
Antioxidant Capacity of Lactic Acid Bacteria and Yeasts from Xinjiang Traditional Fermented Dairy Products

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Microorganisms
2.3. Instruments and Equipment
2.4. Methods
2.4.1. Sample Preparation
2.4.2. Hydroxyl Radical Scavenging Activity
2.4.3. O2− Radical Scavenging Activity
2.4.4. DPPH Radical Scavenging Activity
2.4.5. ABTS+ Radical Scavenging Activity Assays
2.4.6. Lipid Peroxidation Inhibition Activity
2.4.7. Fe2+ Chelating Ability
2.4.8. Antioxidant Enzyme Activity
2.5. Statistical Analysis
3. Results
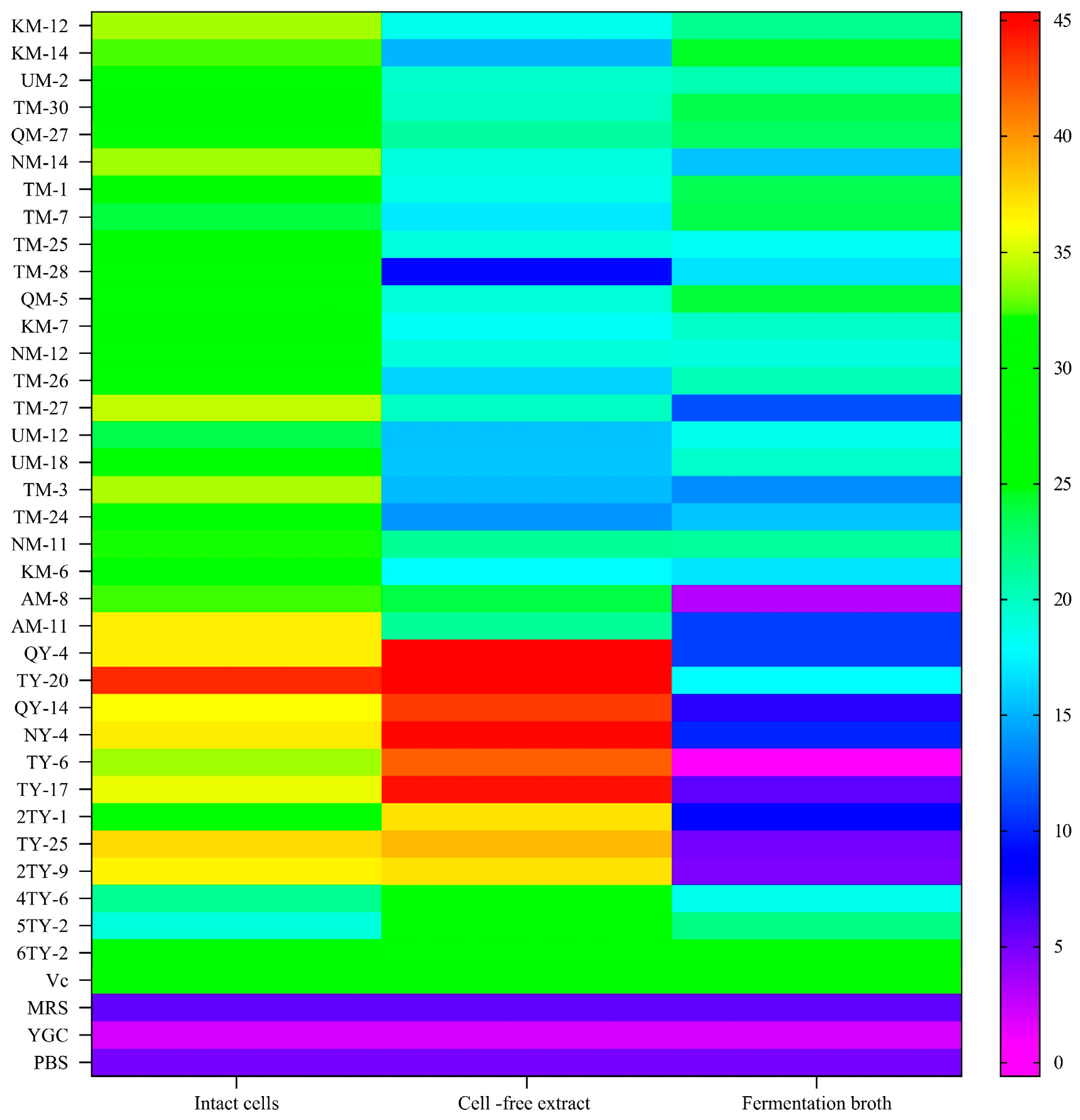
3.1. Scavenging of Hydroxyl Radicals
3.2. O2− Radical Scavenging Activity
3.3. DPPH Radical Scavenging Activity
3.4. ABTS+ Radical Scavenging Activity
3.5. Lipid Peroxidation Inhibition Activity
3.6. Fe2+ Chelating Ability
3.7. Antioxidant Enzyme Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Genus | Species | GenBank | Source |
|---|---|---|---|---|
| KM-12 | Lactobacillus | Lactobacillus gallinarum | MW931798.1 | Uruqmi yogurt pimple |
| KM-14 | Lactobacillus | Lactobacillus gallinarum | MW931799.1 | Uruqmi yogurt pimple |
| UM-2 | Lactobacillus | Lactobacillus gallinarum | MW931811.1 | Uruqmi cheese |
| TM-30 | Lactobacillus | Lactobacillus gallinarum | MW931793.1 | Tashkurgan County cheese |
| QM-27 | Enterococcus | Enterococcus durans | MW931809.1 | Tacheng cheese |
| NM-14 | Enterococcus | Enterococcus durans | MW931841.1 | Habahe County cheese |
| TM-1 | Enterococcus | Enterococcus durans | MW931780.1 | Tashkurgan County cheese |
| TM-7 | Enterococcus | Enterococcus durans | MW931782.1 | Tashkurgan County cheese |
| TM-25 | Enterococcus | Enterococcus durans | MW931789.1 | Tashkurgan County cheese |
| TM-28 | Enterococcus | Enterococcus durans | MW931792.1 | Tashkurgan County cheese |
| QM-5 | Lacticaseibacillus | Lacticaseibacillus paracasei | MW931815.1 | Tacheng cheese |
| KM-7 | Lacticaseibacillus | Lacticaseibacillus paracasei | MW931821.1 | Uruqmi yogurt pimple |
| NM-12 | Lacticaseibacillus | Lacticaseibacillus paracasei | MW931840.1 | Habahe County cheese |
| TM-26 | Lacticaseibacillus | Lacticaseibacillus paracasei | MW931790.1 | Tashkurgan County cheese |
| TM-27 | Lacticaseibacillus | Lacticaseibacillus paracasei | MW931791.1 | Tashkurgan County cheese |
| UM-12 | Enterococcus | Enterococcus faecium | MW931812.1 | Uruqmi cheese |
| UM-18 | Enterococcus | Enterococcus faecium | MW931813.1 | Uruqmi cheese |
| TM-3 | Enterococcus | Enterococcus faecium | MW931781.1 | Tashkurgan County cheese |
| TM-24 | Enterococcus | Enterococcus faecium | MW931788.1 | Tashkurgan County cheese |
| NM-11 | Enterococcus | Enterococcus faecium | MW931839.1 | Habahe County cheese |
| KM-6 | Levilactobacillus | Levilactobacillus brevis | MW931796.1 | Uruqmi yogurt pimple |
| AM-8 | Lacticaseibacillus | Lacticaseibacillus rhamnosus | MW931827.1 | Fuyun County cheese |
| AM-11 | Lacticaseibacillus | Lacticaseibacillus rhamnosus | MW931830.1 | Fuyun County cheese |
| QY-4 | Pichia | Pichia fermentans | GU373759.1 | Tacheng cheese |
| TY-20 | Pichia | Pichia fermentans | MZ314865 | Tashkurgan County cheese |
| QY-14 | Geotrichum | Geotrichum candidum | MN736502.1 | Tacheng cheese |
| NY-4 | Yarrowia | Yarrowia lipolytica | KY110196.1 | Habahe County cheese |
| TY-6 | Yarrowia | Yarrowia lipolytica | MZ314862 | Tashkurgan County cheese |
| TY-17 | zeylanoides | Candida zeylanoides | MZ314864 | Tashkurgan County cheese |
| 2TY-1 | Pichia | Pichia kudriavzevii | OM995975 | Tashkurgan County cheese |
| TY-25 | Guehomyces | Guehomyces pullulans | MZ314866 | Tashkurgan County cheese |
| 2TY-9 | Wickerhamomyces | Wickerhamomyces anomalus | OM995983 | Tashkurgan County cheese |
| 4TY-6 | Kluyveromyces | Kluyveromyces marxianus | OM995998 | Tashkurgan County cheese |
| 5TY-2 | Issatchenkia | Issatchenkia orientalis | OM996002 | Tashkurgan County cheese |
| 6TY-2 | Saccharomyces | Saccharomyces cerevisiae | OM996013 | Tashkurgan County cheese |
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.; Mandeep; Shukla, P. Probiotics of Diverse Origin and Their Therapeutic Applications: A Review. J. Am. Coll. Nutr. 2020, 39, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Mahdavi Abhari, F. The role of plant-derived natural antioxidants in reduction of oxidative stress. Biofactors 2022, 48, 611–633. [Google Scholar] [CrossRef] [PubMed]
- Paulino Do Nascimento, L.C.; Lacerda, D.C.; Ferreira, D.J.S.; De Souza, E.L.; De Brito Alves, J.L. Limosilactobacillus fermentum, Current Evidence on the Antioxidant Properties and Opportunities to be Exploited as a Probiotic Microorganism. Probiotics Antimicrob. Proteins 2022, 14, 960–979. [Google Scholar] [CrossRef]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Liu, A.; Hu, S.; Ares, I.; Martinez-Larranaga, M.R.; Wang, X.; Martinez, M.; Anadon, A.; Martinez, M.A. Synthetic phenolic antioxidants: Metabolism, hazards and mechanism of action. Food Chem. 2021, 353, 129488. [Google Scholar] [CrossRef]
- Kim, M.R. Antioxidants of Natural Products. Antioxidants 2021, 10, 612. [Google Scholar] [CrossRef]
- El-Sayed, M.I.; Awad, S.; Abou-Soliman, N.H.I. Improving the antioxidant properties of fermented camel milk using some strains of Lactobacillus. Food Nutr. Sci. 2021, 12, 352–371. [Google Scholar]
- Kahar, G.; Rahman, N.; Yusan, D.; Yalkun, M. Molecular identification andantioxidant activity of yeasts isolated from traditional fermented milk in southern Xinjiang. Food Mach. 2018, 34, 12–19. [Google Scholar]
- Abreu, I.; Albuquerque, R.; Brandao, A.B.P.; Barssotti, L.; De Souza, L.B.; Ferreira, F.G.; Oliveira, L.C.G.; Yokota, R.; Sparvoli, L.G.; Dias, D.D.S.; et al. Saccharomyces boulardii exerts renoprotection by modulating oxidative stress, renin angiotensin system and uropathogenic microbiota in a murine model of diabetes. Life Sci. 2022, 301, 120616. [Google Scholar] [CrossRef]
- Wang, Q.; He, Y.; Li, X.; Zhang, T.; Liang, M.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus reuteri CCFM8631 Alleviates Hypercholesterolaemia Caused by the Paigen Atherogenic Diet by Regulating the Gut Microbiota. Nutrients 2022, 14, 1272. [Google Scholar] [CrossRef]
- Brandao, L.R.; De Brito Alves, J.L.; Da Costa, W.K.A.; Ferreira, G.A.H.; De Oliveira, M.P.; Gomes Da Cruz, A.; Braga, V.A.; Aquino, J.S.; Vidal, H.; Noronha, M.F.; et al. Live and ultrasound-inactivated Lacticaseibacillus casei modulate the intestinal microbiota and improve biochemical and cardiovascular parameters in male rats fed a high-fat diet. Food Funct. 2021, 12, 5287–5300. [Google Scholar] [CrossRef]
- Fang, Y.; Chen, H.Q.; Zhang, X.; Zhang, H.; Xia, J.; Ding, K.; Fang, Z.Y. Probiotic administration of lactobacillus rhamnosus GR-1 attenuates atherosclerotic plaque formation in ApoE-/- mice fed with a high-fat diet. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3533–3541. [Google Scholar] [PubMed]
- Yasheng, M.; Rixat, E.; Rahman, N. Tolerance and probiotic characteristics of lactic acid bacteria in traditional fermented raw cheese in northern Xinjiang. Microbiol. China 2023, 50, 2044–2062. [Google Scholar]
- Kim, S.; Lee, J.Y.; Jeong, Y.; Kang, C.-H. Antioxidant Activity and Probiotic Properties of Lactic Acid Bacteria. Fermentation 2022, 8, 29. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Zhang, Y.; Li, X.; Mu, G.; Tuo, Y. Screening Human-derived Lactobacillus Strains with Antioxidant Activity. Food Res. Dev. 2019, 40, 199–205. [Google Scholar]
- Yanhong, L.; Bo, J.; Tao, Z.; Wanmeng, M.; Jian, L. Antioxidant and free radical-scavenging activities of chickpea protein hydrolysate (CPH). Food Chem. 2008, 106, 444–450. [Google Scholar]
- Song, S.; Liu, X.; Zhao, B.; Abubaker, M.A.; Huang, Y.; Zhang, J. Effects of Lactobacillus plantarum Fermentation on the Chemical Structure and Antioxidant Activity of Polysaccharides from Bulbs of Lanzhou Lily. ACS Omega 2021, 6, 29839–29851. [Google Scholar] [CrossRef]
- Goto, M.; Kuda, T.; Shikano, A.; Charrouf, Z.; Yamauchi, K.; Yokozawa, M.; Takahashi, H.; Kimura, B. Induction of superoxide anion radical-scavenging capacity in an argan press cake-suspension by fermentation using Lactobacillus plantarum Argan-L1. LWT-Food Sci. Technol. 2019, 100, 56–61. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, P.; Lou, L.; Zhan, J.; Fan, M.; Li, D.; Liao, Q. Antioxidant Activities of Lactic Acid Bacteria for Quality Improvement of Fermented Sausage. J. Food Sci. 2017, 82, 2960–2967. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Hao, X.Y.; Zhang, X.X.; Zhang, G.X.; Li, X.D.; Liu, L.; Sun, Y.; Pan, Y. Identification of antioxidant peptides from cheddar cheese made with Lactobacillus helveticus. LWT-Food Sci. Technol. 2021, 141, 110866. [Google Scholar] [CrossRef]
- Wang, C.F.; Huang, C.R.; Lu, Y.C. Changes in Bio-Functional Compounds, ACE Inhibition, and Antioxidant Capacity after Mixed Fermentation of Eight Whole Grains. Fermentation 2023, 9, 209. [Google Scholar] [CrossRef]
- Lin, M.Y.; Yen, C.L. Reactive oxygen species and lipid peroxidation product-scavenging ability of yogurt organisms. J. Dairy Sci. 1999, 82, 1629–1634. [Google Scholar] [CrossRef]
- Chen, Q.; Kong, B.; Sun, Q.; Dong, F.; Liu, Q. Antioxidant potential of a unique LAB culture isolated from Harbin dry sausage: In vitro and in a sausage model. Meat Sci. 2015, 110, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Amanatidou, A.; Smid, E.J.; Bennik, M.H.J.; Gorris, L.G.M. Antioxidative properties of Lactobacillus sake upon exposure to elevated oxygen concentrations. FEMS Microbiol. Lett. 2001, 203, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-S.; Ma, Y.; Chen, L.-J.; Zhao, C.-H.; Maubois, J.-L.; Jiang, T.-M.; Li, H.-M.; He, S.-H. Antioxidant activity of two yeasts and their attenuation effect on 4-nitroquinoline 1-oxide inducedin vitrolipid peroxidation. Int. J. Food Sci. Technol. 2010, 45, 555–561. [Google Scholar] [CrossRef]
- Halliwell, B.; Adhikary, A.; Dingfelder, M.; Dizdaroglu, M. Hydroxyl radical is a significant player in oxidative DNA damage in vivo. Chem. Soc. Rev. 2021, 50, 8355–8360. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Takamura, H.; Matoba, T.; Terao, J. HPLC method for evaluation of the free radical-scavenging activity of foods by using 1,1-diphenyl-2-picrylhydrazyl. Biosci. Biotechnol. Biochem. 1998, 62, 1201–1204. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.R.; Chen, M.J.; Lin, C.W. Antimutagenic and antioxidant properties of milk-kefir and soymilk-kefir. J. Agric. Food Chem. 2005, 53, 2467–2474. [Google Scholar] [CrossRef]
- Mendonca, J.D.S.; Guimaraes, R.C.A.; Zorgetto-Pinheiro, V.A.; Fernandes, C.D.P.; Marcelino, G.; Bogo, D.; Freitas, K.C.; Hiane, P.A.; De Padua Melo, E.S.; Vilela, M.L.B.; et al. Natural Antioxidant Evaluation: A Review of Detection Methods. Molecules 2022, 27, 3563. [Google Scholar] [CrossRef]
- Martinez-Cayuela, M. Oxygen free radicals and human disease. Biochimie 1995, 77, 147–161. [Google Scholar] [CrossRef]
- Ren, D.; Li, C.; Qin, Y.; Yin, R.; Du, S.; Ye, F.; Liu, C.; Liu, H.; Wang, M.; Li, Y.; et al. In vitro evaluation of the probiotic and functional potential of Lactobacillus strains isolated from fermented food and human intestine. Anaerobe 2014, 30, 1–10. [Google Scholar] [CrossRef]
- Ding, W.; Wang, L.; Zhang, J.; Ke, W.; Zhou, J.; Zhu, J.; Guo, X.; Long, R. Characterization of antioxidant properties of lactic acid bacteria isolated from spontaneously fermented yak milk in the Tibetan Plateau. J. Funct. Foods 2017, 35, 481–488. [Google Scholar] [CrossRef]
- Zhao, X.; Hu, D.; Zhang, S.; Qi, Y.; Wu, Y.; Gao, B.; Liu, Z. Screening and Identification of Lactic ACID Bacteria in Traditional Pickles and Analysis of the Antioxidant Properties. China Condiment 2022, 47, 5–9. [Google Scholar]
- Huang, Y.; Jiang, J.; Zhang, K.; Yao, P. Screening of High Antioxidant Lactic Acid Bacteria from Preserved Vegetables in Northern Anhui Province. Farm Prod. Process. 2020, 2, 19–21, 28. [Google Scholar]
- Siesto, G.; Pietrafesa, R.; Infantino, V.; Thanh, C.; Pappalardo, I.; Romano, P.; Capece, A. In Vitro Study of Probiotic, Antioxidant and Anti-Inflammatory Activities among Indigenous Saccharomyces cerevisiae Strains. Foods 2022, 11, 1342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cai, Y.; Chen, Z.; Shi, H.; Zhou, Y.; Yang, Y.; Tu, R.; Chen, G.; Wang, S. Screening and Characterization of Potential Antioxidant Probiotics Isolated from the Gut of Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Front. Mar. Sci. 2022, 9, 1175. [Google Scholar] [CrossRef]
- Pieniz, S.; Andreazza, R.; Okeke, B.C.; Camargo, F.A.; Brandelli, A. Antimicrobial and antioxidant activities of Enterococcus species isolated from meat and dairy products. Braz. J. Biol. 2015, 75, 923–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lili, D.; Xinran, L.; Yongyue, G.; Xiaoling, C.; Xiaomi, W.; Fengling, B.; Shumin, Y.; Xiaohua, G. Screening for and Identification of Lactic Acid Bacteria with Antioxidant Activity from the Intestinal Tract of Fish. Food Sci. 2021, 42, 127–132. [Google Scholar]
- Han, Q.; Kong, B.; Chen, Q.; Sun, F.; Zhang, H. In vitro comparison of probiotic properties of lactic acid bacteria isolated from Harbin dry sausages and selected probiotics. J. Funct. Foods 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Jie, W.; Wu, Y.; Bai, X.; Zhou, L. Isolation, Identification and Antioxidant Evaluation of Lactic Acid Bacteria from Sichuan Traditional Pickles. Food Ferment. Sci. Technol. 2022, 58, 35–41. [Google Scholar]
- Fakruddin, M.; Hossain, M.N.; Ahmed, M.M. Antimicrobial and antioxidant activities of Saccharomyces cerevisiae IFST062013, a potential probiotic. BMC Complement. Altern. Med. 2017, 17, 64. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, L. Comprehensive Evaluation of Probiotic Property, Hypoglycemic Ability and Antioxidant Activity of Lactic Acid Bacteria. Foods 2022, 11, 1363. [Google Scholar] [CrossRef]




| Strain Number | SOD | GPX | CAT Vitality | ||
|---|---|---|---|---|---|
| Inhibition Rate (%) | Vitality (U/mL) | Inhibition Rate (%) | Vitality (U/mL) | (U/mL) | |
| KM-14 | 29.50 ± 0.90 cd | 4.6545 ± 0.1995 cd | 9.90 ± 0.99 efg | 0.0340 ± 0.0034 efg | - |
| NM-14 | 39.55 ± 2.32 ab | 7.3254 ± 07368 ab | - | - | 19.7980 ±0.4009 d |
| TM-28 | 15.15 ± 0.60 e | 1.9854 ± 0.0927 e | 23.94 ± 2.82 de | 0.0579 ± 0.0068 de | 49.5629 ±2.5470 c |
| NM-12 | 30.35 ± 0.90 cd | 4.8464 ± 0.2069 cd | 34.45 ± 2.23 b | 0.1396 ± 0.0090 b | 19.9336 ± 3.0720 d |
| TM-27 | 40.88 ± 0.94 ab | 7.6904 ± 0.2942 ab | - | - | 16.3052 ± 1.8400 de |
| UM-12 | 44.99 ± 1.18 a | 9.1054 ± 0.4284 a | 31.73 ± 1.67 bc | 0.1123 ± 0.0059 bc | 3.7488 ± 0.9260 ef |
| UM-18 | 33.91 ± 0.86 bcd | 5.7053 ± 0.2181 bcd | 24.83 ± 1.34 bc | 0.1260 ± 0.0068 bc | 27.1753 ± 2.0760 d |
| TM-24 | 44.84 ± 1.34 a | 9.0542 ± 0.4795 a | 10.08 ± 1.46 ef | 0.0409 ± 0.0059 ef | - |
| NM-11 | 43.95 ± 1.03 a | 8.7241 ± 0.3646 a | 4.08 ± 1.02 fg | 0.0136 ± 0.0068 fg | 92.0948 ± 1.0170 a |
| AM-11 | 28.21 ± 0.79 d | 4.3687 ± 0.1717 d | 58.82 ± 1.68 a | 0.2383 ± 0.0068 a | - |
| QY-4 | 36.95 ± 0.79 bc | 6.5178 ± 0.2234 bc | 29.35 ± 1.88 cd | 0.0919 ± 0.0059 cd | 97.2890 ± 0.5346 a |
| TY-20 | 33.38 ± 3.84 bcd | 5.6766 ± 0.9557 bcd | 8.75 ± 2.50 fg | 0.0238 ± 0.0068 fg | 3.3724 ± 1.7850 ef |
| TY-6 | 35.10 ± 0.47 bcd | 6.0116 ± 0.1247 bcd | 22.73 ± 0.91 cd | 0.0851 ± 0.0034 cd | - |
| TY-25 | 29.60 ± 1.44 cd | 4.6842 ± 0.3294 cd | 11.54 ± 1.67 ef | 0.0409 ± 0.0059 ef | 63.9560 ± 2.1760 b |
| 6TY-2 | 27.67 ± 0.59 d | 4.2520 ± 0.1239 d | 27.27 ± 4.16 ef | 0.1021 ± 0.0156 bc | 14.8899 ± 2.1130 de |
| PBS | 5.32 ± 0.32 e | 0.6230 ± 0.0409 e | 3.74 ± 0.94 fg | 0.0255 ± 0.0034 fg | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abduxukur, D.; Tursuntay, A.; Zhu, X.; Wang, X.; Rahman, N. Antioxidant Capacity of Lactic Acid Bacteria and Yeasts from Xinjiang Traditional Fermented Dairy Products. Fermentation 2023, 9, 639. https://doi.org/10.3390/fermentation9070639
Abduxukur D, Tursuntay A, Zhu X, Wang X, Rahman N. Antioxidant Capacity of Lactic Acid Bacteria and Yeasts from Xinjiang Traditional Fermented Dairy Products. Fermentation. 2023; 9(7):639. https://doi.org/10.3390/fermentation9070639
Chicago/Turabian StyleAbduxukur, Dilihumar, Adila Tursuntay, Xiaoying Zhu, Xiaoyi Wang, and Nurgvl Rahman. 2023. "Antioxidant Capacity of Lactic Acid Bacteria and Yeasts from Xinjiang Traditional Fermented Dairy Products" Fermentation 9, no. 7: 639. https://doi.org/10.3390/fermentation9070639