

Production of Glyoxylate from Glucose in Engineered Escherichia coli
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Disruption
2.2. Plasmid Construction
2.3. Fermentation Production
2.4. Analytical Methods
3. Results and Discussion
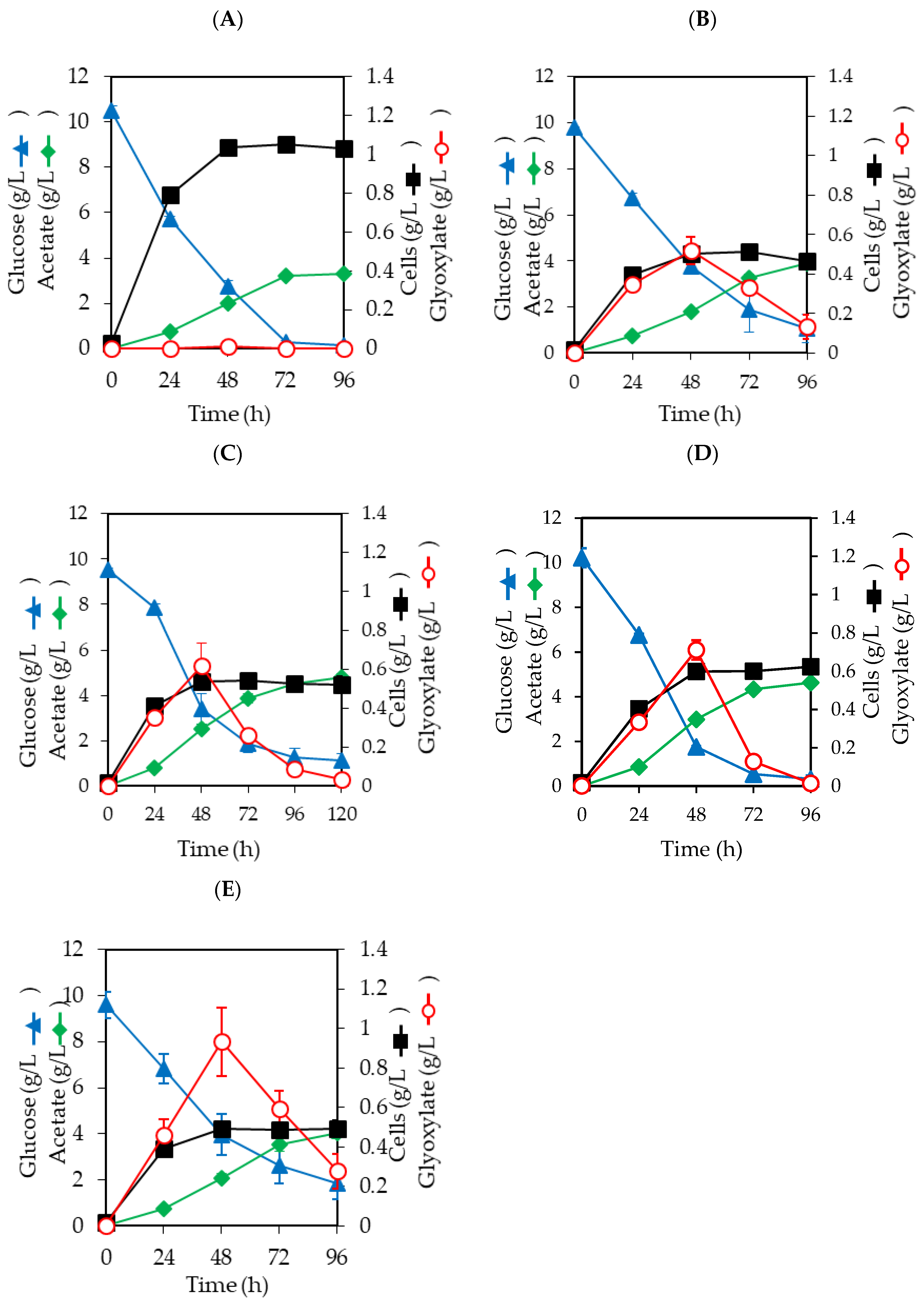
3.1. Effects of Gene Disruption on Glyoxylate Production
3.2. Effects of Gene Overexpression on Glyoxylate Production
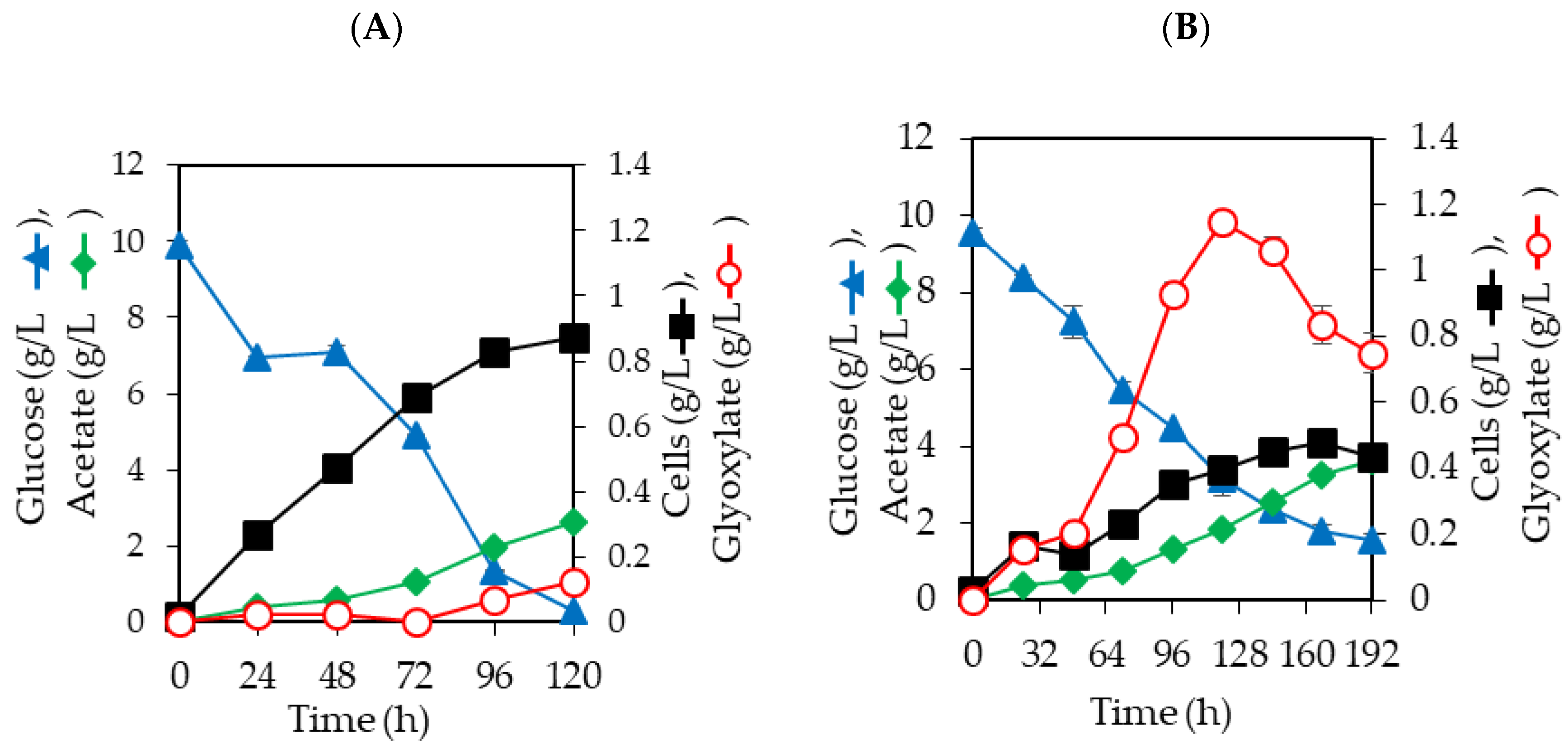
3.3. Effects of pyc Expression on Glyoxylate Production in E. coli
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skarstad, K.; Boye, E.; Steen, H.B. Timing of initiation of chromosome replication in individual Escherichia coli cells. EMBO J. 1986, 5, 1711–1717. [Google Scholar] [CrossRef] [PubMed]
- Castiñeiras, T.S.; Williams, S.G.; Hitchcock, A.G.; Smith, D.C. E. coli strain engineering for the production of advanced biopharmaceutical products. FEMS Microbiol. Lett. 2018, 365, fny162. [Google Scholar] [CrossRef] [PubMed]
- Sauer, U.; Eikmanns, B. The PEP-pyruvate-oxaloacetate node as the switch point for carbon flux distribution in bacteria. FEMS Microbiol. Rev. 2005, 29, 765–794. [Google Scholar] [CrossRef]
- Aso, Y.; Tsubaki, M.; Long, B.H.D.; Murakami, R.; Nagata, K.; Okano, H.; Dung, N.T.P.; Ohara, H. Continuous production of D-lactic acid from cellobiose in cell recycle fermentation using β-glucosidase-displaying Escherichia coli. J. Biosci. Bioeng. 2019, 127, 441–446. [Google Scholar] [CrossRef]
- Förster, A.H.; Gescher, J. Metabolic engineering of Escherichia coli for production of mixed-acid fermentation end products. Front. Bioeng. Biotechnol. 2014, 2, 16. [Google Scholar] [PubMed]
- Okamoto, S.; Chin, T.; Nagata, K.; Takahashi, T.; Ohara, H.; Aso, Y. Production of itaconic acid in Escherichia coli expressing recombinant α-amylase using starch as substrate. J. Biosci. Bioeng. 2015, 119, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Sato, R.; Tanaka, T.; Ohara, H.; Aso, Y. Engineering Escherichia coli for direct production of 1,2-propanediol and 1,3-propanediol from starch. Curr. Microbiol. 2020, 77, 3704–3710. [Google Scholar] [CrossRef]
- Sato, R.; Tanaka, T.; Ohara, H.; Aso, Y. Disruption of glpF gene encoding the glycerol facilitator improves 1,3-propanediol production from glucose via glycerol in Escherichia coli. Lett. Appl. Microbiol. 2021, 72, 68–73. [Google Scholar] [CrossRef]
- Gupta, N.K.; Vennesland, B. Glyoxylate carboligase of Escherichia coli: A flavoprotein. J. Biol. Chem. 1964, 239, 3787–3789. [Google Scholar] [CrossRef]
- Pozdniakov, M.A.; Zhuk, I.V.; Lyapunova, M.V.; Salikov, A.S.; Botvin, V.V.; Filimoshkin, A.G. Glyoxylic acid: Synthesis, isolation, and crystallization. Russ. Chem. Bull. 2019, 68, 472–479. [Google Scholar] [CrossRef]
- Nishizawa, R.; Saino, T.; Suzuki, M.; Fujii, T.; Shirai, T.; Aoyagi, T.; Umezawa, H. A facile synthesis of bestatin. J. Antibiot. 1983, 36, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; He, A.; Li, X. Synthesis of Allantoin Catalyzed by SO42−/La2O3-SiO2-ZrO2. Asian J. Chem. 2012, 24, 2298–2300. [Google Scholar]
- Mattioda, G.; Christidis, Y. Glyoxylic Acid. In Ullmann’s Encyclopedia of Industrial Chemistry, 6th ed.; Elvers, B., Ed.; Wiley-VCH: Weinheim, Germany, 2000; pp. 89–92. [Google Scholar]
- Kalikar, R.G.; Deshpande, R.S.; Chandalia, S.B. Synthesis of vanillin and 4-hydroxybenzaldehyde by a reaction scheme involving condensation of phenols with glyoxylic acid. J. Chem. Technol. Biotechnol. 1986, 36, 38–46. [Google Scholar] [CrossRef]
- Sun, Z.C.; Eli, W.; Xu, T.Y.; Zhang, Y.G. Oxidation of glyoxal with hydroperoxide compounds prepared from maleic acid by ozonation to produce glyoxylic acid. Ind. Eng. Chem. Res. 2006, 45, 1849–1852. [Google Scholar] [CrossRef]
- Jin, J.; Tan, T.; Wang, H.; Su, G. The expression of spinach glycolate oxidase (GO) in E. coli and the application of GO in the production of glyoxylic acid. Mol. Biotechnol. 2003, 25, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Section 17.4: The Glyoxylate Cycle Enables Plants and Bacteria to Grow on Acetate. In Biochemistry, 5th ed.; Berg, J.M., Tymoczko, J.L., Stryer, L., Eds.; W. H. Freeman: New York, NY, USA, 2002. [Google Scholar]
- Dunn, M.F.; Ramírez-Trujillo, J.A.; Hernández-Lucas, I. Major roles of isocitrate lyase and malate synthase in bacterial and fungal pathogenesis. Microbiology 2009, 155, 3166–3175. [Google Scholar] [CrossRef] [PubMed]
- Sneeden, R.P.A. 50.2—Organic Syntheses where Carbon Monoxide is the Unique Source of Carbon. In Comprehensive Organometallic Chemistry; Wilkinson, G., Stone, F.G., Abel, E.W., Eds.; Pergamon: Oxford, UK, 1982; pp. 19–100. [Google Scholar]
- Li, L.K.; Shi, L.L.; Hong, P.H.; Tan, T.W.; Li, Z.J. Metabolic engineering of Escherichia coli for the production of glyoxylate from xylose. Biochem. Eng. J. 2018, 129, 113–118. [Google Scholar] [CrossRef]
- Foyer, C.H.; Bloom, A.J.; Queval, G.; Noctor, G. Photorespiratory metabolism: Genes, mutants, energetics, and redox signaling. Annu. Rev. Plant Biol. 2009, 60, 455–484. [Google Scholar] [CrossRef]
- Rojas, C.M.; Mysore, K.S. Glycolate oxidase is an alternative source for H2O2 production during plant defense responses and functions independently from NADPH oxidase. Plant Signal. Behav. 2012, 7, 752–755. [Google Scholar] [CrossRef]
- Rojas, C.M.; Senthil-Kumar, M.; Wang, K.; Ryu, C.M.; Kaundal, A.; Mysore, K.S. Glycolate oxidase modulates reactive oxygen species-mediated signal transduction during nonhost resistance in Nicotiana benthamiana and Arabidopsis. Plant Cell 2012, 24, 336–352. [Google Scholar] [CrossRef]
- Gavagan, J.E.; Fager, S.K.; Seip, J.E.; Payne, M.S.; Anton, D.L.; DiCosimo, E. Glyoxylic acid production using microbial transformant catalysts. J. Org. Chem. 1995, 60, 3957–3963. [Google Scholar] [CrossRef]
- Zhu, T.; Yao, D.; Li, D.; Xu, H.; Jia, S.; Bi, C.; Cai, J.; Zhu, X.; Zhang, X. Multiple strategies for metabolic engineering of Escherichia coli for efficient production of glycolate. Biotechnol. Bioeng. 2021, 118, 4699–4707. [Google Scholar] [CrossRef]
- Martínez, I.; Bennett, G.N.; San, K.Y. Metabolic impact of the level of aeration during cell growth on anaerobic succinate production by an engineered Escherichia coli strain. Metab. Eng. 2010, 12, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zheng, T.; Ye, X.; Zheng, T.; Ye, X.; Xin, F.; Zhang, W.; Dong, W.; Ma, J.; Jiang, M. Metabolic engineering of Escherichia coli for L-malate production anaerobically. Microb. Cell Fact. 2010, 19, 165. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ma, D.; Liu, J.; Luo, Q.; Liu, L. Engineering the transmission efficiency of the noncyclic glyoxylate pathway for fumarate production in Escherichia coli. Biotechnol. Biofuels. 2020, 13, 132. [Google Scholar] [CrossRef]
- Li, X.T.; Thomason, L.C.; Sawitzke, J.A.; Costantino, N.; Court, D.L. Positive and negative selection using the tetA-sacB cassette: Recombineering and P1 transduction in Escherichia coli. Nucleic Acids Res. 2013, 41, e204. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Higuchi, R.; Krummel, B.; Saiki, R. A general method of in vitro preparation and specific mutagenesis of DNA fragments: Study of protein and DNA interactions. Nucleic Acids Res. 1988, 16, 7351–7367. [Google Scholar] [CrossRef]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Froger, A.; Hall, J.E. Transformation of plasmid DNA into E. coli using the heat shock method. J. Vis. Exp. 2007, 6, 253. [Google Scholar]
- Lange, M.; Mályusz, M. Fast method for the simultaneous determination of 2-oxo acids in biological fluids by high-performance liquid chromatography. J. Chromatogr. B Biomed. 1994, 662, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Canelas, A.B.; ten Pierick, A.; Ras, C.; Seifar, R.M.; van Dam, J.C.; van Gulik, W.M.; Heijnen, J.J. Quantitative evaluation of intracellular metabolite extraction techniques for yeast metabolomics. Anal. Chem. 2009, 81, 7379–7389. [Google Scholar] [CrossRef]
- Lassalle, L.; Engilberge, S.; Madern, D.; Vauclare, P.; Franzetti, B.; Girard, E. New insights into the mechanism of substrates trafficking in glyoxylate/hydroxypyruvate reductases. Sci. Rep. 2016, 6, 20629. [Google Scholar] [CrossRef]
- Nishihara, H.; Dekker, E.E. Purification, substrate specificity and binding, β-decarboxylase activity, and other properties of Escherichia coli 2-keto-4-hydroxyglutarate aldolase. J. Biol. Chem. 1972, 247, 5079–5087. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Noh, M.H.; Lim, H.G.; Kang, C.W.; Im, D.K.; Oh, M.K.; Jung, G.Y. Precise tuning of the glyoxylate cycle in Escherichia coli for efficient tyrosine production from acetate. Microb. Cell Fact. 2019, 18, 57. [Google Scholar] [CrossRef]
- Jing, P.; Cao, X.; Lu, X.; Zong, H.; Zhuge, B. Modification of an engineered Escherichia coli by a combined strategy of deleting branch pathway, fine-tuning xylose isomerase expression, and substituting decarboxylase to improve 1,2,4-butanetriol production. J. Biosci. Bioeng. 2018, 126, 547–552. [Google Scholar] [CrossRef]
- Pinhal, S.; Ropers, D.; Geiselmann, J.; de Jong, H. Acetate Metabolism and the Inhibition of Bacterial Growth by Acetate. J. Bacteriol. 2019, 201, e00147-19. [Google Scholar] [CrossRef]
- Deng, Y.; Mao, Y.; Zhang, X. Metabolic engineering of E. coli for efficient production of glycolic acid from glucose. Biochem. Eng. J. 2015, 103, 256–262. [Google Scholar] [CrossRef]
- Renilla, S.; Bernal, V.; Fuhrer, T.; Castaño-Cerezo, S.; Pastor, J.M.; Iborra, J.L.; Sauer, U.; Cánovas, M. Acetate scavenging activity in Escherichia coli: Interplay of acetyl-CoA synthetase and the PEP-glyoxylate cycle in chemostat cultures. Appl. Microbiol. Biotechnol. 2012, 93, 2109–2124. [Google Scholar] [CrossRef]
- Hua, Q.; Yang, C.; Baba, T.; Mori, H.; Shimizu, K. Responses of the central metabolism in Escherichia coli to phosphoglucose isomerase and glucose-6-phosphate dehydrogenase knockouts. J. Bacteriol. 2003, 185, 7053–7067. [Google Scholar] [CrossRef]
- Takahashi-Íñiguez, T.; Aburto-Rodríguez, N.; Vilchis-González, A.L.; Flores, M.E. Function, kinetic properties, crystallization, and regulation of microbial malate dehydrogenase. J. Zhejiang Univ. Sci. B 2016, 17, 247–261. [Google Scholar] [CrossRef]
- Kováč, L.; Berta, F.; Pšenák, M.; Slezáriková, V. Metabolic differences between Escherichia coli cultures growing aerobically and anaerobically in the presence of fluoroacetate. Folia Microbiol. 1966, 11, 263–270. [Google Scholar]
- Riesenberg, D.; Schulz, V.; Knorre, W.A.; Pohl, H.D.; Korz, D.; Sanders, E.A.; Roß, A.; Deckwer, W.D. High cell density cultivation of Escherichia coli at controlled specific growth rate. J. Biotech. 1991, 20, 17–27. [Google Scholar]
- Causey, T.B.; Shanmugam, K.T.; Yomano, L.P.; Ingram, L.O. Engineering Escherichia coli for efficient conversion of glucose to pyruvate. Proc. Natl. Acad. Sci. USA 2004, 101, 2235–2240. [Google Scholar] [PubMed]
- Lin, H.; Castro, N.M.; Bennett, G.N.; San, K.Y. Acetyl-CoA synthetase overexpression in Escherichia coli demonstrates more efficient acetate assimilation and lower acetate accumulation: A potential tool in metabolic engineering. Appl. Microbiol. Biotechnol. 2006, 71, 870–874. [Google Scholar] [PubMed]
- Negrete, A.; Majdalani, N.; Phue, J.N.; Shiloach, J. Reducing acetate excretion from E. coli K-12 by over-expressing the small RNA SgrS. New Biotechnol. 2013, 30, 269–273. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Description | Reference |
|---|---|---|
| Strains | ||
| Escherichia coli BW25113 | NBRC | |
| E. coli EG_1 | BW25113 ΔaceB | This work |
| E. coli EG_2 | BW25113 ΔaceB ΔglcB | This work |
| E. coli EG_3 | BW25113 ΔaceB ΔglcB Δgcl | This work |
| E. coli EG_4 | BW25113 ΔaceB ΔglcB Δgcl ΔycdW | This work |
| E. coli EG_5 | BW25113/pSR13 | This work |
| E. coli EG_6 | BW25113 ΔaceB ΔglcB Δgcl ΔycdW/pSR13 | This work |
| E. coli EG_7 | BW25113 ΔaceB ΔglcB Δgcl ΔycdW/pSR13/pLB6 | This work |
| Corynebacterium glutamicum NBRC 12168 | NBRC | |
| Plasmids | ||
| pGV3 | Expression vector | [6] |
| pTV118N | Expression vector | Takara Bio, Japan. |
| pSR13 | pGV3 harboring gltA and aceA, ΔoprI’ | This work |
| pLB6 | pTV118N harboring pyc | This work |
| pKD46 | λ-red recombinase expressing helper plasmid | [30] |
| Strains and Conditions | Highest Glyoxylate Titers (g/L) | Specific Productivities (g/g-Cell) | Other Products | |||
|---|---|---|---|---|---|---|
| Acetate (g/L) | Formate (g/L) | Lactate (g/L) | Cell Biomass (g/L) | |||
| BW25113, M9 medium | 0.01 ± 0.00 | 0.01 ± 0.00 | 1.98 ± 0.09 | 0.20 ± 0.04 | 0.01 ± 0.00 | 1.03 ± 0.02 |
| EG_1, M9 medium | 0.52 ± 0.07 | 1.01 ± 0.08 | 1.82 ± 0.04 | 0.09 ± 0.00 | 0 | 0.50 ± 0.03 |
| EG_2, M9 medium | 0.62 ± 0.12 | 1.12 ± 0.12 | 2.53 ± 0.18 | 0.09 ± 0.01 | 0.03 ± 0.00 | 0.54 ± 0.05 |
| EG_3, M9 medium | 0.71 ± 0.05 | 1.18 ± 0.06 | 2.99 ± 0.04 | 0.07 ± 0.00 | 0 | 0.60 ± 0.01 |
| EG_4, M9 medium | 0.93 ± 0.17 | 1.92 ± 0.32 | 2.09 ± 0.83 | 0.04 ± 0.02 | 0.01 ± 0.01 | 0.49 ± 0.03 |
| EG_5, M9 medium | 0.12 ± 0.00 | 0.14 ± 0.00 | 2.63 ± 0.02 | 0.03 ± 0.00 | 0.38 ± 0.00 | 0.87 ± 0.01 |
| EG_6, M9 medium | 1.15 ± 0.02 | 2.88 ± 0.02 | 1.83 ± 0.04 | 0.05 ± 0.00 | 0 | 0.40 ± 0.01 |
| EG_7, M9 medium | 1.30 ± 0.11 | 4.90 ± 0.57 | 2.45 ± 0.39 | 0.04 ± 0.01 | 0 | 0.28 ± 0.06 |
| EG_7, M9 medium, yeast extract | 2.42 ± 0.00 | 4.22 ± 0.09 | 1.72 ± 0.04 | 0.05 ± 0.01 | 0 | 0.57 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, B.H.D.; Nishiyama, M.; Sato, R.; Tanaka, T.; Ohara, H.; Aso, Y. Production of Glyoxylate from Glucose in Engineered Escherichia coli. Fermentation 2023, 9, 534. https://doi.org/10.3390/fermentation9060534
Long BHD, Nishiyama M, Sato R, Tanaka T, Ohara H, Aso Y. Production of Glyoxylate from Glucose in Engineered Escherichia coli. Fermentation. 2023; 9(6):534. https://doi.org/10.3390/fermentation9060534
Chicago/Turabian StyleLong, Bui Hoang Dang, Masahiro Nishiyama, Rintaro Sato, Tomonari Tanaka, Hitomi Ohara, and Yuji Aso. 2023. "Production of Glyoxylate from Glucose in Engineered Escherichia coli" Fermentation 9, no. 6: 534. https://doi.org/10.3390/fermentation9060534
APA StyleLong, B. H. D., Nishiyama, M., Sato, R., Tanaka, T., Ohara, H., & Aso, Y. (2023). Production of Glyoxylate from Glucose in Engineered Escherichia coli. Fermentation, 9(6), 534. https://doi.org/10.3390/fermentation9060534