Abstract
Pepsinogen A (PGA) plays an important role in the treatment of human gastrointestinal diseases. At present, PGA is mainly extracted from pig stomach, so its source is very limited and its price is very expensive. Production of PGA by microbial fermentation using an engineered strain with high PGA yield would be an ideal solution. This paper presents a new system for the high-level expression of PGA from Homo sapiens (hPGA) in Aspergillus niger. The hPGA5 gene codon was optimized according to the codon bias of A. niger and then connected to a strong promoter and signal peptide to construct an hPGA5 expression cassette. An ingenious multi-copy knock-in expression strategy mediated by the CRISPR/Cas9 tool was used to improve the production of hPGA in A. niger. By optimizing the copy number and integration sites of the hPGA5 gene, an engineering strain with a high yield of hPGA was constructed. After shake-flask fermentation for 7 d, the enzyme activity of recombinant hPGA reached 542.3 U/mL, which is the highest known activity. This lays a foundation for the production of hPGA by microbial fermentation.
1. Introduction
Pepsin (EC 3.4.23.1) is an aspartic endopeptidase, belonging to peptidase family A [1], which can catalyze the hydrolysis of a variety of proteins under acidic conditions. Pepsinogen (PG), the precursor of pepsin, is transformed into active pepsin by hydrochloric acid or active pepsin in the gastric cavity [2]. PG is widely used and plays an important role in the diagnosis and treatment of human gastrointestinal diseases [3,4] in addition to its use in normal digestion of foods. PG in human gastric mucosa can be divided into two major groups, pepsinogen I (PGA, PGI) and pepsinogen II (PGC, PGII) [5], and PGA is predominant in most mammals [6,7,8]. PGA from Homo sapiens (hPGA) is a polypeptide chain of 388 amino acids, including a 15-amino acid signal peptide, 44 amino acids longer than pepsin at the N-terminus. In terms of drug treatment, because of some people being allergic to porcine pepsinogen, hPGA is safer. At present, PGA is mainly extracted from pig stomach, so its source is very limited and the price is very expensive. Production of PGA by microbial fermentation using engineered strains would be ideal. To date, studies on PGA have mainly focused on the expression of porcine pepsinogen A in Escherichia coli and yeast. For example, in 1996, Takana et al. [9] successfully expressed porcine pepsin in Escherichia coli and analyzed its enzymatic properties. Although active porcine pepsin was obtained, the expression level was also very low, only 9 mg/L. In 2002, Mark et al. [10] first expressed porcine pepsin in Pichia pastoris, and the expression amount was only 30 mg/L. In 2006, Malik et al. [11] fused and expressed recombinant hPGA in Escherichia coli, but the expression amount was only 100 μg/L. Therefore, we need to improve expression level of hPGA for its application.
Aspergillus niger is a microorganism generally recognized as safe (GRAS) and has been widely used in the food industry [12]. Meanwhile, with many advantageous properties, including an exceptional capacity for protein secretion, the presence of post-translational modifications (protein glycosylation, proteolytic cleavage and formation of multiple disulfide bonds), and the ability to grow on fairly simple and inexpensive substrates, A. niger has become the preferred industrial host strain for the production of a variety of homologous and heterologous proteins [13,14,15,16,17,18]. In recent years, the genome-editing system CRISPR/Cas9 has been successfully applied to a variety of filamentous fungi, including A. niger, for gene knockout, knock-in and genetic alteration [19,20,21,22]. Our lab has also established an efficient marker-free CRISPR/Cas9 genome editing technique which could simultaneously edit several genes in A. niger [23]. In this CRISPR/Cas9 system, a 5SrRNA promoter was used to initiate highly efficient expression of sgRNA and a co-expression plasmid of sgRNA and Cas9 was constructed based on the initial fragment of AMA1 replication [23]. When several co-expression plasmids of sgRNA and Cas9 and donor DNA fragments were co-transformed, strains with different copies at different loci could be achieved easily using this CRISPR/Cas9 system [23].
In this study, we hope to improve hPGA expression by integrating the hPGA5 gene into the genome of A. niger 41683 using a multi-copy gene knock-in strategy. Using the CRISPR/Cas9 technique, nineteen strains with different hPGA5 copies at 10 loci were constructed and a recombinant strain with a high yield of hPGA was obtained. After shake-flask fermentation for 7 days, the enzyme activity of recombinant hPGA reached 542.3 U/mL, which is the highest known activity. This lays a foundation for the production of hPGA.
2. Materials and Methods
2.1. Strains and Culture Conditions
A. niger 41683, a high glucoamylase-producing strain, was used as the host for expressing hPGA. A. niger 41683 was cultured in CD (Czapek–Dox) medium or in DPY (dextrose peptone yeast extract) at pH 5.5 and 30 °C. The fermentation of positive transformants was cultured in malt extract medium at 30 °C and 200 r/min. E. coli DH5α was cultured in Luria–Bertani (LB) medium with 100 μg/μL ampicillin at 37 °C and was used for construction and propagation of recombinant plasmid.
2.2. Construction of Recombinant Plasmid
First, the hPGA5 expression cassette, consisting of strong promoter Pglucan703, SP (signal peptide), hPGA5 and Tglucan693, was constructed by overlap PCR. Pglucan703, a stronger promoter than PglaA (identified by our lab), SP and Tglucan693 were from the glucoamylase gene of Talaromyces emersonii and were cloned by PCR. The nucleotide sequence of hPGA5 was optimized according to the codon usage bias of A. niger and was obtained via gene synthesis by the GeneScript company (Nanjing, China). The free expression plasmid pFC332-hPGA5 was constructed by cloning the hPGA5 expression cassette into the Bgl II and BamH I sites of pFC332. In this study, we selected neutral amylase gene amyA (An05g02100, An12g06930), glucoamylase gene glaA (An03g06550) and seven α-glucosidase genes (agd) as the knock-in targets, named agdA (An04g06920), agdB (An01g10930), agdC (An02g13240), agdD (An13g03710), agdE (An09g05880), agdF (An18g05620) and agdG (An07g00350). The sgRNA expression cassette is composed of promoter 5SrRNA, 20 bp target, gRNA scaffold and T6 terminator. Target sequences were designed as fully complementary paired 20 bp upstream and downstream primers with an additional four bases of CACC at the 5′ end of the upstream primer and four additional bases of CAAA at the 5′ end of the downstream primer (Table S1). The primers target-sgRNA-F and target-sgRNA-R anneal to double-stranded DNA fragments containing sticky ends (CACC- and -CAAA). The plasmid pUC57-target-sgRNA (Figure 1B) was constructed by ligating the DNA double-stranded fragment containing the sticky end to the digested plasmid pUC57-sgRNA by Bbs I [23]. The plasmid pUC57-2sgRNA was constructed by ligating sgRNA cassette1 digested by Xho I/BamH I to pUC57-sgRNA cassette2 digested by Xho I and Bgl II (Figure 1B). The plasmid pFC332-2sgRNA (Figure 1B) was constructed by ligating 2sgRNA cassette digested by Bgl II/Nhe I to the digested plasmid pFC332-prtT-sgRNA-Nhe I by Bgl II/Nhe I [23]. For the construction of donor DNA (dDNA) fragments, the plasmid pUC57-hPGA5 (Figure 1A) was first constructed by cloning hPGA5 cassette in between the Xho I/Pac I sites of plasmid pUC57-sgRNA. Then, the up-arm and the down-arm of the target genes were amplified from the A. niger 41683 genome by PCR and inserted, respectively, between the Xho I/Bgl II and BamH I/Hind III sites of plasmid pUC57-hPGA5 for constructing the plasmid pUC57-up arm-hPGA5-down arm (Figure 1A). The dDNAs containing the hPGA5 cassette were obtained by digesting plasmid pUC57-up arm-hPGA5-down arm with Xho I/Hind III.
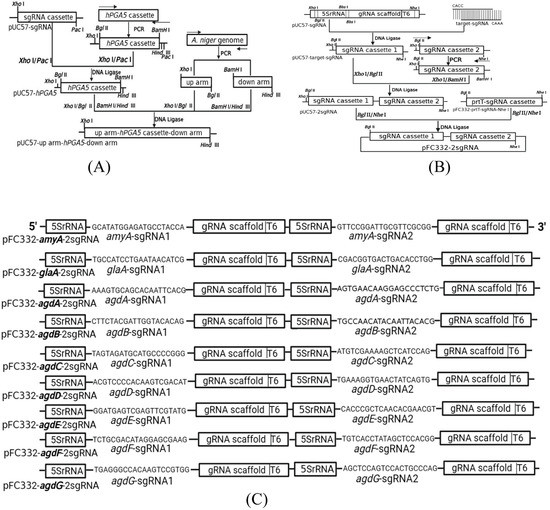
Figure 1.
Construction of recombinant hPGA expression plasmid and knockout plasmid. (A) Schematic of plasmid pUC57-up arm-hPGA5-down arm construction. (B) Schematic of knockout plasmid pFC332-2sgRNA construction. (C) Schematic diagram of the sequence containing the sgRNA expression cassette in the knockout plasmid pFC332-2sgRNA.
2.3. Transformation and Expression
A. niger 41683 was cultivated for 48 h at 30 °C in DPY medium for the preparation of protoplasts. The hPGA expression plasmid pFC332-hPGA5 was transformed into A. niger by using the protoplast transformation method [24]. The positive transformants were identified on CD plates by PCR and verified by DNA sequencing.
To integrate the hPGA5 cassette into the genome of A. niger, dDNAs containing the hPGA5 cassette and plasmid pFC332-2sgRNA were co-transformed into A. niger 41683 by PEG-mediated protoplast transformation method [24]. First, two plasmids (pFC332-amyA-2sgRNA and pFC332-glaA-2sgRNA) and three dDNAs (hPGA5 cassette-amyA (An05), hPGA5 cassette-amyA (An12) and hPGA5 cassette-glaA) were co-transformed into A. niger in order to integrate the hPGA5 cassette into amyA and glaA sites. After protoplast transformation was completed, recombinant hPGA strains with different integration sites and copy numbers were generated by screening and verifying the transformants one by one. Similarly, four plasmids pFC332-agdA/D/E/F-2sgRNA and four dDNAs, including hPGA5 cassette-agdA, hPGA5 cassette-agdD, hPGA5 cassette-agdE and hPGA5 cassette-agdF, were co-transformed into A. niger in order to integrate the hPGA5 cassette into agdA/D/E/F sites. In the same way, three plasmids pFC332-agdB/C/G-2sgRNA and three dDNAs, including hPGA5 cassette-agdB, hPGA5 cassette-agdC and hPGA5 cassette-agdG, were co-transformed into A. niger in order to integrate the hPGA5 cassette into agdB/C/G sites.
The identified transformants were inoculated into 50 mL of fermentation medium (malt extract medium) in 250 mL shaking flasks at 200 r/min and 30 °C. The incubation period was 8 d, samples were taken at 24 h intervals, and extracellular hPGA activity was measured after centrifugation and filtration of the fermentation broth.
2.4. Assay of Pepsin Activity
The activity of recombinant hPGA were determined using haemoglobin as a substrate using methods previously described by André Brodkorb [25] with some modifications. Briefly, 2 g of hemoglobin was dissolved in 80 mL of purified water, the pH was adjusted to 2 with 300 mM HCl, and the volume was completed to 100 mL to obtain a 2% w/v hemoglobin solution at pH 2. The fermentation supernatant was added to the reaction with 2% substrate hemoglobin (pH = 2) at 37 °C, and the reaction was terminated with 5% trichloroacetic acid (TCA). The specific methods for measuring enzyme activity determination were divided into experimental group and control group. First, 300 µL of 2% hemoglobin was added to a 1.5 mL centrifugal tube in the experimental group and control group and preheated at 37 °C for 5 min. Then, 600 µL of 5% TCA was added to the control group, followed by 100 µL of fermentation supernatant, whereas only 100 µL of fermentation supernatant was added to the experimental group. The control group and the experimental group reacted at 37 °C for 10 min, and 600 µL 5% TCA was added to the experimental group to terminate the reaction. At the end of the reaction, the supernatant was centrifuged at 12,000 r/min for 5 min at room temperature, and the absorbance was measured at OD280 nm. One unit of enzyme activity was defined as 0.001 of the absorbance difference, ∆A280 nm, produced by TCA-soluble products per minute at pH 2.0 and 37 °C. The enzyme activity of the recombinant hPGA was calculated as the measured activity of the recombinant hPGA strain minus that of the wild-type strain, because the fermentation supernatant of the control wild-type strain could also react with the substrate hemoglobin to form TCA-soluble products.
3. Results
3.1. Free Expression of hPGA
To express hPGA in A. niger 41683, we constructed the hPGA expression plasmid pFC332-hPGA5. The hPGA5 contained 373 amino acids mature peptide and a 15-amino acid signal peptide predicted by SignalP-5.0 Server. In order to allow the extracellular secretion of recombinant hPGA, the native signal peptide of the hPGA5 gene was replaced by the signal peptide of the glaA gene from T. emersonii. The hPGA expression plasmid pFC332-hPGA5 was transformed into A. niger 41683 by the protoplast transformation method [24]. After 5 days of incubation, monoclonal transformants were selected and verified by PCR. After PCR verification, the free expression strains Fr-hPGA were constructed.
The validated Fr-hPGA transformants were subjected to shake-flask fermentation and sampled at 24 h intervals over an 8 d period. The enzyme activity assay of the fermentation supernatant showed that, with the extension of fermentation time, the enzyme activity of Fr-hPGA transformants gradually increased, reaching a maximum of 109.4 U/mL after 6 d fermentation (Figure 2A). There was a faint protein band (shown as a red arrow) at a position of approximately 42.97 kDa that was absent in the host (Figure 2B), indicating that hPGA has been successfully expressed in A. niger 41683, but the expression level is low. Figure 2B shows that the band of hPGA was more obvious on day 8 than on days 4–7, but the enzyme activity decreased on day 8. The following two reasons could be explain this observation. First, the amount of protein may not be proportional to the enzyme activity measured. For example, in 2020, Dong et al. [16] measured the enzyme activity of recombinant Trehalase in Du-Trem transformants, and the enzyme activity reached the highest on day 7, but the SDS-PAGE results showed that the protein band on day 8was more obvious than that on day 7. Secondly, other by-products in the fermentation supernatant will become more and more complex as the fermentation time is prolonged, which may affect the enzyme activity.
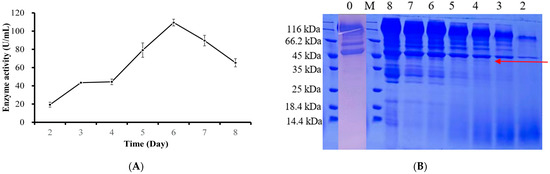
Figure 2.
Free expression of hPGA in A. niger 41683. (A) Enzyme activity assay. Determination of hPGA enzyme activity at different fermentation times. Each plot represents the mean value with standard deviations. (B) The SDS-PAGE analysis of secreted hPGA proteins. Lanes 2–8, fermentation supernatants collected from 2 to 8 days; lane 0: the wild type host at day 6. M: protein molecular weight marker. The red arrow represents the protein band of recombinant hPGA.
3.2. Construction and Screening of Recombinant hPGA Strains
To further improve the expression of hPGA in A. niger 41683, different recombinant hPGA strains with different hPGA5 copies sited at different loci were constructed using the CRISPR/Cas9 tool. To integrate the hPGA5 cassette into the genome of A. niger, dDNAs containing the hPGA5 cassette and plasmid pFC332-2sgRNA were co-transformed into A. niger 41683. After 6 days of incubation, the transformants were selected and subcultured twice on CD plates. Then, we detected whether the hPGA5 expression cassette was integrated into the A. niger 41683 genome by PCR with primers F-hPGA160/R-hPGA1077 (Table S1), the results showed that almost all transformants contained the hPGA5 cassette. To confirm whether the hPGA5 cassettes were precisely located at the site of amyA, glaA, agdA, agdB, agdC, agdD, agdE, agdF or agdG, PCR using primers of the up-arm and the down-arm of the target genes (Table S1) was performed, the results are shown in Figure 3. All bands were the expected size, showing that the hPGA5 cassettes were precisely located at the sites of the genes of interest and nineteen recombinant hPGA strains with different copy numbers at different loci were generated, of which, eight strains harbored one hPGA5 copy, eight strains harbored two hPGA5 copies, two strains harbored three hPGA5 copies and only one strain harbored four hPGA5 copies. All the generated recombinant hPGA5 strains of the different genotypes are shown in Table 1.

Figure 3.
Screening and validation of recombinant hPGA strains by PCR. (A) PCR location of hPGA5 expression cassette in 6 transformants (An_1-1~An_1-6). (B) PCR location of hPGA5 expression cassette in 9 transformants (An_2-1~An_2-9). (C) PCR location of hPGA5 expression cassette in 4 transformants (An_2-10~An_2-13). M: 1 kb DNA ladder.

Table 1.
Recombinant hPGA strains of different genotypes.
3.3. Expression of Recombinant hPGA at Different Integration Sites
In order to detect the expression of hPGA at different sites on A. niger 41683, eight recombinant hPGA strains with different integration sites were selected, which were An_1-4, An_1-5, An_1-6, An_2-1, An_2-2, An_2-5, An_2-10 and An_2-11. Eight recombinant hPGA strains were inoculated in fermentation media and cultivated at 30 °C and 200 r/min. Fermentation supernatants were collected at 2–8 days. The enzyme activity of hPGA in these fermentation supernatants was measured, and the results are shown in Figure 4A. With the extension of fermentation time, enzyme activity of hPGA increased gradually, and generally reached the highest level on day 7. The enzyme activities of hPGA expressed in recombinant strains An_1-5, An_2-1, An_2-2 and An_2-10 were over 500 U/mL, which is much higher thanin other recombinant strains. According to the results of SDS-PAGE analysis of recombinant hPGA on the 7th day (Figure 5A–C), the protein bands (shown as a red arrow) of recombinant strains An_1-5, An_2-1, An_2-2 and An_2-10 were more obvious than those of other single-copy recombinant strains. The hPGA activity expressed in An_1-6 was the lowest, and only 106.9 U/mL at day 7, whereas hPGA activity expressed in An_2-2 was the highest, and reached 542.3 U/mL at day 7. SDS-PAGE of recombinant hPGA (Figure 5A,C) also showed that the protein band of recombinant strain An_1-6 was the faintest and the protein band of An_2-2 was the most obvious compared with other single-copy recombinant strains. In recombinant hPGA strains An_1-5, An_2-1, An_2-2 and An_2-10, hPGA cassettes were, respectively, integrated at amyA (12), agdA, agdF and agdB loci, showing that these sites were favorable integration sites for single-copy hPGA expression. In contrast, the glaA site in strain An_1-6 was an inappropriate integration site because hPGA expression was the lowest.
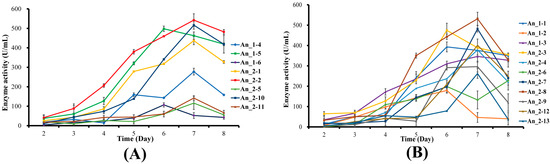
Figure 4.
Expression of recombinant hPGA. (A) Expression of recombinant hPGA at different integration sites. Enzyme activity analysis of 8 transformants, 2–8 days. Each plot represents the mean value with standard deviations. (B) Expression of recombinant hPGA with different copy numbers. Enzyme activity analysis of 11 transformants, 2–8 days. Each plot represents the mean value with standard deviations.

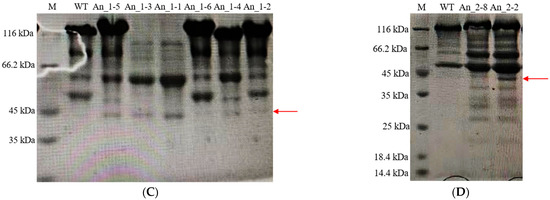
Figure 5.
SDS-PAGE analysis of recombinant hPGA on the 7th day. (A) Transformants An_2-1~An_2-8; (B) Transformants An_2-9~An_2-13; (C) Transformants An_1-1~An_1-6. M: protein molecular weight marker; WT: wild-type strain. D) SDS-PAGE analysis of hPGA proteins secreted by 2 transformants (An_2-2, An_2-8) on the 7th day. M: protein molecular weight marker; WT: wild-type strain on the 7th day. The red arrows represent the protein band of recombinant hPGA.
3.4. Expression of Recombinant hPGA with Different Copy Numbers
To detect expression of recombinant hPGA with different copy numbers, 11 recombinant hPGA strains with 2~4 hPGA5 copies, which were, respectively, An_1-1 (2 copies), An_1-2 (2 copies), An_1-3 (3 copies), An_2-3 (2 copies), An_2-4 (4 copies), An_2-6 (2 copies), An_2-7 (2 copies), An_2-8 (2 copies), An_2-9 (3 copies), An_2-12 (2 copies) and An_2-13 (2 copies), were subjected to shake-flask fermentation at 30 °C and 200 r/min and sampled at 24 h intervals over an 8-day period. The enzyme activity of hPGA in these fermentation supernatants was measured, and the results are shown in Figure 4B. With the extension of fermentation time, enzyme activity of hPGA increased gradually, and generally reached its highest at 6–7 d. Strain An_2-4 harbored 4 hPGA5 copies located at agdA, agdD, agdE and agdF sites; hPGA activity was only 378.9 U/mL, which was lower than that in strains An_1-1, An_2-3, An_2-7, An_2-8 and An_2-12, which contained 2 hPGA5 copies, but was higher than that in strains An_1-3 (346.5 U/mL) and An_2-9 (296.2 U/mL), which contained 3 hPGA5 copies. There was a protein band (shown as a red arrow) at a position of approximately 42.97 kDa, which was not present in the wildtype strain (Figure 5A–C), indicating that hPGA has been successfully expressed in multi-copy recombinant hPGA strains. The hPGA activity expressed in strain An_1-2 was the lowest and only reached 179.0 U/mL on day 7, whereas the hPGA activity expressed in strain An_2-8 was the highest and reached 531.3 U/mL on day 7. SDS-PAGE analysis of recombinant hPGA on the 7th day (Figure 5A,C) showed that the protein band of recombinant strain An_1-2 was the faintest and the protein band of An_2-8 was more obvious compared with other multi-copy recombinant strains. Strain An_2-8 contained two hPGA5 copies at the agdE and agdF sites, and An_1-2 contained two hPGA5 copies at the glaA and amyA (An12) sites. These results showed that the integration sites of hPGA5 were the most important for expression of hPGA, and agdF and agdE sites were the preferred integration sites for multiple-copy hPGA expression.
To compare the difference in protein production between the more active recombinant strains, we selected two strains with higher activities, An_2-2 in 8 single-copy hPGA recombinant strains and An_2-8 in 11 multi-copy hPGA recombinant strains, to visually show the difference in protein production. SDS-PAGE analysis was performed on strains An_2-2 and An_2-8. The results (Figure 5D) showed that the protein bands of strain An_2-2 were more obvious than those of strain An_2-8.
4. Discussion
For the recombinant expression of heterologous proteins, in addition to the selection of appropriate promoters, the chromosomal integration site of heterologous genes is generally considered to be an important factor affecting gene expression [26]. The expression level of heterologous genes varies in different transformants at different chromosomal locations of target genes [27,28]. In Trichoderma reesei, the exocellobiohydrolase I encoding gene (cbh1) locus is the preferred integration site, where CBHI protein is the major abundant extracellular protein, resulting in high expression of homologous or heterologous genes. The neutral amylase and the glucoamylase were abundantly secreted in A. niger 41683. In addition, α-glucosidase has a dual role in the catalytic reaction of sugar hydrolysis and transglucoside. Owing to the transglycosidic activity of α-glucosidase, it can form non-fermentable oligosaccharides, e.g., isomaltose, by transfering glucose groups to substrate molecules such as maltose and glucose. Therefore, in this study, the amyA, glaA and seven α-glucosidase gene (agdA, agdB, agdC, agdD, agdE, agdF and agdG) sites were selected as the integration sites for the hPGA5 cassette. The results showed that hPGA activity improved when the hPGA5 cassette was integrated at amyA, agdA, agdF and agdB loci, compared with free expression. This showed that the integration sites are very important for hPGA expression and that amyA, agdA, agdF and agdB loci are favorable integration sites (Figure 4).
In addition to selected promoters and chromosomal integration sites, gene expression is also influenced by gene copy number [29]. To integrate hPGA5 at different loci of the A. niger genome with different hPGA5 copy numbers, an effective marker-free CRISPR/Cas9 genome editing technique established by our lab, which could simultaneously edit several genes [23], was applied. In this study, the co-expression plasmid pFC332-2sgRNA contained two sgRNA expression cassettes at different positions on the target gene site in order to improve integration efficiency. Through three co-transformations of pFC332-2sgRNAs and dDNAs, 19 recombinant hPGA strains with different hPGA5 copies at 10 loci were obtained, of which, 8 recombinant hPGA strains contained one hPGA5 copy, 8 recombinant hPGA strains contained two hPGA5 copies, 2 recombinant hPGA strains contained three hPGA5 copies and 1 recombinant hPGA strain contained four hPGA5 copies (Table 1). The hPGA activities expressed in these strains were measured (Figure 4), and it was found that hPGA activity by integrated expression was significantly higher than that by free expression in A. niger 41683. In the strains with one hPGA5 copy, hPGA activity integrated at the agdF site (An_2-2) was the highest, reaching 542.3 U/mL. Secondly, when the hPGA5 gene was integrated at the agdB (An_2-10), amyA(12) (An_1-5) and agdA (An_2-1) sites, hPGA activities were 516.5 U/mL, 497.8 U/mL and 436.9 U/mL, respectively. hPGA activity integrated at the glaA site (An_1-6) was the lowest; it was only 106.9 U/mL. However, in A. niger 41683, the expression level of GlaA was the highest, the reason why hPGA activity integrated at the glaA site (An_1-6) was the lowest could be that the utilization of sugars was affected when the glaA gene was replaced by the hPGA5 gene. In the strains with two hPGA5 copies, hPGA activities expressed in An_2-8 (531.3 U/mL), An_2-7 (481.6 U/mL), An_2-3 (474.4 U/mL), An_2-12 (401.3 U/mL) and An_1-1 (393.3 U/mL) were higher than those expressed in the strains with three hPGA5 copies (An_1-3, 346.5 U/mL; An_2-9, 296.2 U/mL) and in the strains with four hPGA5 copies (An_2-4, 378.9 U/mL). These results indicate that both hPGA5 copy number and their integration site affected the expression of hPGA, and the influence of integration site on hPGA expression was the most important. As an explanation for the phenomenon that hPGA activities expressed in three and four-hPGA5-copy strains were not the highest, the ER-associated degradation (ERAD) and unfolded protein response (UPR) associated with protein overexpression may have limited the increase in hPGA expression [30,31]; this needs further study.
5. Conclusions
We presented a new expression system for the high-level expression of PGA from Homo sapiens (hPGA) in Aspergillus niger. Using a multi-copy knock-in expression strategy mediated by the CRISPR/Cas9 system, nineteen strains with different hPGA5 copies at 10 loci were constructed. It was found that hPGA activity by integrated expression was significantly higher than that by free expression in A. niger 41683, and the integrated expression of hPGA was greatly affected by the copy number and integration site of the hPGA5 gene; the influence of integration site on hPGA expression was the most important. By optimizing the copy number and integration sites of hPGA5, a recombinant strain with a high yield of hPGA was obtained. After shake-flask fermentation for 7 days, the highest enzyme activity of recombinant hPGA reached 542.3 U/mL. This lays a foundation for the production of hPGA by microbial fermentation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation9060538/s1, Table S1: Primers used in this study.
Author Contributions
Experiment method design, M.L., F.L. and J.C.; experiment performance, J.C., L.G., B.C. and Y.S.; writing—original draft preparation, M.L., F.L. and J.C.; writing—review and editing, J.C., L.G., B.C. and Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Natural Science Foundation of China (No. 32072161, No. 22278321) and National Key Research and Development Program of China (No. 2021YFC2101800, No. 2021YFC2100400).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are available from the corresponding author upon request.
Acknowledgments
The authors wish to take the opportunity to thank Yuyu Shen for her helpful input during discussions. The authors would like to thank Zhongxiu Chen for his help with experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Herriott, R.M. Isolation, crystallization, and properties of swine pepsinogen. J. Gen. Physiol. 1938, 21, 501–540. [Google Scholar] [CrossRef] [PubMed]
- Plebani, M.; Szabo, O.S. Pepsinogens in Health and Disease. Crit. Rev. Clin. Lab. Sci. 1993, 30, 273–328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-M.; Li, J.-X.; Zhang, G.-Y.; Li, X.-H.; Gu, H. The value of serum pepsinogen levels for the diagnosis of gastric diseases in Chinese Han people in midsouth China. BMC Gastroenterol. 2014, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Furihata, C. Human gastric cancer risk screening: From rat pepsinogen studies to the ABC method. Proc. Jpn. Acad. Ser. B 2021, 97, 462–478. [Google Scholar] [CrossRef] [PubMed]
- Gritti, I.; Banfi, G.; Roi, G. Pepsinogens: Physiology, pharmacology pathophysiology and exercise. Pharmacol. Res. 2000, 41, 265–281. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Narita, Y.; Oda, S.-I.; Moriyama, A.; Takenaka, O.; Kageyama, T. Purification and characterization of goat pepsinogens and pepsins. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1999, 122, 453–460. [Google Scholar] [CrossRef]
- Samloff, I.M. Slow moving protease and the seven pepsinogens. Electrophoretic demontration of the existence of eight proteolytic fractions in human gastric mucosa. Gastroenterology 1969, 57, 659–669. [Google Scholar] [CrossRef]
- Narita, Y.; Oda, S.-I.; Moriyama, A.; Takenaka, O.; Kageyama, T. Pepsinogens and Pepsins from House Musk Shrew, Suncus murinus: Purification, Characterization, Determination of the Amino-Acid Sequences of the Activation Segments, and Analysis of Proteolytic Specificities. J. Biochem. 1997, 121, 1010–1017. [Google Scholar] [CrossRef]
- Tanaka, T.; Yada, R.Y. Expression of soluble cloned porcine pepsinogen A in Escherichia coli. Biochem. J. 1996, 315 Pt 2, 443–446. [Google Scholar] [CrossRef]
- Yoshimasu, A.M.; Ahn, J.-K.; Tanaka, T.; Yada, R.Y. Soluble expression and purification of porcine pepsinogen from Pichia pastoris. Protein Expr. Purif. 2002, 25, 229–236. [Google Scholar] [CrossRef]
- Malik, A.; Rudolph, R.; Söhling, B. A novel fusion protein system for the production of native human pepsinogen in the bacterial periplasm. Protein Expr. Purif. 2006, 47, 662–671. [Google Scholar] [CrossRef]
- Fleißner, A.; Dersch, P.; Fleissner, A. Expression and export: Recombinant protein production systems for Aspergillus. Appl. Microbiol. Biotechnol. 2010, 87, 1255–1270. [Google Scholar] [CrossRef]
- He, Y.; Pan, L.; Wang, B. Efficient Over-expression and Application of High-performance Pectin Lyase by Screening Aspergillus niger Pectin Lyase Gene Family. Biotechnol. Bioprocess Eng. 2018, 23, 662–669. [Google Scholar] [CrossRef]
- Mekmouche, Y.; Zhou, S.; Cusano, A.M.; Record, E.; Lomascolo, A.; Robert, V.; Simaan, A.J.; Rousselot-Pailley, P.; Ullah, S.; Chaspoul, F.; et al. Gram-scale production of a basidiomycetous laccase in Aspergillus niger. J. Biosci. Bioeng. 2014, 117, 25–27. [Google Scholar] [CrossRef]
- Chen, X.; Wang, B.; Pan, L. Heterologous expression and characterization of Penicillium citrinum nuclease P1 in Aspergillus niger and its application in the production of nucleotides. Protein Expr. Purif. 2019, 156, 36–43. [Google Scholar] [CrossRef]
- Dong, L.; Yu, D.; Lin, X.; Wang, B.; Pan, L. Improving expression of thermostable trehalase from Myceliophthora sepedonium in Aspergillus niger mediated by the CRISPR/Cas9 tool and its purification, characterization. Protein Expr. Purif. 2020, 165, 105482. [Google Scholar] [CrossRef]
- Liu, F.; Wang, B.; Ye, Y.; Pan, L. High level expression and characterization of tannase tan7 using Aspergillus niger SH-2 with low-background endogenous secretory proteins as the host. Protein Expr. Purif. 2018, 144, 71–75. [Google Scholar] [CrossRef]
- Benghazi, L.; Record, E.; Suárez, A.; Gomez-Vidal, J.A.; Martínez, J.; de la Rubia, T. Production of the Phanerochaete flavido-alba laccase in Aspergillus niger for synthetic dyes decolorization and biotransformation. World J. Microbiol. Biotechnol. 2014, 30, 201–211. [Google Scholar] [CrossRef]
- Zheng, X.; Zheng, P.; Zhang, K.; Cairns, T.C.; Meyer, V.; Sun, J.; Ma, Y. 5S rRNA Promoter for Guide RNA Expression Enabled Highly Efficient CRISPR/Cas9 Genome Editing in Aspergillus niger. ACS Synth. Biol. 2019, 8, 1568–1574. [Google Scholar] [CrossRef]
- Shi, T.-Q.; Liu, G.-N.; Ji, R.-Y.; Shi, K.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. CRISPR/Cas9-based genome editing of the filamentous fungi: The state of the art. Appl. Microbiol. Biotechnol. 2017, 101, 7435–7443. [Google Scholar] [CrossRef]
- Yin, C.; Wang, B.; He, P.; Lin, Y.; Pan, L. Genomic analysis of the aconidial and high-performance protein producer, industrially relevant Aspergillus niger SH2 strain. Gene 2014, 541, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Nødvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chen, Z.; Chen, J.; Zhao, B.; Lu, J.; Gui, L.; Lu, F.; Li, M. An efficient marker-free genome editing method for Aspergillus niger. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2022, 38, 4744–4755. [Google Scholar] [CrossRef]
- Van Hartingsveldt, W.; Mattern, I.E.; Van Zeijl, C.M.J.; Pouwels, P.H.; van den Hondel, C.A. Development of a homologous transformation system for Aspergillus niger based on the pyrG gene. Mol. Genet. Genom. 1987, 206, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Qin, L.; Jiang, X.; Dong, Z.; Huang, J.; Chen, X. Identification of two integration sites in favor of transgene expression in Trichoderma reesei. Biotechnol. Biofuels 2018, 11, 142. [Google Scholar] [CrossRef]
- Qin, L.-N.; Cai, F.-R.; Dong, X.-R.; Huang, Z.-B.; Tao, Y.; Huang, J.; Dong, Z.-Y. Improved production of heterologous lipase in Trichoderma reesei by RNAi mediated gene silencing of an endogenic highly expressed gene. Bioresour. Technol. 2012, 109, 116–122. [Google Scholar] [CrossRef]
- Flagfeldt, D.B.; Siewers, V.; Huang, L.; Nielsen, J. Characterization of chromosomal integration sites for heterologous gene expression in Saccharomyces cerevisiae. Yeast 2009, 26, 545–551. [Google Scholar] [CrossRef]
- Scorer, C.A.; Clare, J.J.; McCombie, W.R.; Romanos, M.A.; Sreekrishna, K. Rapid Selection Using G418 of High Copy Number Transformants of Pichia pastoris for High–level Foreign Gene Expression. Nat. Biotechnol. 1994, 12, 181–184. [Google Scholar] [CrossRef]
- Heimel, K. Unfolded protein response in filamentous fungi—Implications in biotechnology. Appl. Microbiol. Biotechnol. 2015, 99, 121–132. [Google Scholar] [CrossRef]
- Nevalainen, K.H.; Te′O, V.S.J.; Bergquist, P.L. Heterologous protein expression in filamentous fungi. Trends Biotechnol. 2005, 23, 468–474. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).