceedings of the Sixth Symposium on Lactic Acid Bacteria: Genetics, Metabolism and Applications, Veldhoven, The Netherlands, 19–23 September 1999; Springer: Veldhoven, The Netherlands, 1999. [Google Scholar]
- Belenguer, A.; Duncan, S.H.; Calder, a.G.; Holtrop, G.; Louis, P.; Lobley, G.E.; Flint, H.J. Two routes of metabolic cross-feeding between Bifidobacterium adolescentis and butyrate-producing anaerobes from the human gut. Appl. Environ. Microbiol. 2006, 72, 3593–3599. [Google Scholar] [CrossRef]
- Scheppach, W. Effects of short chain fatty acids on gut morphology and function. Gut 1994, 35, S35–S38. [Google Scholar] [CrossRef]
- Bush, R.S.; Milligan, L.P. Study of the mechanism of inhibition of ketogenesis by propionate in bovine liver. Can. J. Anim. Sci. 1971, 51, 121–127. [Google Scholar] [CrossRef]
- Péant, B.; LaPointe, G.; Gilbert, C.; Atlan, D.; Ward, P.; Roy, D. Comparative analysis of the exopolysaccharide biosynthesis gene clusters from four strains of Lactobacillus rhamnosus. Microbiology 2005, 151, 1839–1851. [Google Scholar] [CrossRef]
- Razumovskij, N.; Sobolev, D. Amino acids-essential and non-essential. Livest. Russ. 2020, 2, 59–62. [Google Scholar]
- Kudryasheva, A.A.; Presnyakova, O.P. Medico-biological features of natural food amino acids. Food Ind. 2014, 3, 68–73. [Google Scholar]
- World Health Organization; United Nations University. Protein and Amino Acid Requirements in Human Nutrition; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Sheybak, V.M.; Pavlyukovets, A.Y. Tryptofan: Key metabolite of homeostasis and regulator of body functions. Hepatol. Gastroenterol. 2021, 5, 143–149. [Google Scholar] [CrossRef]
- Agarkova, Y.; Fedotova, O.; Chilikin, A. The prospect of using natural psychobitics in dairy products to stabilize the diet. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 677, p. 032051. [Google Scholar]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut microbiota: The neglected endocrine organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef]
- Thorburn, A.N.; Laurence, M.; Charles, R.M. Diet, metabolites, and “western-lifestyle” inflammatory diseases. Immunity 2014, 40, 833–842. [Google Scholar] [CrossRef]
- Dankovtsev, R.Y.; Lidokhova, O.V.; Gubina, O.I. Some aspects of the physiological and pathological role of glycine. Sci. Educ. Dev. Trends 2020, 61, 31–34. [Google Scholar]
- Sokolova, O.V.; Fedotova, O.B. About the possibilities of enrichment of polycomponent ferrous dairy products with vitamins and amino acids in a native form. In Proceedings of the International Scientific and Practical Conference Dedicated to the Memory of Vasily Matveyevich Gorbatov; no. 1. Federal State Budgetary Scientific Institution Federal Scientific Center for Food Systems Named after V.I. VM Gorbatov RAS, Moscow, 2015; Available online: https://eng.rudn.ru/cooperation/employment-partnerships/partners/federal-scientific-center-for-food-systems-na-vm-gorbatova-russian-academy-of-sciences-federal-state-budgetary-scientific-institution/ (accessed on 28 March 2023).
- Lim, Y.H.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdullah, N. Comparative studies of versatile extracellular proteolytic activities of lactic acid bacteria and their potential for extracellular amino acid productions as feed supplements. J. Anim. Sci. Biotechnol. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Toe, C.J.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Rahim, R.A.; Idrus, Z. Extracellular proteolytic activity and amino acid production by lactic acid bacteria isolated from Malaysian foods. Int. J. Mol. Sci. 2019, 20, 1777. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef]
- Landi, N.; Ragucci, S.; Di Maro, A. Amino Acid Composition of Milk from Cow, Sheep and Goat Raised in Ailano and Valle Agricola, Two Localities of ‘Alto Casertano’ (Campania Region). Foods 2021, 10, 2431. [Google Scholar] [CrossRef] [PubMed]
- Vasson, M.-P.; Farges, M.-C.; Sarret, A.; Cynober, L. Free amino acid concentrations in milk: Effects of microwaveversus conventional heating. Amino. Acids 1998, 15, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, J.; Vehlow, C.; Battke, F.; Jäger, G.; Weiskopf, D.; Nieselt, K. iHAT: Interactive Hierarchical Aggregation Table for Genetic Association Data. BMC Bioinform. 2012, 13, S2. [Google Scholar] [CrossRef] [PubMed]
- Suissa, R.; Oved, R.; Jankelowitz, G.; Turjeman, S.; Koren, O.; Kolodkin-Gal, I. Molecular genetics for probiotic engineering: Dissecting lactic acid bacteria. Trends Microbiol. 2022, 30, 293–306. [Google Scholar] [CrossRef]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Iskandar, C.F.; Cailliez-Grimal, C.; Borges, F.; Revol-Junelles, A.-M. Review of lactose and galactose metabolism in Lactic Acid Bacteria dedicated to expert genomic annotation. Trends Food Sci. Technol. 2019, 88, 121–132. [Google Scholar] [CrossRef]
- De Angelis, M.; Bottacini, F.; Fosso, B.; Kelleher, P.; Calasso, M.; Di Cagno, R.; Ventura, M.; Picardi, E.; van Sinderen, D.; Gobbetti, M. Lactobacillus rossiae, a Vitamin B12 Producer, Represents a Metabolically Versatile Species within the Genus Lactobacillus. PLoS ONE 2014, 9, e107232. [Google Scholar] [CrossRef]
- Zalán, Z.; Hudáček, J.; Štětina, J.; Chumchalová, J.; Halász, A. Production of organic acids by Lactobacillus strains in three different media. Eur. Food Res. Technol. 2010, 230, 395–404. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
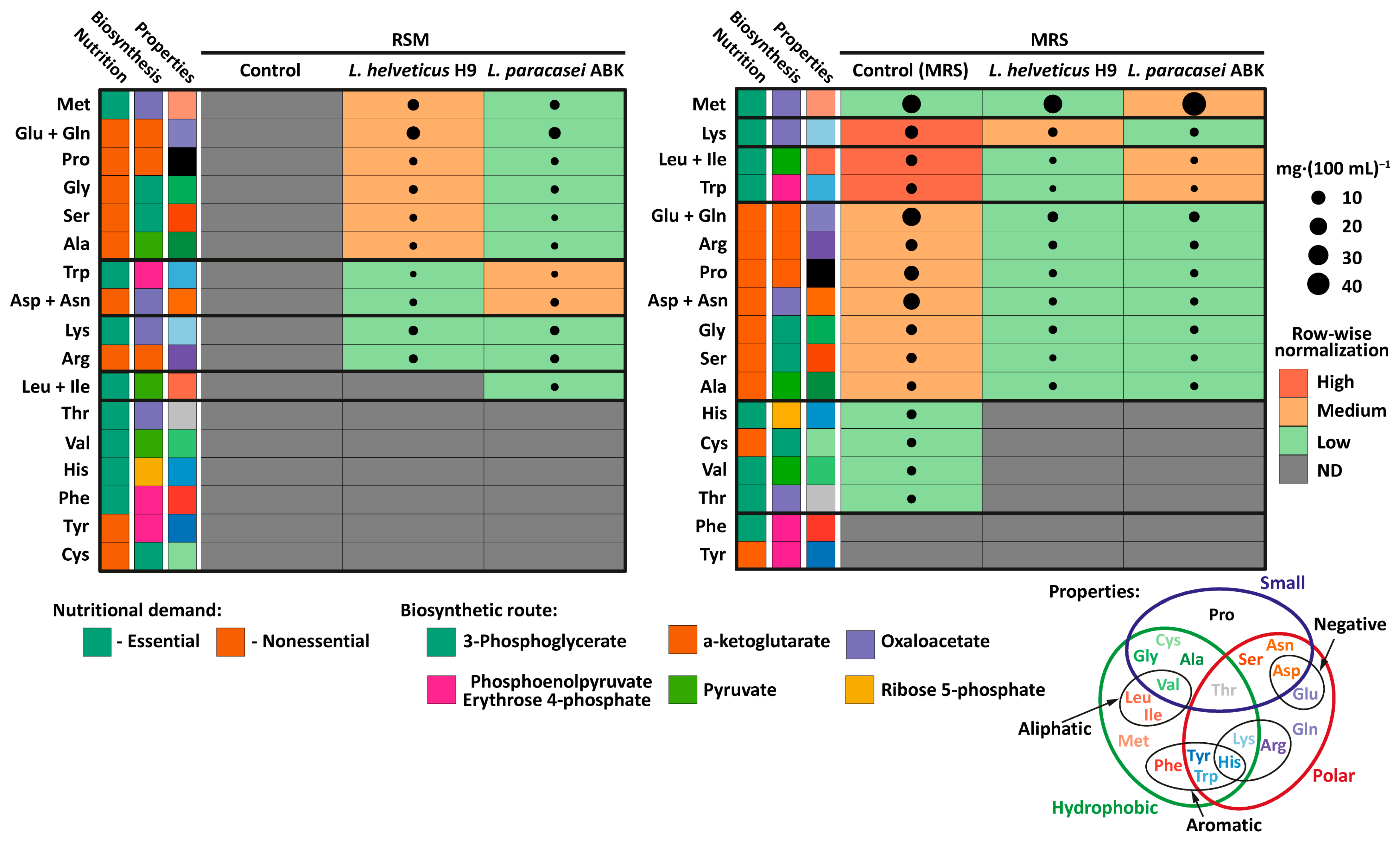
| Amino Acid | Total Amino Acids, mg∙(100 mL)−1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RSM | MRS | |||||||||||
| Control | L. helveticus H9 | L. paracasei ABK | Control | L. helveticus H9 | L. paracasei ABK | |||||||
| Essential | ||||||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Leu + Ile | 296 a | 44 | 20 b | 3 | 14 b | 2 | 177 a | 14 | 183 a | 15 | 184 a | 15 |
| Lys | 138 a | 20 | 8.0 b | 1.2 | 5.3 c | 0.8 | 97 a | 15 | 101 a | 15 | 109 a | 16 |
| Val | 123 a | 18 | 9.3 b | 1.4 | 6.1 c | 0.9 | 102 a | 15 | 100 a | 15 | 85 a | 15 |
| Phe | 106 a | 16 | 4.6 b | 0.7 | 2.8 c | 0.4 | 70 a | 3.1 | 70 a | 3 | 70 a | 3 |
| Thr | 92 a | 14 | 6.4 b | 0.9 | 5.2 b | 0.8 | 49 a | 7.0 | 47 a | 7 | 46 a | 7 |
| Met | 81 a | 12 | 8.6 b | 1.3 | 3.0 c | 0.4 | 39 a | 6.0 | 46 a | 7 | 111 b | 17 |
| His | 83 a | 7 | 12 b | 1 | 10 c | 1 | 56 a | 8.0 | 56 a | 8 | 55 a | 8 |
| Trp | 35 a | 2.3 | 0.8 b | 0.2 | 0.7 b | 0.2 | 3.7 a | 0.6 | 4.8 a | 0.7 | 4.4 a | 0.7 |
| Total: | 953 a | 191 | 70 b | 14 | 47 b | 9 | 594 a | 119 | 608 a | 122 | 664 a | 13 |
| Non-Essential | ||||||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Glu + Gln | 591 a | 88 | 28 b | 4 | 31 b | 5 | 179 a | 27 | 153 a | 23 | 142 a | 21 |
| Pro | 275 a | 55 | 43 b | 9 | 27 c | 6 | 210 a | 42 | 189 a | 38 | 186 a | 37 |
| Asp + Asn | 240 a | 36 | 10 b | 2 | 14 b | 2 | 117 a | 18 | 118 a | 18 | 97 a | 15 |
| Ser | 111 a | 7 | 13 b | 1 | 4.7 c | 0.3 | 65 a | 4 | 57 a | 4 | 60 a | 4 |
| Tyr | 110 a | 4.9 | 4.6 b | 0.2 | 2.1 c | 0.1 | 27 a | 4 | 28 a | 4 | 7.8 a | 1 |
| Ala | 76 a | 11 | 19 b | 2.9 | 8.9 c | 1.3 | 149 a | 22 | 163 a | 24 | 158 a | 24 |
| Arg | 50 a | 8 | 2.7 b | 0.4 | 3.4 b | 0.5 | 63 a | 9 | 71 a | 11 | 54 a | 8 |
| Gly | 35 a | 2 | 3.1 b | 0.1 | 4.0 c | 0.2 | 206 a | 9 | 225 a | 10 | 228 a | 10 |
| Cys | 23 a | 3 | ND | - | ND | - | ND | - | ND | - | ND | - |
| Total: | 919 a | 184 | 96 b | 19 | 64 c | 13 | 837 a | 167 | 852 a | 170 | 791 a | 158 |
| Amino Acid | Free Amino Acids, mg∙(100 mL)−1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RSM | MRS | |||||||||||
| Control | L. helveticus H9 | L. paracasei ABK | Control | L. helveticus H9 | L. paracasei ABK | |||||||
| Essential | ||||||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Met | ND | - | 4.6 a | 1.1 | 2.3 b | 0.5 | 27 a | 6 | 21 a | 5 | 46 b | 11 |
| Lys | ND | - | 1.7 a | 0.3 | 2.0 a | 0.4 | 8.0 a | 1.4 | 2.0 b | 0.4 | 1.4 c | 0.2 |
| Leu + Ile | ND | - | ND | - | 0.56 a | 0.1 | 4.6 a | 0.8 | 0.29 b | 0.05 | 0.46 c | 0.08 |
| Trp | ND | - | 0.05 a | 0.01 | 0.17 b | 0.03 | 3.4 a | 0.7 | 0.12 b | 0.02 | 0.18 c | 0.03 |
| His | ND | - | ND | - | ND | - | 2.4 a | 0.5 | ND | - | ND | - |
| Val | ND | - | ND | - | ND | - | 1.6 a | 0.3 | ND | - | ND | - |
| Thr | ND | - | ND | - | ND | - | 1.3 a | 0.2 | ND | - | ND | - |
| Phe | ND | - | ND | - | ND | - | ND | - | ND | - | ND | - |
| Total: | ND | - | 6.4 a | 1.3 | 5.0a | 1.2 | 48 a | 10 | 23 b | 5 | 48 a | 10 |
| Non-Essential | ||||||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Glu + Gln | ND | - | 8.3 a | 1.7 | 5.6 b | 1.1 | 23 a | 5 | 3.3 b | 0.6 | 3.5 b | 0.7 |
| Asp + Asn | ND | - | 0.50 a | 0.10 | 1.1 b | 0.2 | 17 a | 3 | 0.84 b | 0.17 | 0.64 b | 0.13 |
| Pro | ND | - | 0.81 a | 0.15 | 0.58 b | 0.10 | 12 a | 2 | 0.69 b | 0.12 | 0.90 b | 0.16 |
| Arg | ND | - | 1.3 a | 0.3 | 1.6 a | 0.4 | 5.3 a | 1.2 | 1.3 b | 0.3 | 1.2 b | 0.3 |
| Gly | ND | - | 1.4 a | 0.2 | 0.84 b | 0.15 | 3.6 a | 0.6 | 0.86 b | 0.15 | 1.2 b | 0.2 |
| Ser | ND | - | 0.51 a | 0.09 | 0.11 b | 0.02 | 2.8 a | 0.5 | 0.24 b | 0.04 | 0.22 b | 0.04 |
| Ala | ND | - | 0.51 a | 0.09 | 0.28 b | 0.05 | 2.0 a | 0.4 | 0.62 b | 0.11 | 0.68 b | 0.12 |
| Cys | ND | - | ND | - | ND | - | 1.9 a | 0.5 | ND | - | ND | - |
| Tyr | ND | - | ND | - | ND | - | ND | - | ND | - | ND | - |
| Total: | ND | - | 13 a | 3 | 10 a | 2 | 68 a | 14 | 7.8 b | 0.16 | 8.3 c | 0.2 |
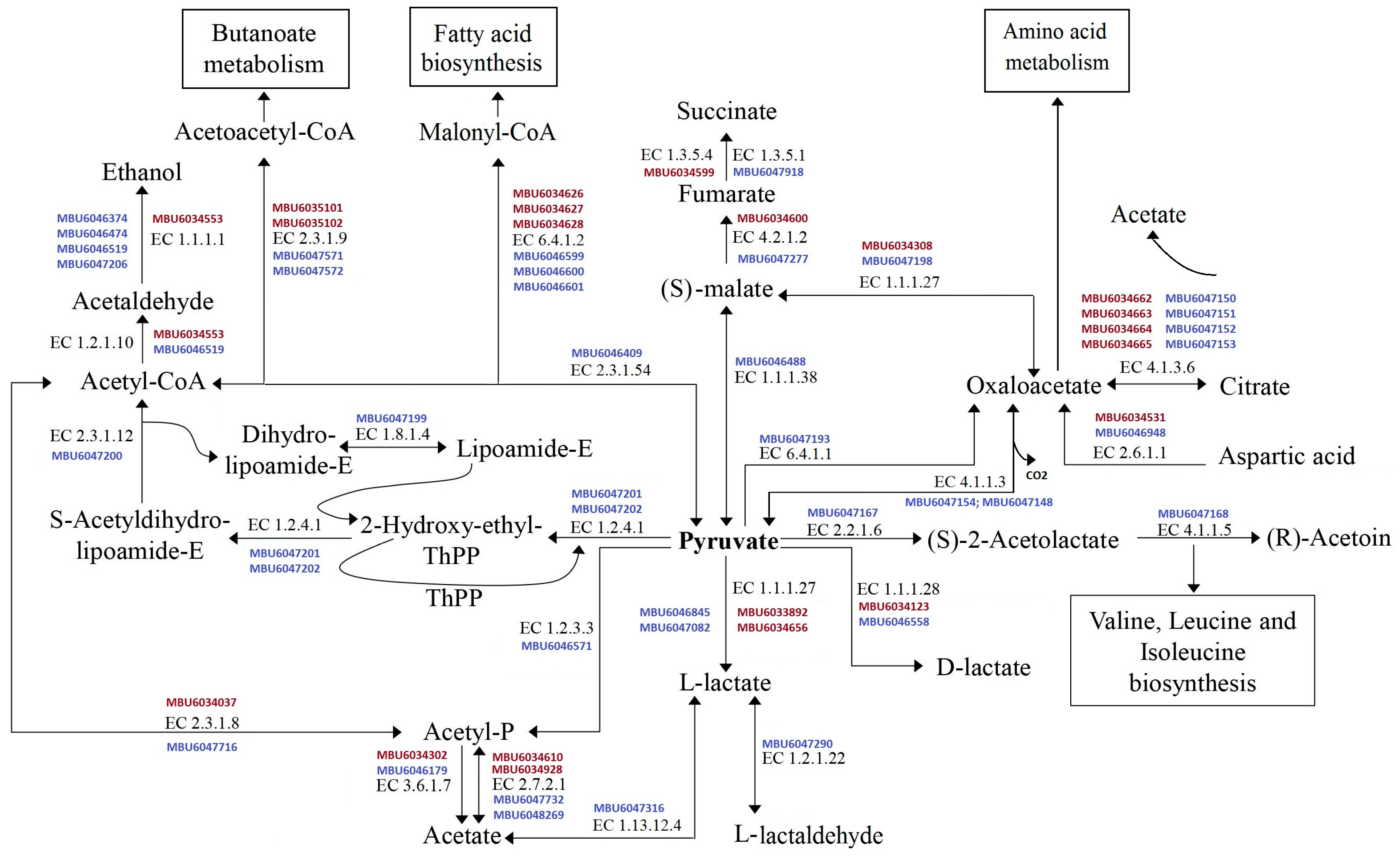
| Enzyme | Gene | L. helveticus H9 | L. paracasei ABK | ||
|---|---|---|---|---|---|
| Number of Genes | Locus Number in the Genome | Number of Genes | Locus Number in the Genome | ||
| CEP proteinases | prtB | 1 | MBU6033914 | 1 | MBU6046327 |
| prtP | 1 | MBU6034695 | 1 | MBU6048028 | |
| Endopeptidase | pepO | 2 | MBU6033720 MBU6034694 | 2 | MBU6046960 MBU6047360 |
| pepF | 1 | MBU6034722 | 3 | MBU6046494 MBU6047890 MBU6047627 | |
| pepE | 1 | MBU6034026 | 1 | MBU6047326 | |
| Aminopeptidases | pepC | 3 | MBU6034983 MBU6034023 MBU6034659 | 1 | MBU6047325 |
| pepN | 1 | MBU6034546 | 1 | MBU6048029 | |
| prolinase, pepP | 1 | MBU6035018 | 1 | MBU6046149 | |
| glutaminopeptidase, pepA | 1 | MBU6034400 | 0 | no * | |
| proline iminopeptidase, pepI | 1 | MBU6034385 | 1 | MBU6046842 | |
| prolidase, pepQ | 1 | MBU6034747 | 1 | MBU6046550 | |
| pepS | 0 | no | 1 | MBU6047768 | |
| Oligo-/Tri-/Di-peptidases | prolinase, pepR | 1 | MBU6034760 | 1 | MBU6047007 |
| tripeptidase, pepT | 2 | MBU6034325 MBU6034429 | 0 | no | |
| X-prolil dipeptidyl aminopeptidase, pepX | 1 | MBU6034686 | 1 | MBU6046155 | |
| dipepidase, pepV | 1 | MBU6034208 | 1 | MBU6047856 | |
| dipepidase, pepD | 4 | MBU6034175 MBU6033703 MBU6034538 MBU6034470 | 3 | MBU6046618 MBU6047722 MBU6047002 | |
| Total: | 23 | 20 | |||
| Sample | Content of Organic Acids, mg (100 mL)−1 | |||||
|---|---|---|---|---|---|---|
| Lactate | Acetate | Succinate | ||||
| RSM | ||||||
| Mean | SD | Mean | SD | Mean | SD | |
| Control | 36 a | 4 | 10 a | 1 | ND | - |
| L. helveticus H9 | 1371 b | 42 | 34 b | 3 | 45 | 2 |
| L. paracasei ABK | 1133 b | 32 | 31 b | 2 | ND | |
| MRS | ||||||
| Mean | SD | Mean | SD | Mean | SD | |
| Control | ND | - | 202 a | 11 | ND | - |
| L. helveticus H9 | 609 a | 22 | 226 a | 13 | 13 | 1 |
| L. paracasei ABK | 798 b | 34 | 217 a | 14 | ND | - |
| Gene | Enzyme Encoded by the Gene | EC: | Locus Number in the Genome | |
|---|---|---|---|---|
| L. helveticus H9 | L. paracasei ABK | |||
| Tagatose-6P pathway | ||||
| lacA | Galactose-6-phosphate isomerase | 5.3.1.26 | no * | MBU6047228 |
| lacB | Galactose-6-phosphate isomerase | 5.3.1.26 | no | MBU6047229 |
| lacC | Tagatose 6-phosphate kinase | 2.7.1.144 | no | MBU6047231 |
| lacD | Tagatose 1,6-diphosphate aldolase | 4.1.2.40 | no | MBU6047230 MBU6047617 |
| lacE | Lactose PTS system EIICB component | 2.7.1.207 | no | MBU6048344 |
| lacF | Lactose PTS system EIIA component | 2.7.1.207 | no | MBU6048346 |
| lacG | 6-phospho-beta-galactosidase | 3.2.1.85 | no | MBU6048345 |
| gatY | Tagatose 1,6-diphosphate aldolase | 4.1.2.40 | no | MBU6048319 |
| Leloir pathway | ||||
| galP lacS lacY | Galactose permease Galactose–Lactose antiporter Lactose permease | no | no | |
| lacL | Beta-galactosidase (GH2) | 3.2.1.23 | MBU6035121 | MBU6048422 |
| lacM | Beta-galactosidase | 3.2.1.23 | MBU6035122 | MBU6048423 |
| galM | Galactose epimerase | 5.1.3.3 | MBU6034706 | MBU6047737 |
| galK | Glucokinase | 2.7.1.6 | MBU6034704 | MBU6047741 |
| galT | Galactose-1-phosphate uridylyltransferase | 2.7.7.12 | MBU6034705 | MBU6047739 |
| galE | UDP-glucose-4-epimerase | 5.1.3.2 | MBU6035123 | MBU6047740 |
| pgm | Phosphoglucomutase | 5.4.2.2 | MBU6034768 | MBU6046814 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozhkova, I.V.; Yurova, E.A.; Leonova, V.A. Evaluation of the Amino Acid Composition and Content of Organic Acids of Complex Postbiotic Substances Obtained on the Basis of Metabolites of Probiotic Bacteria Lacticaseibacillus paracasei ABK and Lactobacillus helveticus H9. Fermentation 2023, 9, 460. https://doi.org/10.3390/fermentation9050460
Rozhkova IV, Yurova EA, Leonova VA. Evaluation of the Amino Acid Composition and Content of Organic Acids of Complex Postbiotic Substances Obtained on the Basis of Metabolites of Probiotic Bacteria Lacticaseibacillus paracasei ABK and Lactobacillus helveticus H9. Fermentation. 2023; 9(5):460. https://doi.org/10.3390/fermentation9050460
Chicago/Turabian StyleRozhkova, Irina Vladimirovna, Elena Anatolyevna Yurova, and Victoria Alexandrovna Leonova. 2023. "Evaluation of the Amino Acid Composition and Content of Organic Acids of Complex Postbiotic Substances Obtained on the Basis of Metabolites of Probiotic Bacteria Lacticaseibacillus paracasei ABK and Lactobacillus helveticus H9" Fermentation 9, no. 5: 460. https://doi.org/10.3390/fermentation9050460
APA StyleRozhkova, I. V., Yurova, E. A., & Leonova, V. A. (2023). Evaluation of the Amino Acid Composition and Content of Organic Acids of Complex Postbiotic Substances Obtained on the Basis of Metabolites of Probiotic Bacteria Lacticaseibacillus paracasei ABK and Lactobacillus helveticus H9. Fermentation, 9(5), 460. https://doi.org/10.3390/fermentation9050460