1. Introduction
Currently, one of the main challenges in the fight against climate change is the search for renewable raw materials that replace fossil resources to generate energy, chemicals and materials. As a consequence, there is an urgent need for platform chemicals obtained through sustainable bioprocesses as alternatives to processes and products obtained via petroleum-based conventional refineries [
1,
2]. The majority of the processes and products deemed promising are being developed in second-generation biorefineries based on lignocellulosic biomass (LCB). LCB is especially attractive, as it is the most abundant and sustainable group of raw materials available worldwide and does not interfere in any food chain [
3]. LCB is mainly composed of cellulose (40–50%), hemicellulose (25–30%) and lignin (15–20%). Depolymerised hemicellulose is composed of 90% xylose, being this monosaccharide the second most abundant sugar available in LCB after glucose, a constituent of cellulose [
4]. However, the number of wild-type microorganisms that can metabolise xylose as a carbon source is very low, meaning that the hemicellulosic part of the waste is usually disposed of instead of being used as a carbon source.
One of the most promising microorganisms in the conversion of xylose into SA is a facultative anaerobic bacterium isolated from the rumen of cattle,
Actinobacillus succinogenes. This bacterium is capable of naturally generating C4 dicarboxylic acids during the pregastric digestion of polysaccharides. In other words, it can produce high amounts of succinic acid from a wide variety of carbon sources. In addition, its use does not generate excessive difficulties in fermentation or in the separation and purification processes, which occurs with certain fungi. This microorganism transforms xylose into glyceraldehyde 3-phosphate through the pentose phosphate pathway. Subsequently, the bacterium incorporates the final product of glycolysis, phosphoenolpyruvate, into the tricarboxylic acid cycle, as shown in
Figure 1. This occurs under anaerobic conditions and is facilitated by CO
2 insufflation, resulting in the production of a substantial amount of succinic acid (SA) [
5,
6]. According to the US Department of Energy (US DOE), SA is one of the main chemical platforms due to its potential to produce a great diversity of chemicals traditionally produced from fossil sources [
7]. This organic acid is widely used in the production of polybutylene succinate, polyester, polyols and in the food and pharmaceutical industries as well as in resins, coatings, pigments and biodegradable polymers [
8].
In recent years, the lack of sustainability of traditional petrochemical SA production has led to a considerable increase in its production through fermentation processes. In 2015, the market price of this acid produced by the biological route was 2.86 USD·kg
−1, while that obtained by the traditional route was 2.50 USD·kg
−1 [
9]. However, due to the great interest in biotechnological processes involving SA, currently, some economically competitive processes have already been developed and even implemented at an industrial level, producing SA with a market price between 2.00 and 2.50 USD·kg
−1 [
10]. With a 20% compound annual growth rate (CAGR), the bio-derived SA market is expected to reach 900 USD million by 2026, much higher than that of 2017 (175.7 USD million) [
11]. The three main factors to consider in the implementation of a successful SA bioprocess at an industrial scale are the following: (i) availability of raw material, (ii) adequate SA productivity and titre and (iii) economically viable isolation and purification steps [
12].
Ferone et al. (2017) [
13] studied the production of succinic acid in batch operational mode by
A. succinogenes employing a synthetic mixture of sugars representative of a lignocellulosic hydrolysis, obtaining a concentration of SA (27 g·L
−1) higher than the ones obtained by a combination of fermentations of each single sugar, as well as a better selectivity for SA. Using the same microorganism, Bukhari et al. (2020) [
14] used oil palm trunk as carbon source after its hydrolysis with oxalic, formic and acetic acid. Carrying out the bioprocess to obtain SA with hydrolysed waste, they obtained a yield of 0.47 g·g
−1 in batch bottle fermentation and a maximum concentration of succinic acid of 10.62 g·L
−1. In order to enhance yield and productivity, some researchers have recently focused on optimising the operational mode. Bradfield et al. (2016) [
15] reached productivities of 3.4 g·L
−1·h
−1 and a maximum SA titre of 10.9 g·L
−1 from xylose operating in continuous mode in a biofilm reactor. Jokodola et al. (2022) [
16] opted for a fed-batch operational mode, producing 33.6 and 28.7 g·L
−1 of succinate from hydrolysates of olive pits and sugarcane bagasse, respectively, with same conversion yield (0.27 g·g
−1). Considering that downstream operations represent approximately 60% of the overall operating costs in the SA production process, it is essential to reduce the by-product generation during fermentation in order to design a competitive process on an industrial level. To do this, this research group carried out a previous study on the viability of succinic acid production employing a biocatalyst composed of
A. succinogenes cells in a resting state [
17]. Operating under nitrogen-limited conditions, and formulating a medium composed exclusively of a carbon source and a buffer solution to maintain osmotic pressure, resting cells are metabolically active even though cell growth is impeded. In this way, the metabolic pathways are active towards SA production and cellular maintenance, but there is no bacterial growth. Concurrently, the number of by-products is also reduced dramatically [
18,
19,
20], this being one of the main advantages associated with this type of operation. In fact, Escanciano et al. (2022) [
17] managed to reduce the quantity of by-products generated by 27.5% in the production of SA from xylose. In order to obtain cells for subsequent use in the production stage in resting state, it is necessary to carry out a conventional fermentation, although it requires the optimisation of conditions so that the cells have the appropriate metabolic state for their subsequent use in resting cells. A notable advantage of separating the stages of growth (or obtaining the biocatalyst) and production (with the cells in the resting cell state) is the culture medium needed in the stages. Generally, to carry out production with growing cells, culture media with components such as yeast extract are used, which, although they do not have a high concentration in the medium, if the total mass is calculated to carry out the process in industrial bioreactors, would entail a high price. When the biomass is produced, it is done on a much smaller scale, so the need for such culture media is considerably reduced. However, the medium used for resting cell production is very simple and cost-effective, devoid of nitrogen sources and usually expensive. Separating growth and production into two different steps allows for a better optimisation of each step and of the overall process. Considering the scale-up, the proposed bioprocess would lead to higher productivity linked to the reduction of dead times as well as to avoid substrate inhibition that was observed when using growing cells as biocatalyst [
18,
20,
21].
In this present study, SA production using
A. succinogenes in both growth and resting states was researched, exploring different forms of operation through biocatalyst reuse by means of fed-batch and repeated batch operations. This is the first work in which the operation with resting cells is studied in different types of operation, allowing us to optimise the stages of cell growth and SA production separately and reduce the operating costs associated with the nitrogen source and SA purification. Additionally, the effect on production with cells in growth and resting state of the use of a cheaper culture medium than the one used in previous studies of the research group was compared [
17]. Finally, kinetic modelling of the bioprocess was developed to predict the evolution of biomass, xylose, succinic acid and the by-product concentration throughout the fermentation time.
2. Materials and Methods
2.1. Bacterial Strain, Adaptation, Preinoculum and Inoculum Stages
The strain
Actinobacillus succinogenes DSM 22,257 was provided by the Leibniz Institute DSMZ-German Collection of Microorganisms and Cultures GmbH (
https://www.dsmz.de/accessed on 15 February 2023). The bacterial strain was reactivated on one agar plate with Brain Heart Infusion (BHI) for two days. A single bacterial colony was inoculated in a bottle containing 60 mL of sterile Tryptic Soy Broth (TSB) [
22,
23,
24]. The bottle was incubated for 1 day at 37 °C. For the long-term preservation, the culture was mixed with glycerol (1:1 v·v
−1) and stored at −80 °C.
Following the procedure described in a previous study [
17], thawed bacteria were incubated at 37 °C for 24 h in anaerobic bottles with TSB medium. Subsequently, a two-step procedure was developed to adapt the biocatalyst to the carbon source. Broth from anaerobic bottles was employed as inoculum in a 5% (v·v
−1) ratio by inoculating it in a second bottle containing the production medium (PM) at 37 °C [
25]. This production medium contained (in g·L
−1): yeast extract, 10; K
2HPO
4, 3; MgCl
2.6H
2O, 0.427; CaCl
2, 0.2; NaCl, 1; NaHCO
3, 10; and commercial xylose, 10. Using an inoculum of this second bottle, another stage of adaptation in a third bottle was carried out, increasing the concentration of xylose and NaHCO
3 to 20 g·L
−1 and the biomass initial inoculation volume from the previous step to 10% (v·v
−1).
2.2. Succinic Acid Production Employing Growing Cells as the Biocatalyst
Runs were carried out in duplicate in a 2 L stirred tank bioreactor (STBR) BIOSTAT B-Plus (Sartorius AG, Göttingen, Germany) with a working volume of 1 L. Batch experiments were carried out with PM medium. The initial amount of xylose was 20 g·L−1 in all runs. The pH was maintained at 6.8 (NaOH 5 M) at 37 °C and CO2 was sparged at 0.1 vvm with a stirring speed of 300 rpm. The experiments were carried out starting from 0.05 g·L−1 of biomass in the exponential phase of growth, obtained in the final adaptation step in bottles.
2.3. Succinic Acid Production Employing Resting Cells as the Biocatalyst
To successfully perform a fermentation process employing cells in a resting state as the biocatalyst, it is necessary to previously carry out a batch operational mode with growing cells as was described in previously, in order to obtain a high amount of initial biomass. After 15 h of fermentation, suspended biomass was separated from the broth by centrifugation (9000 rpm, 5 min), washed with a K2HPO4 solution and transferred to a 1 L STBR with 0.5 L of working volume at 37 °C and 300 rpm. The production media consisted of a buffered solution (K2HPO4 50 mM) of xylose 20 g·L−1. The pH was controlled at 6.8 (NaOH 5 M), and carbon dioxide was supplied at 0.1 vvm. Runs were performed in duplicate.
2.4. Succinic Acid Production Operating Fed-Batch and Repeated Batch Fermentations
Fed-batch and repeated batch runs were carried out in duplicate with PM medium both with growing cells and resting cells biocatalysts, In the case of the fed-batch operation, three stages were carried out, feeding a concentrated solution of xylose at the beginning of each one of them. In repeated batch fermentations, at the end of the first and second stages, the suspended biomass was separated from the liquid broth by centrifugation and subsequently inoculated in the reactor of the next stage. The pH was maintained at 6.8 (NaOH 5 M), temperature at 37 °C and the stirring speed at 300 rpm, and CO2 was sparged at 0.1 vvm.
2.5. Analytical Methods
Biomass concentration was determined by measuring the optical density of broth samples at 600 nm using a spectrophotometer (Shimadzu UV-Vis spectrophotometer UV-1603, Kyoto, Japan).
Xylose and fermentation products (ethanol and succinic, lactic, acetic and formic acids) were analysed through an Agilent Technologies 100 series equipment by high-performance liquid chromatography (HPLC) equipped with a refractive index detector (RID) at 55 °C and a REZEX ROA-Monosaccharide H+ (8%) column (300 × 7.8 mm, Phenomenex, Torrance, CA, USA) at 80 °C. Acid water (H2SO4 5 mM) was eluted at a flow of 0.5 mL·min−1.
2.6. Mathematical Methods
The estimation of the model kinetic parameters was carried out by means of the computer software Aspen Custom Modeler v11 (AspenTech, Bedford, MA, USA), using an implicit Euler method to integrate the ODEs of the kinetic model coupled to a non-linear least-squares solver algorithm (NL2SOL) to obtain optimal values of the kinetic parameters.
To assess the adequacy of the fit, it is necessary to ensure that Fisher’s F-value (F)—as seen in Equation (1)—has a higher value than the critical value at a 95% confidence level. Additionally, the sum of squared residuals (SSR) and the residual mean-squared error (RMSE) should be minimised, approaching zero as closely as possible—as seen in Equation (2)—and the variation explained (VE) should be close to or equal to 100%—as seen in Equation (3).
where
K is the number of parameters,
SSR is the squared sum of residues,
N is the total number of experimental data,
SSQl is the sum of the quadratic residues and
SSQmeanl is the squared sum of deviations between the experimental and the mean score with respect to the calculated values.
In order to determine the influence of initial substrate concentration, consumed substrate, product distribution and cell metabolism on succinic acid production, the following parameters were defined:
where [
SA]
max is the maximum concentration of succinic acid (
SA) (g
SA·L
−1), [
Xyl]
0 is the initial concentration of xylose (g
Xyl.
0·L
−1), [
Xyl]
cons is the concentration of consumed xylose (g
Xlycons·L
−1), [
X]
max is the maximum concentration of biomass (g
X·L
−1) and [
BP]
max is the maximum concentration of by-products [
BP] (g
BP·L
−1).
3. Results and Discussion
3.1. Influence of Medium Composition on Succinic Acid Production Employing Growing Cells and Resting Cells as the Biocatalyst
Growth medium composition as well as type and availability of carbon source play an important role in SA titre and productivity. Furthermore, the development of an effective biocatalyst is also a key factor that determines the viability of the bioprocess. Recently,
A. succinogenes in the resting state was proven to be a promising biocatalyst, revealing a higher selectivity to the target acid when compared to growing cells [
17]. In that previous study [
17], a complex growth medium, TSB, had been selected due to its richness in nutrients, considered in the bibliography as a preferred medium to promote cell growth [
22,
23,
24]. However, with the aim of reducing the cost of the process, the present study verifies the viability of carrying out cell growth, prior to the operation with resting cells, with a lower-cost synthetic medium, PM.
Table 1 summarises the obtained results in batch runs, in terms of the maximum product concentration achieved; its selectivity, yields and productivities in each evaluated medium culture (PM and TSB); and the biocatalyst state (growing and resting cells). Regarding product concentration, it is noticed that the synthetic medium promotes SA production, whose final concentration in broth reaches the highest values when the PM medium is employed (11.7 g‧L
−1 and 12.5 g‧L
−1 using growing and resting cells, respectively). As a consequence, in these conditions, higher values of SA yield with respect to initial carbon source are achieved (0.55 g
SA‧g
Xyl.
0−1 for growing state and 0.65 g
SA‧g
Xyl.
0−1 for resting state) than in runs in which the TSB medium was used. However, in fermentations with PM medium culture, an increase of by-product production (acetic acid and formic acid) is observed, a fact that is reflected in a slight reduction of selectivity values.
Medium composition barely affects SA yield respect to final biocatalyst concentration employing growing cells. However, a biocatalyst composed of resting cells cultivated in synthetic broth presents cell metabolism pathways that more actively produce SA. This is reflected in the increase of this acid yield respect to biocatalyst concentration (2.91 gSA‧gX−1) in comparison to the value when TSB is employed (2.03 gSA‧gX−1). A similar tendency is observed for productivity (PSA), whose meaning is closely linked to a macroscopic SA production rate. This acid productivity is boosted by the PM medium employing the resting cells as a biocatalyst. However, this is not improved in growing cell. As the results showed, acid production from resting cells takes place 2.4 times faster due to the change of the medium composition (with synthetic medium culture: 0.43 gSA‧L−1‧h−1; when using complex medium culture: 0.18 gSA‧L−1‧h−1).
Based on these results, it can be concluded that the employment of the synthetic growth medium PM using A. succinogenes in growing and resting states enhances SA production in terms of final titre and yield with respect to initial xylose concentration. In addition, productivity and yield with respect to cell concentration are also improved for the biocatalyst in resting cells. In short, an expensive complex growth medium such as TSB is not necessary when producing with resting cells; an economical and synthetic medium such as PM is a promising alternative for the development of the bioprocess on an industrial scale.
Figure 2 shows the evolution of xylose (Xyl), product (SA), by-product (BP) and biomass (X) concentrations in batch bioreactor employing the PM medium. Substrate exhaustion is reached at around 30 h for both biocatalysts. However, the experimental tendencies are different: whereas lag phase of growth leads to slower substrate consumption and product production rates at the beginning of the fermentation (
Figure 2A), a quick decrease on xylose concentration and, therefore, SA production is observed when a resting cells biocatalyst is employed (
Figure 2B). Even if the achieved target product concentration is similar in both cases, the global by-product concentration is reduced employing resting cells biocatalyst. This behaviour is also reflected in the parameters showed in
Table 1. When synthetic medium is used, all the evaluated parameters are improved by resting cells in comparison to growing biocatalyst. This means that cells under nitrogen-limited conditions reveal a metabolism more specific and selective towards the production of SA from xylose as sole carbon source.
3.2. Succinic Acid Production in a Repeated Batch Bioreactor
As previously mentioned, some authors had managed to significantly increase their yields and/or productivities by carrying out fermentations with growing cells in other modes of operation [
15,
16,
26]. In addition, in previous studies this research group demonstrated in previous works that successive growth stages prior to the inoculum stage improve the yield and productivity of the process. Therefore, successive repeat batch fermentations could have this same effect [
17]. For this reason, with the aim of improving the production process of succinic acid with resting cells, as well as further reducing the costs associated with the separation stages, it was decided to carry out fermentations through a repeated batch operation.
In the present section, the possibility of reusing the biocatalyst in three consecutive cycles was evaluated. Essentially, repeated batch experiments explored the viability of separating the biocatalyst at the end of the fermentation and, afterwards, harvested cells were employed as inoculum in the next cycle, as was explained in
Section 2.4. Whereas the PM synthetic medium is used for the maintenance of the growing biocatalyst in repeated batch runs, only the carbon source and a buffer solution are employed as the medium for resting cells.
Each batch run was conducted employing approximately 20 g·L
−1 of initial xylose concentration, which corresponds to the concentration range of carbon source found in raw vegetable materials [
27]. When carbon substrate concentration reached 5 g·L
−1, the experiment was stopped in order to avoid a metabolic stress to the bacterial cells. Subsequently, the biocatalyst was separated from the broth by means of centrifugation and the recovered pellet was employed as a biocatalyst in the next batch run. The main advantage of repeated batch in comparison to fed-batch operation is that repeated batch avoids inhibition due to the accumulation of cytotoxic compounds in the broth, as may occur with the fed-batch operation [
28].
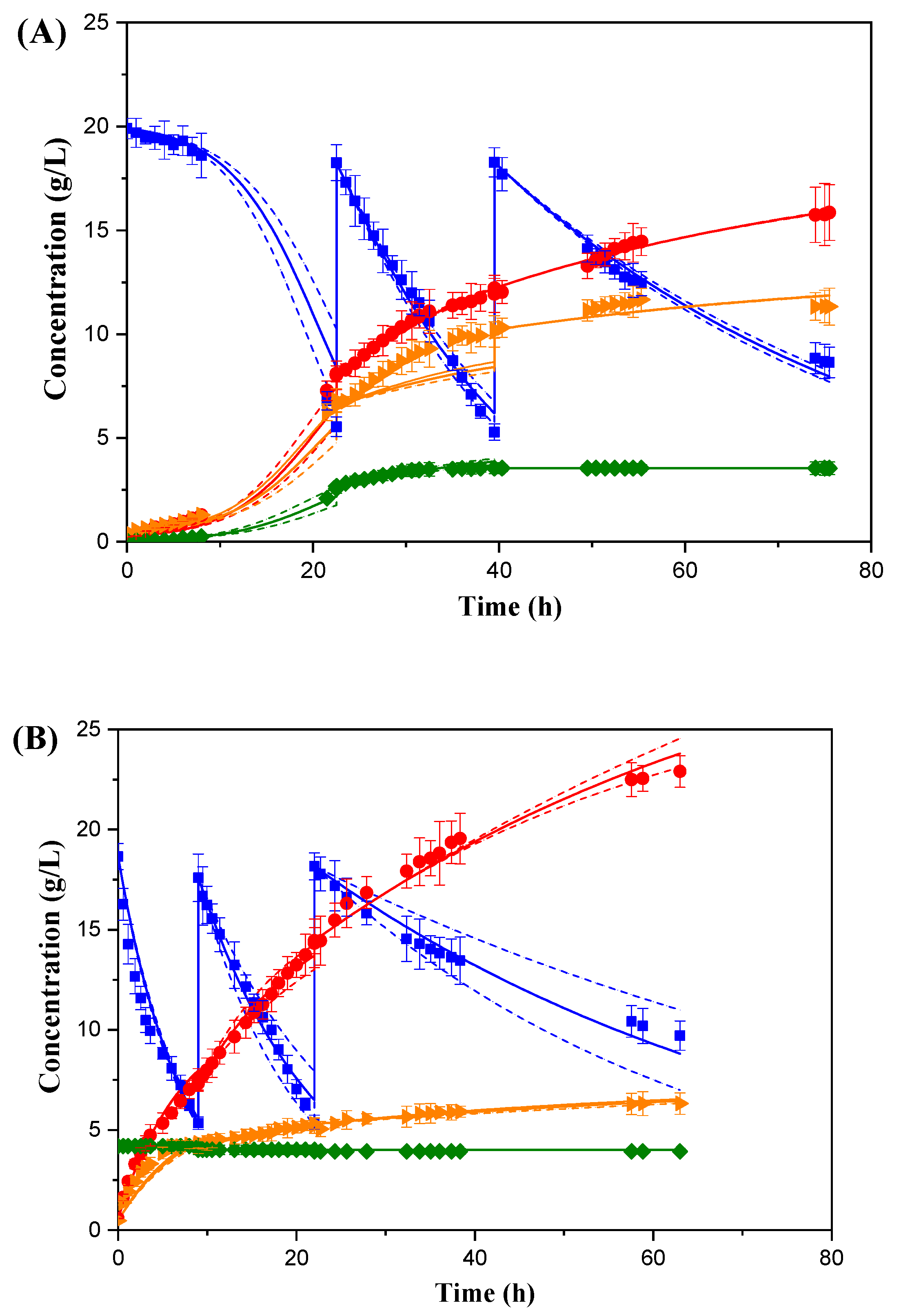
Figure 3 shows the evolution of xylose, biomass, SA and by-product concentrations in repeated batch cycles evaluating the influence of the state of the biocatalyst: growing cells (
Figure 3A) and resting cells (
Figure 3B). While three consecutive cycles employing growing cells took less than 60 h, the use of the resting cells biocatalyst extended the fermentation time by 40 h. However, the first cycle using a biocatalyst composed of resting cells was much faster than the analogous experiment for growing cells, which is probably due to the greater biocatalyst concentration at the beginning and throughout the run. In the following steps, a deceleration of substrate consumption rate is observed when resting cells are reused. Resting cells concentration decreases progressively, due to the experimental cell recovery procedure between cycles. It was observed that, although in the second cycle the cells were still considerably active, in the third cycle, the process suffered a significant slowdown, probably caused by the mechanical stress of the biocatalyst separation process and the scarcity of nutrients during the consecutive fermentation cycles.
Regarding repeated batch working with the growing biocatalyst, it is to be noted that productivity is enhanced throughout the cycles, in terms of both substrate consumption and acid production, as is shown in
Table 2. Although the initial biomass concentration at the beginning of each step is increased, the importance of the bacteria adaptation to carbon source also plays an important role in the bioprocess, as has previously been demonstrated by Escanciano et al. [
17]. In fact, with growing cells, the yield with respect to the xylose consumed and the productivity in the third stage are 72% and 56% higher than those corresponding to the first stage, respectively, reaching an average yield with respect to xylose consumed of 0.51 g g
−1 and a productivity of 0.53 g·L
−1·h
−1 (
Table 2). Regarding specific SA productivity, cells in growth state obtain higher values than the resting cells, maintaining practically constant productivities throughout the three stages, while in the runs with resting cells, the specific productivity is already reduced by 70% from the first to the second stage.
In conclusion, although selectivity was favoured by production with resting cells, the reuse of growing cells led to better overall SA production, in terms of concentration, yield, and productivity, which results improve from batch to the next one.
3.3. Succinic Acid Production in a Fed-Batch Bioreactor
To avoid cell damage during separation processes between cycles, fed-batch operation was explored as an alternative to extend operational time and, therefore, increase final SA titre and productivity. Three feeding pulses were carried out (at the beginning and when substrate concentration was around 5 g·L−1) to obtain similar conditions as the repeated batch operation, which allows for the comparison of the results.
Figure 4 shows the evolution of xylose, biomass, SA and by-product concentrations in the fed-batch operation employing different biocatalysts: growing cells (
Figure 4A) and resting cells (
Figure 4B).
Table 3 summarises the obtained results in terms of final concentrations, selectivity, yields and productivity after the consumption of each feeding pulse as well as overall parameters.
As can be observed, the duration of the first two stages of the fed-batch operation with resting cells was considerably shorter than the corresponding ones with growing cells. In both cases, the total substrate consumption was not reached in the last step. In fact, in the operation with resting cells, after 30 h of fermentation, only approximately half of the xylose fed in the third stage was consumed.
The slowdown of the processes in the last part of the experiments can be caused by various factors: the depletion of essential nutrients that are not added in the pulse feeding, the aging of the cells, the accumulation of acids (both the target product as well as by-products) or toxins with an inhibitory effect [
29]. This reduction in productivity at long fed-batch fermentation times has also been observed by other authors, such as Jiang et al. [
25], who, with fermentation from glucose, noted a drop in the growth rate of
A. succinogenes and of the succinic acid generated from the third production cycle (14 h of fermentation), which became especially pronounced in the fourth cycle.
It should also be noted that, when using cells in resting state, the selectivity of SA is fully enhanced, reaching a final concentration of SA 3.6 times higher than the global concentration of by-products. However, in the case of using growing cells, only a final concentration of SA 1.4 times higher than the total concentration of by-products is reached. Regarding SA yields, they had a decreasing trend between cycles in the growing cell experiment (cycle 1: 0.40 g‧g−1, cycle 2: 0.21 g‧g−1, cycle 3: 0.20 g‧g−1), while the opposite behaviour was observed with resting cells (cycle 1: 0.40 g‧g−1, cycle 2: 0.21 g‧g−1, cycle 3: 0.20 g‧g−1). The concentration of the final product was 1.5 times higher using resting cells (22.9 g‧L−1). In both cases, productivity was drastically reduced throughout the cycles, with a higher average value in the case of operating with resting cells (0.36 g‧L−1‧h−1). These results showed that the metabolism of the biocatalyst slowed down throughout the cycles but was more selective. In conclusion, the fed-batch operation with cells in the resting state leads to higher yield, productivity and selectivity than with growing cells.
Based on these results, the possibility of a reusing biocatalyst by means of repeated batch and fed-batch operational modes was successfully proven. Specifically, employing growing cells under repeated batch conditions and resting cells in fed-batch mode led to better SA production yields (0.51 g‧g
−1 and 0.68 g‧g
−1, respectively). In terms of productivity, production by growing cells in repeated batch mode took place faster (0.12 g·L
−1·h
−1 and 0.09 g·L
−1·h
−1, respectively). Moreover, working with growing cells in repeated batch mode, the third cycle results did not show inhibition or aging cell phenomena. However, in fermentation with resting cells in fed-batch mode, it was possible to increase the selectivity by 32% with respect to the operation in repeated batch with growing cells. This is of great importance if we consider that one of the main operating costs in the production of succinic acid is found in the purification process [
30].
Table 4 summarises published results regarding the production of SA mainly from xylose, using
A. succinogenes as s biocatalyst. Up to now, most of the studies have been undertaken with conventional submerged growth culture. The authors who carried out batch-type operations from xylose with growing cells did not achieve yields greater than 55% [
13] nor SA productivities higher than 0.36 g·L
−1·h
−1 [
17]. However, substantially higher yields have been achieved with other types of operation, as in the case of Patsalou et al. [
31], who managed to produce SA using citrus peel waste hydrolysate as a renewable carbon source with fed-batch type fermentation, reaching a yield of 0.73 g·g
−1. To date, the fermentations carried out operating in repeated batch mode have mostly been studied with glucose as carbon source (or hydrolysates rich in this sugar) and with immobilised cells, which is why this study has focused in part on the production of SA through this operation mode from xylose with free cells, achieving a yield of 51% and an average productivity of 0.12 g·L
−1·h
−1.
Productions with
A. succinogenes in a resting state were evaluated for the first time in a previous study of this research group [
17], achieving a batch yield of 43% while the productivity was 0.18 g·L
−1·h
−1. In this present work, it was possible to double the productivity thanks to the operation in fed-batch conditions. Therefore, these results are considered to pave the way for development novel biocatalyst and give new insight towards operational modes in SA production.
3.4. Kinetic Modelling of the Bioprocess
Kinetic modelling enables the simulation of bio/chemical processes and facilitates their implementation and operational control. To obtain a mathematical model able to fit the kinetic data in this work, a very simple unstructured and non-segregated kinetic model was proposed, which describes the observed experimental tendencies regarding xylose, biomass, SA and by-product concentrations. The following considerations were taken into account:
- -
A laboratory-scale bioreactor was considered as a completely mixed tank bioreactor.
- -
The only by-products detected in the culture medium were acetic and formic acids, which are formed almost directly from pyruvate, as is reflected in
Figure 1. Moreover, the evolution of their concentrations exhibited remarkable similarity across all experiments. Based on that, acetic and formic acids concentrations were lumped or summed to obtain a global by-product concentration in the kinetic model.
- -
Growth, SA production and BP generation depend proportionally on the availability of carbon source and the biocatalyst concentration.
Considering this information, a sole kinetic model (Equations (10)–(15)) was developed to describe the behaviour of growing cells and resting cells as biocatalysts for all operational modes. Employing resting cells as a biocatalyst, the biomass concentration remains constant, resulting in zero yield of biomass from product and biomass growth rate.
Substrate consumption rate
By-product production rate
where Y
i/j is the macroscopic yield of compound “i” with respect to compound “j”, and k
P is the second-order kinetic constant in the SA production.
The fitting of the proposed kinetic equation to experimental data (substrate, biomass, product and by-products) for each cycle employing both biocatalysts was carried out to estimate the value of each kinetic parameter involved in the kinetic model. The model prediction fits very reasonably to all relevant data, as is reflected in
Figure 3 and
Figure 4 as solid lines. Consequently, the hypotheses assumed in the model are valid in the studied conditions, where no substrate or product inhibition phenomena were considered.
Table 5 and
Table 6 show the estimated kinetic parameters, as well as the statistical parameters that provide the information on the goodness of fit. Fitting of experimental data regarding batch run and first cycles of repeated batch and fed-batch runs were carried out together. Goodness-of-fit statistical parameters indicate a high value for Fisher’s F, considerably above the limiting value, and a low value for the RMSE and the SSR. Moreover, the experimental tendencies and those predicted from the model are quite similar, as the higher than 90% VE percentages showed.
The influence of biocatalyst state and reutilisation cycles on the kinetic parameters values is shown in
Figure 5. The second-order kinetic parameter k
P decreases along the cycles of reutilisation, independently of biocatalyst state and operational mode. These results point at a biocatalyst deactivation throughout the fermentation cycles. However, the deactivation of growing cells under repeated batch takes place slower than it does under other conditions. Regarding yields (substrate from product, by-product from product and biomass from product), smaller values indicate an effective SA production bioprocess, as it requires less substrate and biomass for product generation and minimises side products generation. As it is reflected on yield tendencies, biocatalyst composed of resting cells is more specific for SA production, especially in the second and third uses.
4. Conclusions
In the present study, a bioprocess for SA production using a synthetic growth medium and xylose as the carbon source was analysed, which proved to offer economic advantages over a medium based on complex carbon and nitrogen sources. Furthermore, for the first time, the reutilisation of the biocatalyst composed by A. succinogenes cells under growing and resting states was explored under diverse operation modes. Interestingly, whereas the highest average SA yield and productivity in repeated batch mode were observed with growing cells, the best results in fed-batch mode were reached with resting cells, which had limited nutrients availability. In fact, in fed-batch mode, the yield of SA with respect to xylose consumed was 66% higher with resting cells than with growing cells. Furthermore, the selectivity was 37% higher. This means that, for the first time, it was possible to produce succinic acid during three cycles of xylose feeding in the absence of a nitrogen source from a single stage of cell growth, also achieving yields and selectivities superior to the equivalent operation with growing cells, in other words, in a culture medium with nitrogen source.
Carrying out the bioprocess with resting cells presents several biological and technical advantages: (i) metabolic pathways towards the target product are promoted, as indicated by the reduction in by-product generation; (ii) the culture medium only contains phosphate and sugar, eliminating the use of expensive nitrogen sources; and (iii) SA is accumulated in the broth during the cycles, resulting in a higher final concentration compared to using a growing cells biocatalyst. All these factors contribute to cost reduction in subsequent purification and isolation operations. Thus, the possibility of producing SA with A. succinogenes in resting state becomes an innovative option with great potential and opens up new opportunities for bioprocess intensification (e.g., biocatalyst immobilisation).
Finally, a simple non-structured non-segregated kinetic model was successfully developed to describe cell growth, SA production and by-product generation. The estimated kinetic parameters are supported by the observed experimental trends and provide valuable information for a later bioprocess scale-up. In short, this research affords key progresses towards the incorporation of SA in biorefineries on an industrial scale.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}