
Improvement of the Nutritional Quality of Rapeseed Meal through Solid-State Fermentation with B. subtilis, S. cerevisiae, and B. amyloliquefaciens
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Solid-State Fermentation
2.3. Measurement of Chemical Compositions
2.4. The Molecular Weight Distribution of the Proteins
2.5. Antioxidant Activity
2.6. Functional Properties
2.7. Correlations among Variables Analysis
2.8. Structural Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Effects of Fermentation on Anti-Nutritional Factors of RSM
3.2. Effects of Fermentation on Nutritional Components of RSM
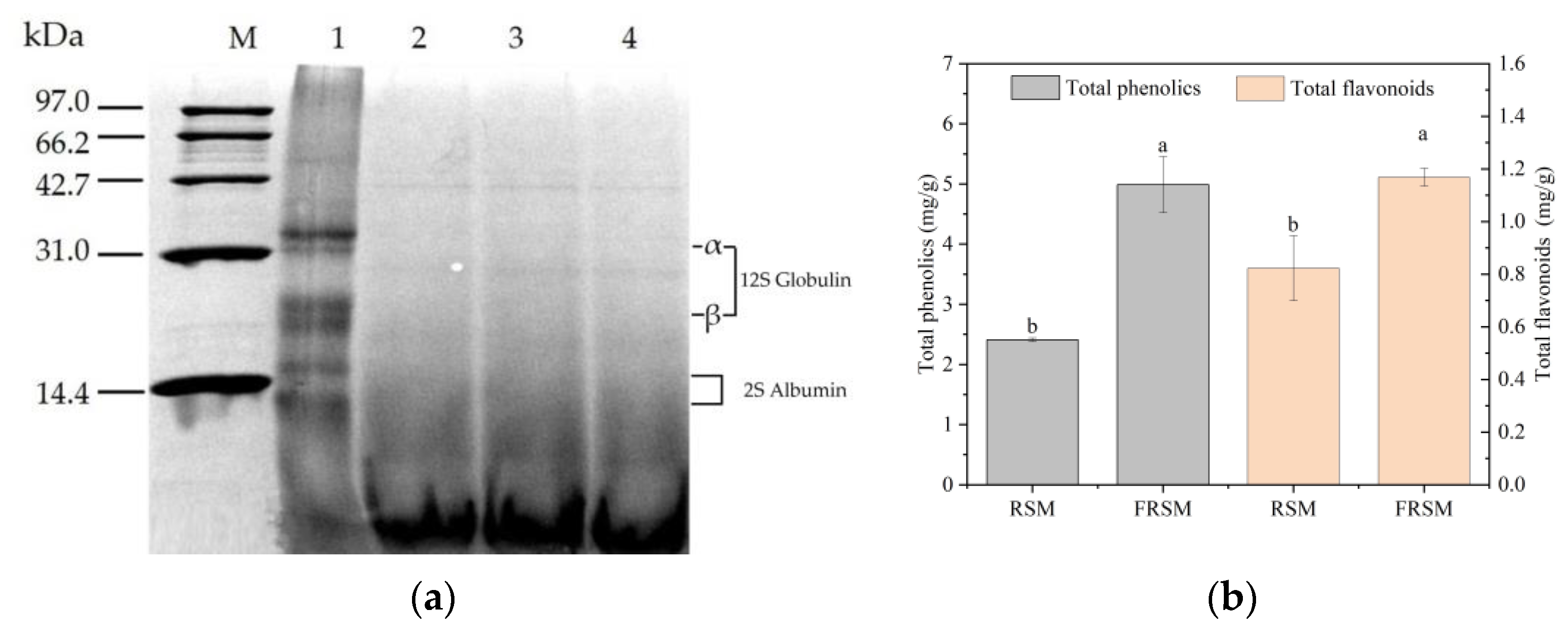
3.3. Effects of Fermentation on Protein Molecular Distribution of RSM
3.4. Effects of Fermentation on Bioactive Compounds of RSM
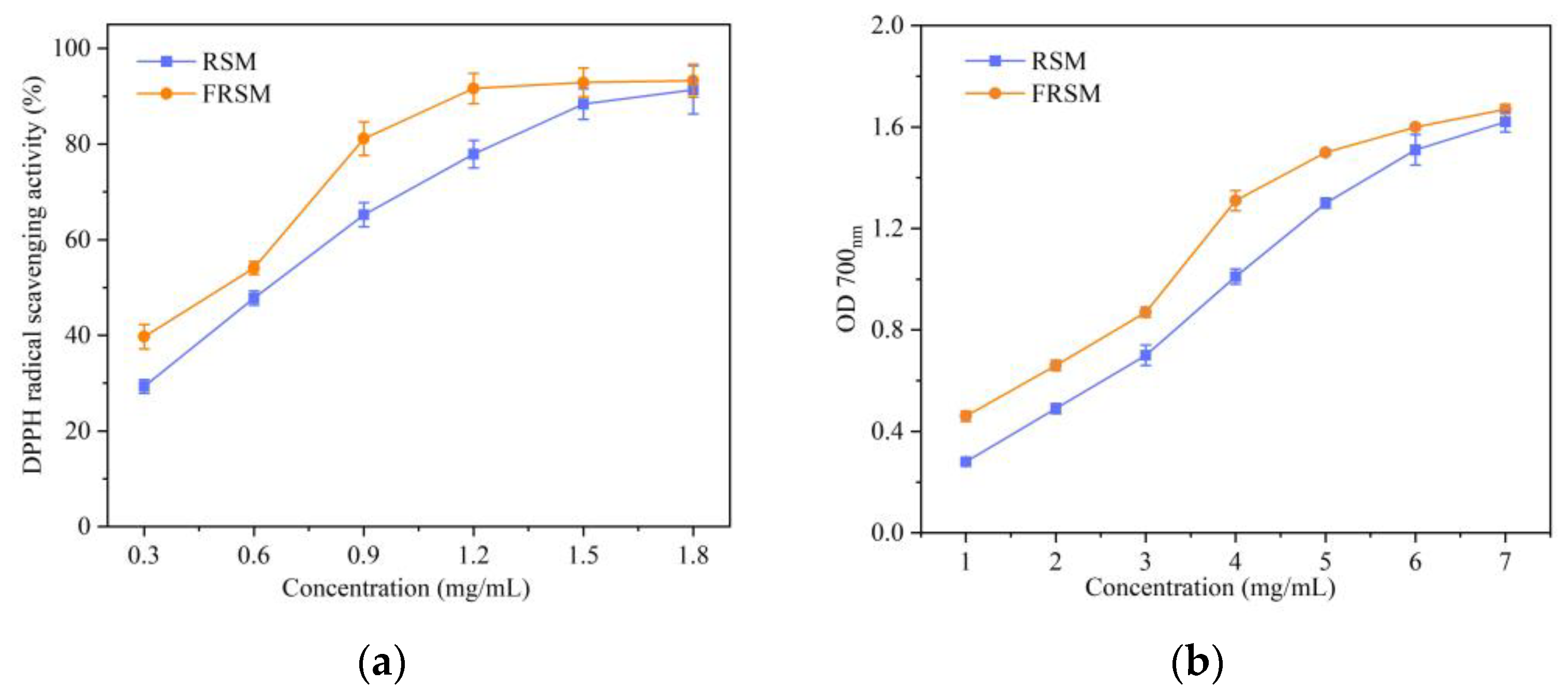
3.5. Effects of Fermentation on Antioxidant Activity of RSM
3.6. Effects of Fermentation on Functional Properties of RSM
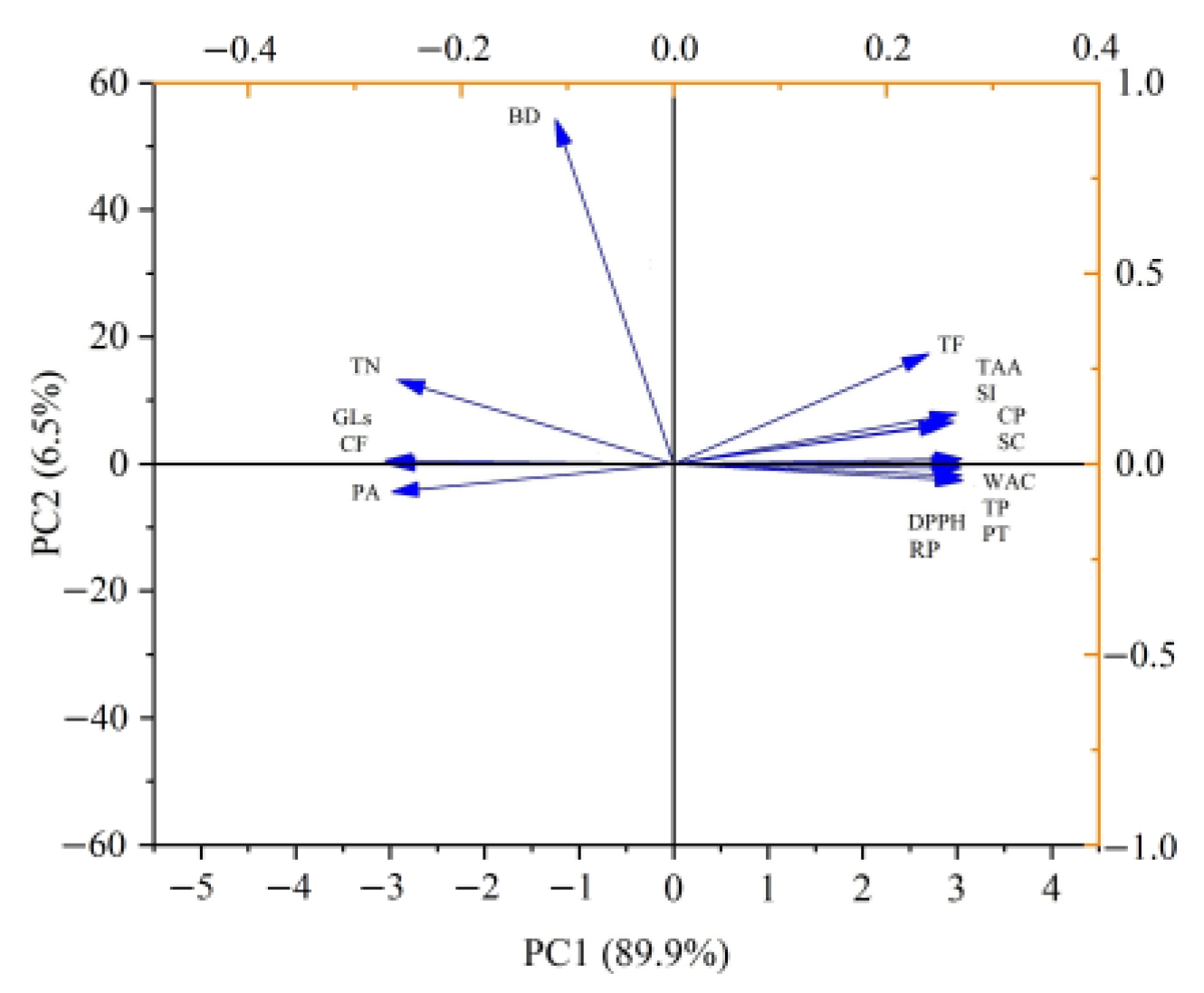
3.7. Correlations among Variables of RSM
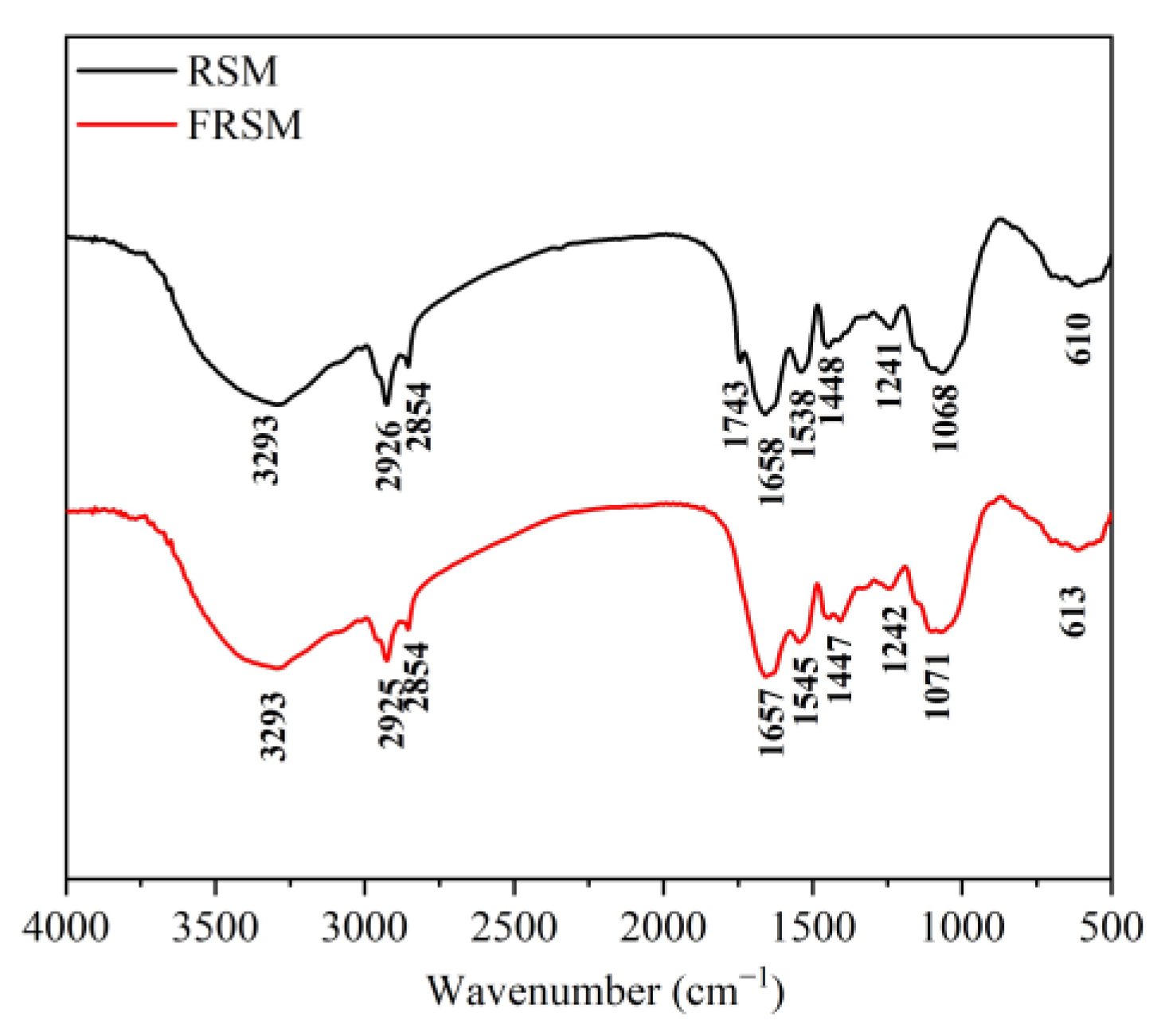
3.8. Effects of Fermentation on Functional Groups and Chemical Bonds of RSM
3.9. Effects of Fermentation on Microstructures of RSM
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yusuf, H.A.; Piao, M.; Ma, T.; Huo, R.; Tu, Y. Enhancing the Quality of Total Mixed Ration Containing Cottonseed or Rapeseed Meal by Optimization of Fermentation Conditions. Fermentation 2021, 7, 234. [Google Scholar] [CrossRef]
- Dossou, S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Dawood, M.A.O.; El Basuini, M.F.; El-Hais, A.M.; Olivier, A. Effect of Partial Replacement of Fish Meal by Fermented Rapeseed Meal on Growth, Immune Response and Oxidative Condition of Red Sea Bream Juvenile, Pagrus major. Aquaculture 2018, 490, 228–235. [Google Scholar] [CrossRef]
- Kaur, H.; Wang, L.; Stawniak, N.; Sloan, R.; van Erp, H.; Eastmond, P.; Bancroft, I. The Impact of Reducing Fatty Acid Desaturation on the Composition and Thermal Stability of Rapeseed oil. Plant. Biotechnol. J. 2020, 18, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Liu, X.; Xiao, Q.; Zhang, F.; Liu, N.; Tang, L.; Wang, J.; Ma, X.; Tan, B.; Chen, J.; et al. Rapeseed Meal and Its Application in Pig Diet: A Review. Agriculture 2022, 12, 849. [Google Scholar] [CrossRef]
- Wongsirichot, P.; Gonzalez-Miquel, M.; Winterburn, J. Recent Advances in Rapeseed Meal as Alternative Feedstock for Industrial Biotechnology. Biochem. Eng. J. 2022, 180, 108373. [Google Scholar] [CrossRef]
- Xie, C.; Li, W.; Gao, R.; Yan, L.; Wang, P.; Gu, Z.; Yang, R. Determination of Glucosinolates in Rapeseed Meal and Their Degradation by Myrosinase from Rapeseed Sprouts. Food Chem. 2022, 382, 132316. [Google Scholar] [CrossRef]
- Yang, Z.; Huang, Z.; Cao, L. Biotransformation Technology and High-value Application of Rapeseed Meal: A Review. Bioresour. Bioprocess. 2022, 9, 103. [Google Scholar] [CrossRef]
- Hao, Y.; Wang, Z.; Zou, Y.; He, R.; Ju, X.; Yuan, J. Effect of Static-state Fermentation on Volatile Composition in Rapeseed Meal. J. Sci. Food Agric. 2020, 100, 2145–2152. [Google Scholar] [CrossRef]
- Konkol, D.; Szmigiel, I.; Domzal-Kedzia, M.; Kulazynski, M.; Krasowska, A.; Opalinski, S.; Korczynski, M.; Lukaszewicz, M. Biotransformation of Rapeseed Meal Leading to Production of Polymers, Biosurfactants, and Fodder. Bioorg. Chem. 2019, 93, 102865. [Google Scholar] [CrossRef] [PubMed]
- Khajali, F.; Slominski, B.A. Factors that Affect the Nutritive Value of Canola Meal for Poultry. Poult. Sci. 2012, 91, 2564–2575. [Google Scholar] [CrossRef]
- Yao, J.; Chen, P.; Ringo, E.; Zhang, G.; Huang, Z.; Hua, X. Effect of Diet Supplemented with Rapeseed Meal or Hydrolysable Tannins on the Growth, Nutrition, and Intestinal Microbiota in Grass Carp (Ctenopharyngodon idellus). Front. Nutr. 2019, 6, 154. [Google Scholar] [CrossRef] [PubMed]
- Olukomaiya, O.O.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Solid-state Fermentation of Canola Meal with Aspergillus sojae, Aspergillus ficuum and Their co-cultures: Effects on Physicochemical, Microbiological and Functional properties. LWT 2020, 127, 109362. [Google Scholar] [CrossRef]
- Olukomaiya, O.; Fernando, C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Solid-state Fermented Plant Protein Sources in the Diets of Broiler Chickens: A Review. Anim. Nutr. 2019, 5, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Lomascolo, A.; Uzan-Boukhris, E.; Sigoillot, J.C.; Fine, F. Rapeseed and Sunflower Meal: A Review on Biotechnology Status and Challenges. Appl. Microbiol. Biotechnol. 2012, 95, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Czech, A.; Stępniowska, A.; Kiesz, M. Effect of Fermented Rapeseed Meal as a Feed Component on the Redox and Immune System of Pregnant Sows and Their Offspring. Ann. Anim. Sci. 2022, 22, 201–219. [Google Scholar] [CrossRef]
- Bu, X.; Wang, Y.; Chen, F.; Tang, B.; Luo, C.; Wang, Y.; Ge, X.; Yang, Y. An Evaluation of Replacing Fishmeal with Rapeseed Meal in the Diet of Pseudobagrus ussuriensis: Growth, Feed Utilization, Nonspecific Immunity, and Growth-related Gene Expression. J. World Aquac. Soc. 2018, 49, 1068–1080. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S. Application of Fermentation Strategy in Aquafeed for Sustainable Aquaculture. Rev. Aquac. 2019, 12, 987–1002. [Google Scholar] [CrossRef]
- Vig, A.P.; Walia, A. Beneficial Effects of Rhizopus oligosporus Fermentation on Reduction of Glucosinolates, Fibre and Phytic acid in Rapeseed (Brassica napus) Meal. Bioresour. Technol. 2001, 78, 309–312. [Google Scholar]
- Wang, Y.; Liu, J.; Wei, F.; Liu, X.; Yi, C.; Zhang, Y. Improvement of the Nutritional Value, Sensory Properties and Bioavailability of Rapeseed Meal Fermented with Mixed Microorganisms. LWT 2019, 112, 108238. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, H.; Han, B.; Li, H.Y.; Liu, X.L. Improvement of Nutritional Value, Molecular Weight Patterns (soluble peptides), Free Amino Acid Patterns, Total Phenolics and Antioxidant Activity of Fermented Extrusion Pretreatment Rapeseed meal with Bacillus subtilis YY-1 and Saccharomyces cerevisiae YY-2. LWT 2022, 160, 113280. [Google Scholar] [CrossRef]
- Vlassa, M.; Filip, M.; Taranu, I.; Marin, D.; Untea, A.E.; Ropota, M.; Dragomir, C.; Saracila, M. The Yeast Fermentation Effect on Content of Bioactive, Nutritional and Anti-Nutritional Factors in Rapeseed Meal. Foods 2022, 11, 2972. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wu, X.B.; Chen, H.L.; Sun-Waterhouse, D.; Zhong, H.B.; Cui, C. A Value-added Approach to Improve the Nutritional Quality of Soybean Meal Byproduct: Enhancing Its Antioxidant Activity Through Fermentation by Bacillus amyloliquefaciens SWJS22. Food Chem. 2019, 272, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Ngalimat, M.S.; Yahaya, R.S.R.; Baharudin, M.M.A.; Yaminudin, S.M.; Karim, M.; Ahmad, S.A.; Sabri, S. A Review on the Biotechnological Applications of the Operational Group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef]
- Yang, L.; Zeng, X.; Qiao, S. Advances in Research on Solid-state Fermented Feed and Its Utilization: The Pioneer of Private Customization for Intestinal Microorganisms. Anim. Nutr. 2021, 7, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, H.; Liu, X. A Novel Fermented Rapeseed Meal, Inoculated with Selected Protease-Assisting Screened B. subtilis YY-4 and L. plantarum 6026, Showed High Availability and Strong Antioxidant and Immunomodulation Potential Capacity. Foods 2022, 11, 2118. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; He, J.; Yu, J.; Yu, B.; Mao, X.; Zheng, P.; Huang, Z.; Chen, D. Physicochemical Properties Analysis and Secretome of Aspergillus niger in Fermented Rapeseed Meal. PLoS ONE 2016, 11, e0153230. [Google Scholar] [CrossRef]
- Rawdkuen, S.; Sai-Ut, S.; Khamsorn, S.; Chaijan, M.; Benjakul, S. Biochemical and Gelling Properties of Tilapia Surimi and Protein Recovered Using an Acid-alkaline Process. Food Chem. 2009, 112, 112–119. [Google Scholar] [CrossRef]
- Sotelo, A.; Gonzalez-Osnaya, L.; Sanchez-Chinchillas, A.; Trejo, A. Role of Oxate, Phytate, Tannins and Cooking on Iron Bioavailability from Foods Commonly Consumed in Mexico. Int. J. Food Sci. Nutr. 2010, 61, 29–39. [Google Scholar] [CrossRef]
- Sugiharto, S.; Yudiarti, T.; Isroli, I. Assay of Antioxidant Potential of Two Filamentous Fungi Isolated from the Indonesian Fermented Dried Cassava. Antioxidants 2016, 5, 6. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar] [CrossRef]
- Oskoueian, E.; Abdullah, N.; Hendra, R.; Karimi, E. Bioactive compounds, antioxidant, xanthine oxidase inhibitory, tyrosinase inhibitory and anti-inflammatory activities of selected agro-industrial by-products. Int. J. Mol. Sci. 2011, 12, 8610–8625. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Song, M.; Wang, N.; Wang, Y.; Wang, R.; An, X.; Qi, J. The Effects of Solid-state Fermentation on the Content, Composition and in Vitro Antioxidant Activity of Flavonoids from Dandelion. PLoS ONE 2020, 15, e0239076. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Singh, S.; Kumari, D. Evaluation of Functional Properties of Composite Flours and Sensorial Attributes of Composite Flour Biscuits. J. Food Sci. Technol. 2015, 52, 3681–3688. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ji, X.; Deng, X.; Hu, S.; Wang, J.; Ding, K.; Liu, N. Effect of Rapeseed Meal Degraded by Enzymolysis and Fermentation on the Growth Performance, Nutrient Digestibility and Health Status of Broilers. Arch. Anim. Nutr. 2023, 76, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, Y.; Li, A.; Wang, Z.; Zhang, X.; Yun, T.; Qiu, L.; Yin, Y. Effects of Fermented Rapeseed Meal on Antioxidant Functions, Serum Biochemical Parameters and Intestinal Morphology in Broilers. Food Agric. Immunol. 2015, 27, 182–193. [Google Scholar] [CrossRef]
- Plaipetch, P.; Yakupitiyage, A. Effect of Replacing Soybean Meal with Yeast-fermented Canola Meal on Growth and Nutrient Retention of Nile tilapia, Oreochromis niloticus (Linnaeus 1758). Aquac. Res. 2014, 45, 1744–1753. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Q.; Huang, Z.; Wang, Y.; Roubik, H.; Yang, K.; Cai, M.; Sun, P. Solid-State Fermentation of Soybean Meal with Edible Mushroom Mycelium to Improve Its Nutritional, Antioxidant Capacities and Physicochemical Properties. Fermentation 2023, 9, 322. [Google Scholar] [CrossRef]
- Pereira, R.T.; Rosa, P.V.; Gatlin, D.M. Glutamine and arginine in diets for Nile tilapia: Effects on growth, innate immune responses, plasma amino acid profiles and whole-body composition. Aquaculture 2017, 473, 135–144. [Google Scholar] [CrossRef]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of Fermentation in Improving Nutritional Quality of Soybean Meal—A Review. Asian-Australas. J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef]
- Cho, K.M.; Lee, J.H.; Yun, H.D.; Ahn, B.Y.; Kim, H.; Seo, W.T. Changes of phytochemical constituents (isoflavones, flavanols, and phenolic acids) during cheonggukjang soybeans fermentation using potential probiotics Bacillus subtilis CS90. J. Food Compos. Anal. 2011, 24, 402–410. [Google Scholar] [CrossRef]
- Ballus, C.A.; Meinhart, A.D.; de Souza Campos, F.A.; Godoy, H.T. Total Phenolics of Virgin Olive Oils Highly Correlate with the Hydrogen Atom Transfer Mechanism of Antioxidant Capacity. J. Am. Oil Chem. Soc. 2015, 92, 843–851. [Google Scholar] [CrossRef]
- Sreerama, Y.N.; Sasikala, V.B.; Pratape, V.M. Nutritional Implications and Flour Functionality of Popped/Expanded Horse Gram. Food Chem. 2008, 108, 891–899. [Google Scholar] [CrossRef]
- Odey, G.N.; Lee, W.Y. Evaluation of the Quality Characteristics of Flour and Pasta from Fermented Cassava Roots. Int. J. Food Sci. Technol. 2019, 55, 813–822. [Google Scholar] [CrossRef]
- Ghumman, A.; Kaur, A.; Singh, N. Impact of Germination on Flour, Protein and Starch Characteristics of Lentil (Lens culinari) and Horsegram (Macrotyloma uniflorum L.) Lines. LWT 2016, 65, 137–144. [Google Scholar] [CrossRef]
- Zhao, Q.; Yan, X.; Yue, Y.; Yue, T.; Yuan, Y. Improved flavonoid content in mulberry leaves by solid-state fermentation: Metabolic profile, activity, and mechanism. Innov. Food Sci. Emerg. Technol. 2023, 84, 103308. [Google Scholar] [CrossRef]
- Ma, M.; Mu, T. Modification of deoiled cumin dietary fiber with laccase and cellulase under high hydrostatic pressure. Carbohydr. Polym. 2016, 136, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Alokika; Singh, B. Enhanced production of bacterial xylanase and its utility in saccharification of sugarcane bagasse. Bioprocess. Biosyst. Eng. 2020, 43, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.P.; Shen, Q.R.; Yu, G.H.; Ran, W.; Xu, Y.C. Fate of biopolymers during rapeseed meal and wheat bran composting as studied by two-dimensional correlation spectroscopy in combination with multiple fluorescence labeling techniques. Bioresour. Technol. 2012, 105, 88–94. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Z.; Dai, C.; Wang, Y.; Chen, W.; Ju, X.; Yuan, J.; He, R. Physical stability and microstructure of rapeseed protein isolate/gum Arabic stabilized emulsions at alkaline pH. Food Hydrocoll. 2019, 88, 50–57. [Google Scholar] [CrossRef]
- Yasar, S.; Tosun, R.; Sonmez, Z. Fungal fermentation inducing improved nutritional qualities associated with altered secondary protein structure of soybean meal determined by FTIR spectroscopy. Measurement 2020, 161, 107895. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anti-Nutritional Factors | RSM | FRSM | SEM | Degradation Rate (%) | p |
|---|---|---|---|---|---|
| Glucosiolates (μmol/g) | 47.67 a ± 0.70 | 0.26 b ± 0.1 | 10.60 | 99.18 | <0.01 |
| Phytic acid (%) | 3.11 a ± 0.18 | 1.79 b ± 0.12 | 0.30 | 42.41 | <0.01 |
| Crude fiber (%) | 11.25 a ± 0.26 | 8.19 b ± 0.07 | 0.69 | 27.21 | <0.01 |
| Tannins (%) | 2.67 a ± 0.15 | 1.76 b ± 0.02 | 0.21 | 34.17 | <0.01 |
| Composition (%) | RSM | FRSM | SEM | Increase Rate(%) | p |
|---|---|---|---|---|---|
| Crude protein | 33.16 a ±1.39 | 38.56 b ±1.51 | 1.32 | 16.28 | <0.05 |
| Peptides | 4.13 a ± 0.17 | 16.47 b ± 0.31 | 2.76 | 298.79 | <0.01 |
| Asp | 2.65 b ± 0.06 | 3.01 a ± 0.06 | 0.08 | 13.57 | <0.01 |
| Ile | 1.54 a ± 0.07 | 1.51 a ± 0.05 | 0.02 | −1.74 | >0.05 |
| Leu | 2.42 b ± 0.06 | 2.91 a ± 0.06 | 0.10 | 20.08 | <0.01 |
| Lys | 1.77 a ± 0.05 | 1.84 a ± 0.05 | 0.02 | 4.14 | >0.05 |
| Met | 0.33 a ± 0.04 | 0.37 a ± 0.05 | 0.02 | 10.00 | >0.05 |
| Phe | 1.84 a ± 0.06 | 1.85 a ± 0.04 | 0.01 | 0.54 | >0.05 |
| Thr | 1.45 b ± 0.05 | 1.82 a ± 0.08 | 0.08 | 25.00 | <0.01 |
| Val | 2.16 a ± 0.06 | 2.16 a ± 0.06 | 0.02 | 0.00 | >0.05 |
| Ser | 1.31 b ± 0.04 | 1.73 a ± 0.02 | 0.10 | 32.32 | <0.01 |
| Glu | 5.69 b ± 0.15 | 7.62 a ± 0.05 | 0.39 | 33.86 | <0.01 |
| Gly | 1.60 b ± 0.09 | 1.97 a ± 0.07 | 0.09 | 22.66 | <0.01 |
| Ala | 1.59 b ± 0.08 | 1.79 a ± 0.06 | 0.05 | 12.13 | <0.05 |
| Cys | 0.70 a ± 0.06 | 0.74 a ± 0.04 | 0.02 | 5.74 | >0.05 |
| His | 0.79 b ± 0.07 | 1.15 a ± 0.03 | 0.08 | 44.96 | <0.01 |
| Arg | 1.47 b ± 0.07 | 2.27 a ± 0.04 | 0.18 | 54.20 | <0.01 |
| Pro | 2.64 b ± 0.10 | 2.84 a ± 0.06 | 0.05 | 7.44 | <0.05 |
| Tyr | 0.93 a ± 0.06 | 0.92 a ± 0.06 | 0.02 | −1.79 | >0.05 |
| EAAs | 14.16 a ± 0.35 | 15.47 a ± 0.34 | 0.32 | 9.18 | >0.05 |
| NEAAs | 16.74 b ± 0.56 | 21.01 a ± 0.34 | 0.98 | 25.58 | <0.01 |
| TAAs | 30.90 b ± 0.91 | 36.48 a ± 0.69 | 1.29 | 18.06 | <0.01 |
| Parameters | RSM | FRSM | SEM | p |
|---|---|---|---|---|
| Bulk density (g/mL) | 0.54 a ± 0.02 | 0.53 a ± 0.01 | 0.00 | >0.05 |
| Water absorption capacity (%) | 227.08 b ± 5.07 | 281.78 a ± 10.84 | 12.62 | <0.01 |
| Swelling index | 1.29 b ± 0.08 | 1.66 a ± 0.05 | 0.08 | <0.01 |
| Swelling capacity (mL) | 14.00 b ± 0.20 | 24.50 a ± 0.50 | 2.35 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Chen, Y.; Hao, S.; Jin, S.; Li, X. Improvement of the Nutritional Quality of Rapeseed Meal through Solid-State Fermentation with B. subtilis, S. cerevisiae, and B. amyloliquefaciens. Fermentation 2023, 9, 492. https://doi.org/10.3390/fermentation9050492
Zhu X, Chen Y, Hao S, Jin S, Li X. Improvement of the Nutritional Quality of Rapeseed Meal through Solid-State Fermentation with B. subtilis, S. cerevisiae, and B. amyloliquefaciens. Fermentation. 2023; 9(5):492. https://doi.org/10.3390/fermentation9050492
Chicago/Turabian StyleZhu, Xiaoyan, Yiling Chen, Shanxin Hao, Shiyu Jin, and Xiangqian Li. 2023. "Improvement of the Nutritional Quality of Rapeseed Meal through Solid-State Fermentation with B. subtilis, S. cerevisiae, and B. amyloliquefaciens" Fermentation 9, no. 5: 492. https://doi.org/10.3390/fermentation9050492
APA StyleZhu, X., Chen, Y., Hao, S., Jin, S., & Li, X. (2023). Improvement of the Nutritional Quality of Rapeseed Meal through Solid-State Fermentation with B. subtilis, S. cerevisiae, and B. amyloliquefaciens. Fermentation, 9(5), 492. https://doi.org/10.3390/fermentation9050492